Responses of Protozoan Communities to Multiple Environmental Stresses (Warming, Eutrophication, and Pesticide Pollution)

 and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
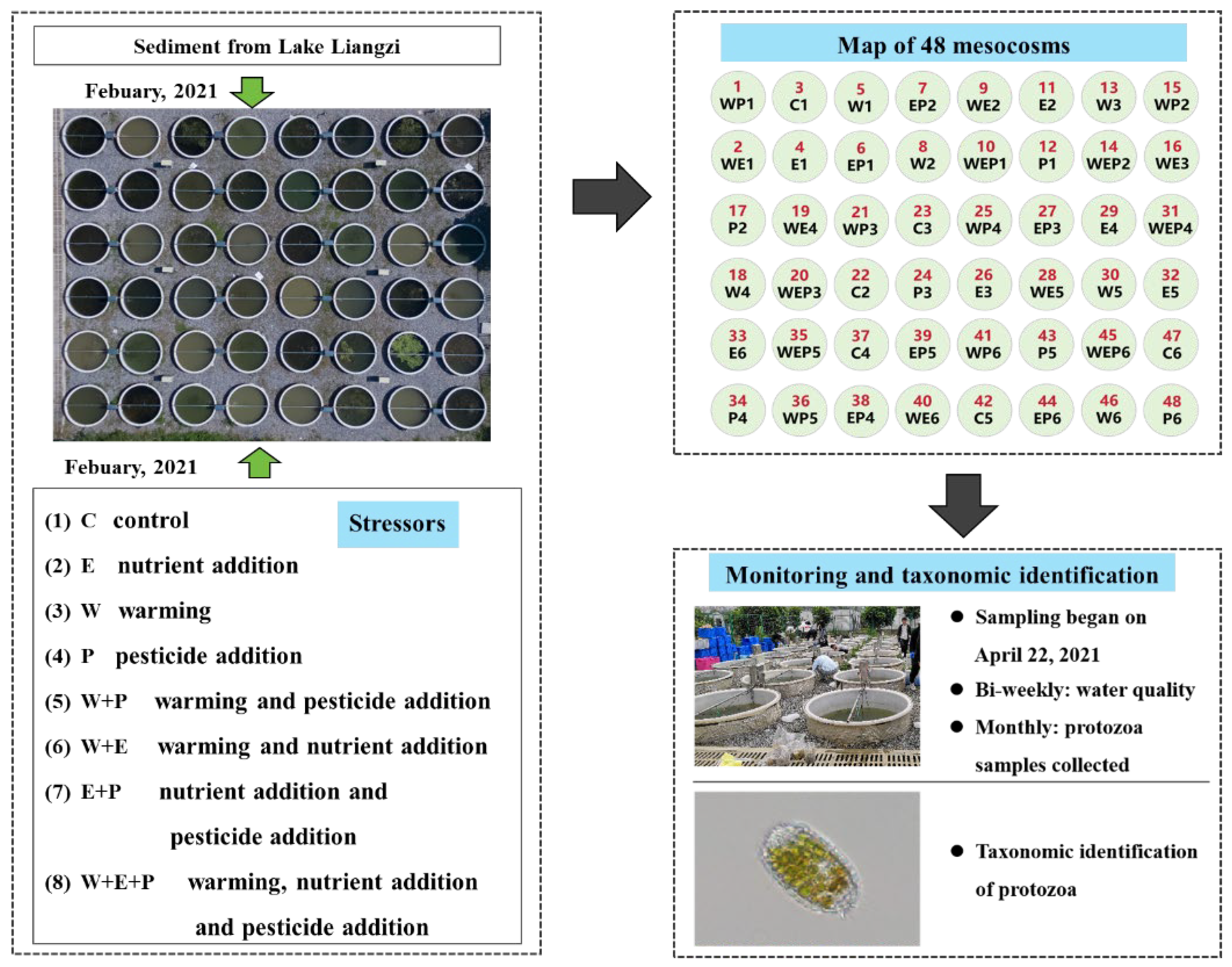
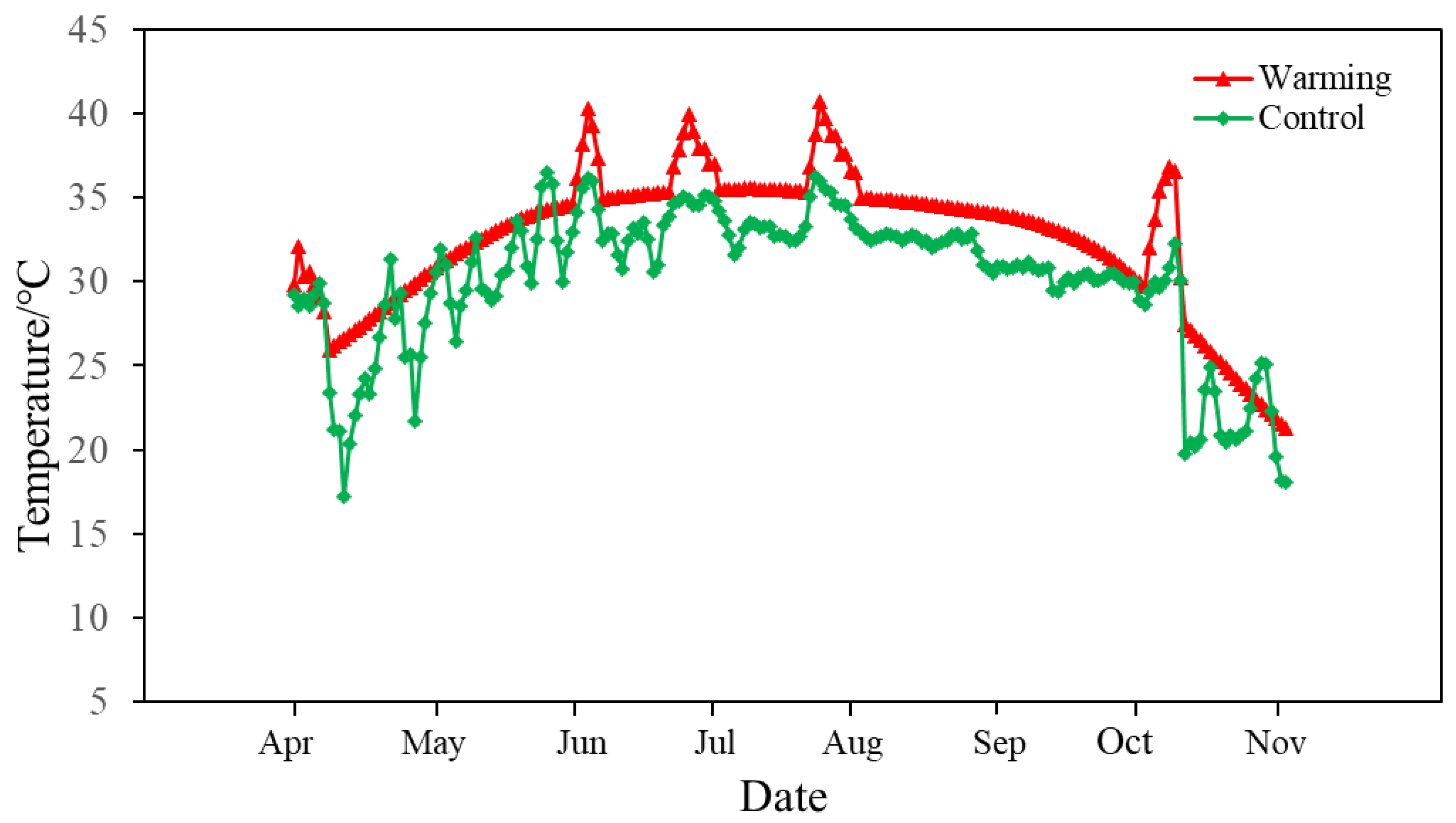
2.1. Experimental Design
2.2. Sample Collection and Processing
2.3. Data Processing and Analysis
J = H’/ln(S);
D = ∑n(n − 1)/N(N − 1).
3. Results
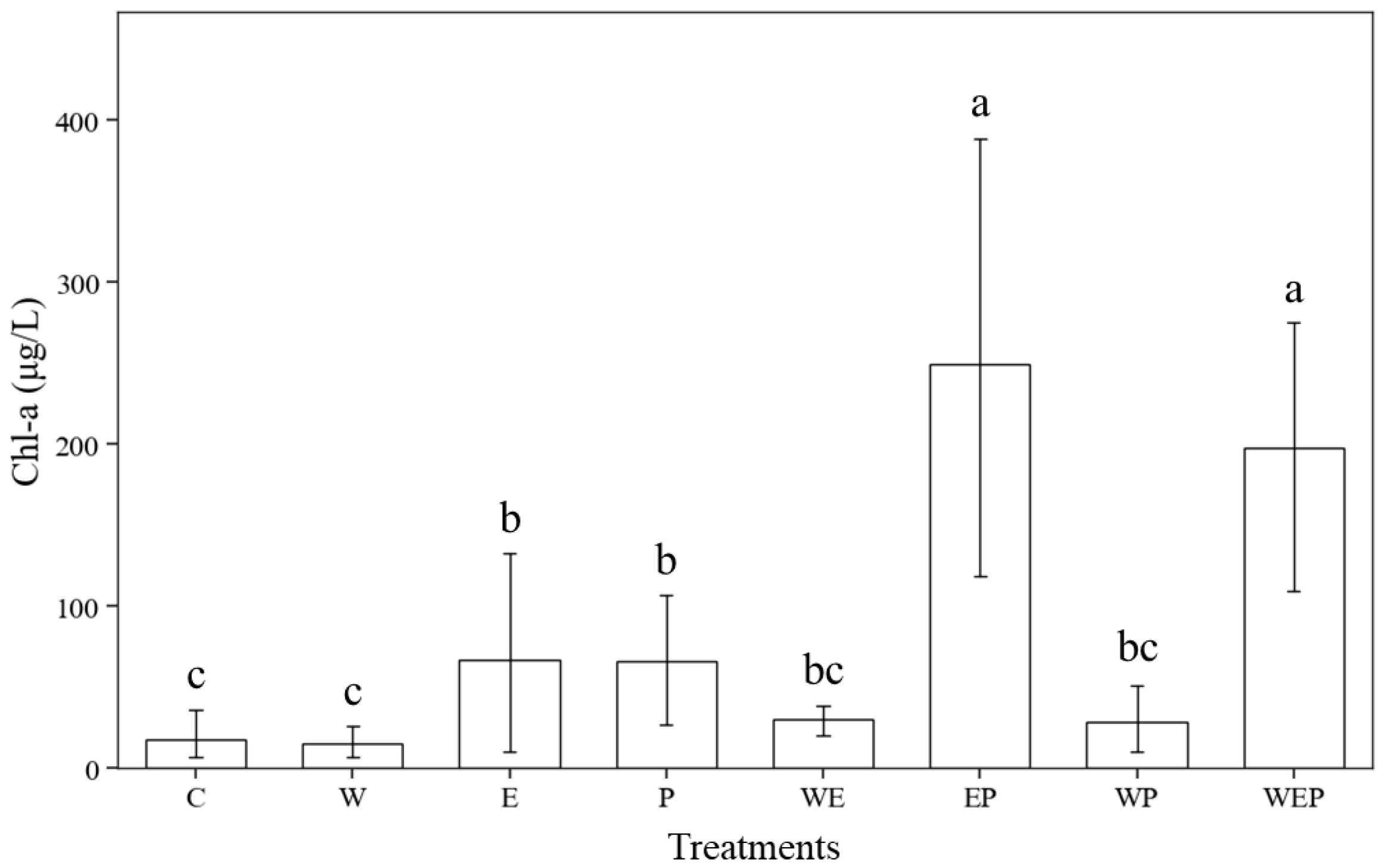
3.1. Effects of Different Treatments on Physical and Chemical Indexes of the Water Body
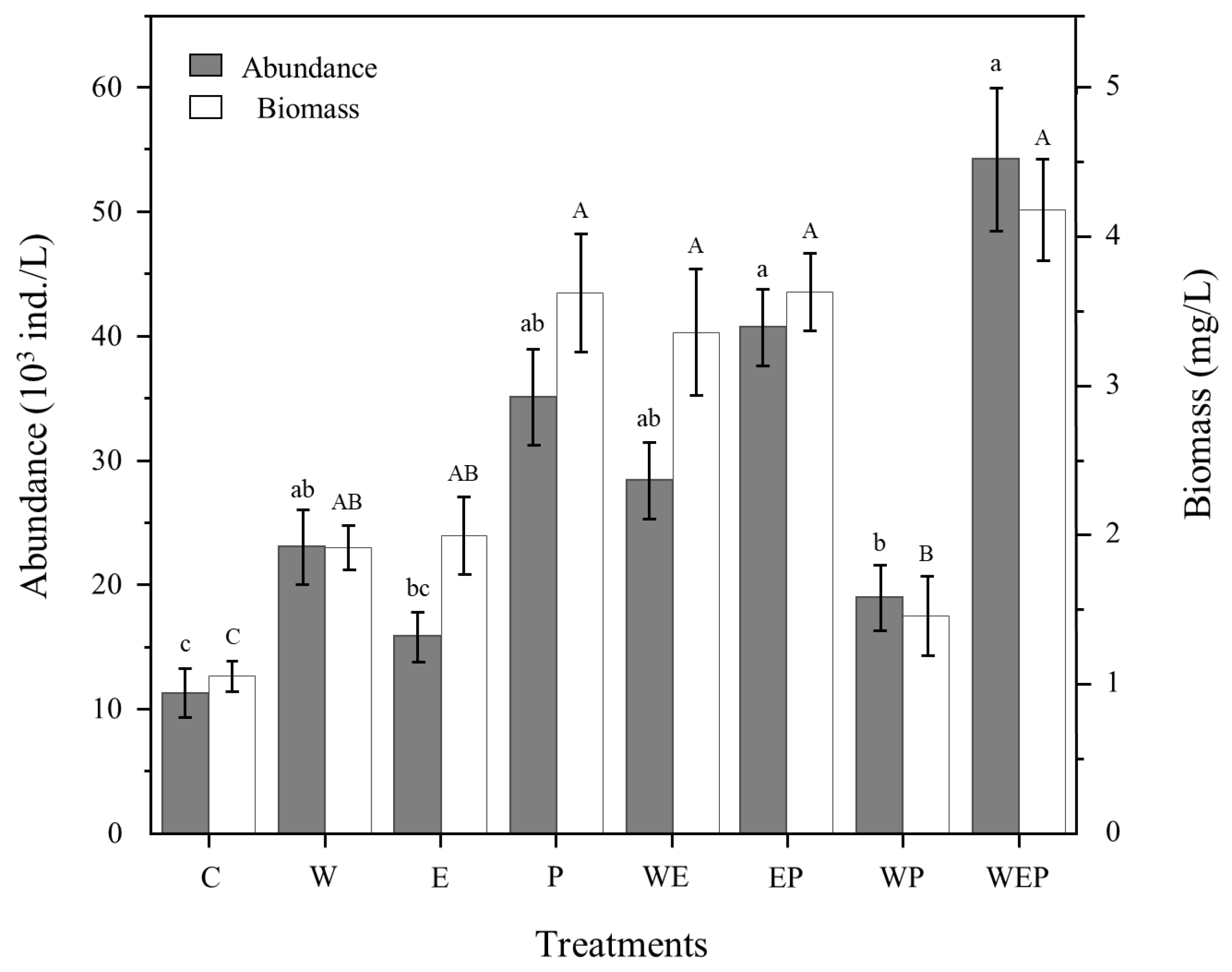
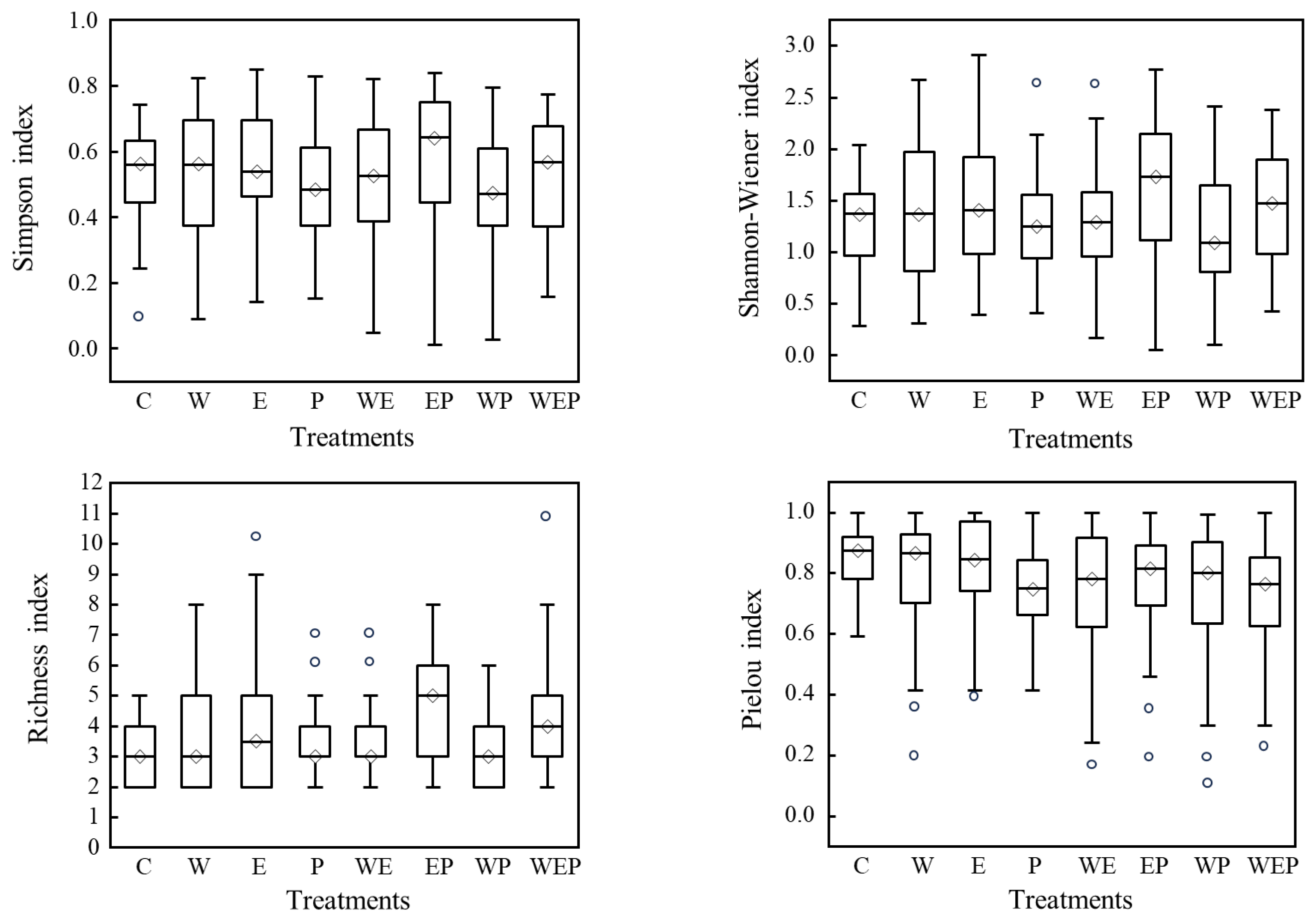
3.2. Effects of Different Treatments on Community Structure of Protozoa
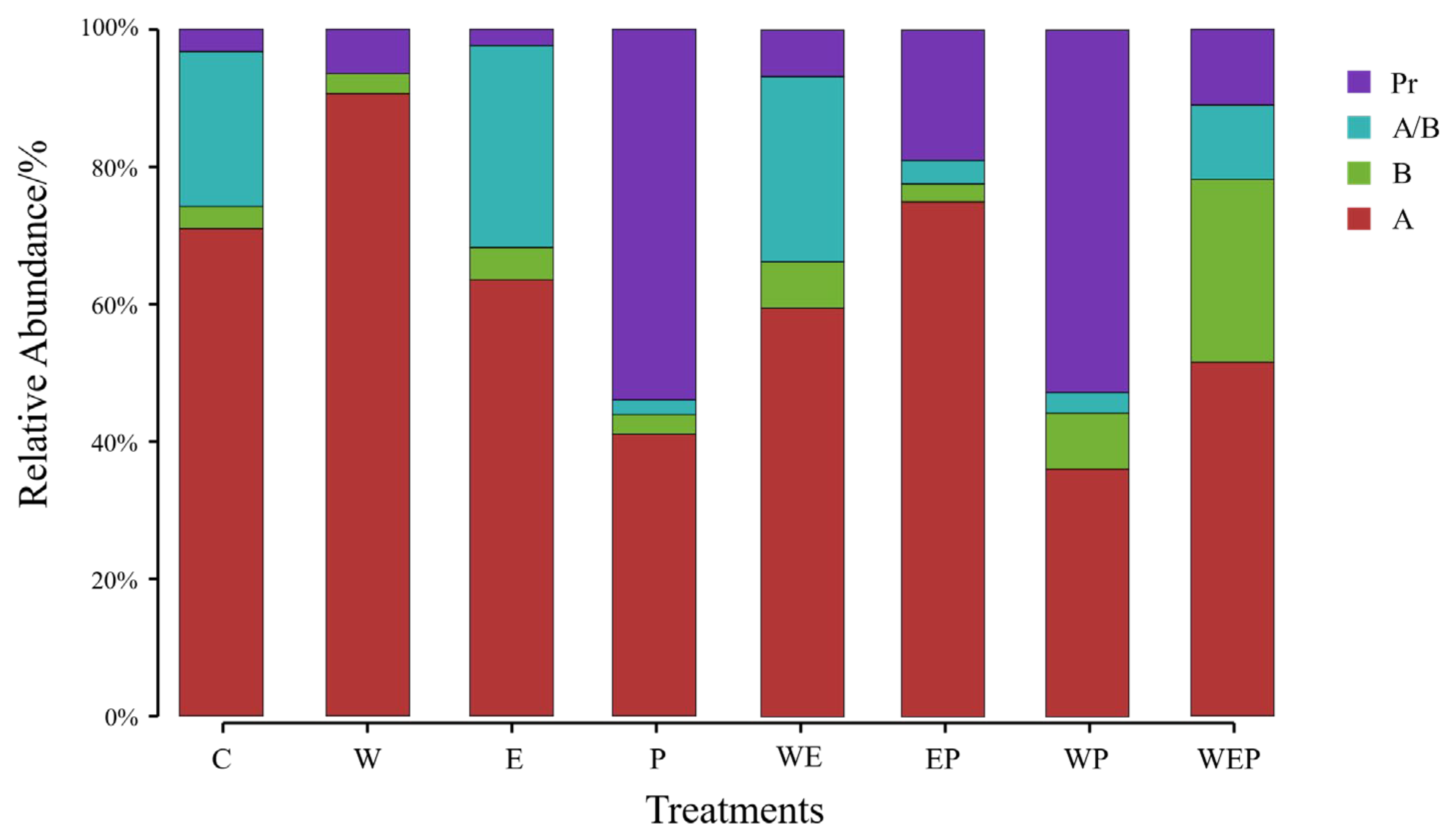
3.3. Effects of Different Treatments on Functional Group Composition of Protozoa
4. Discussion
4.1. Effects of Warming, Eutrophication, and Pesticide Pollution on the Community Structure of Protozoa
4.2. Effects of Warming, Eutrophication, and Pesticide Pollution on Protozoan Functional Groups
4.3. Study Limitations and Directions for Future Exploration
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Cambridge University Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Winder, M.; Schindler, D.E. Climate change uncouples trophic interactions in an aquatic ecosystem. Ecology 2004, 85, 2100–2106. [Google Scholar] [CrossRef]
- Winder, M.; Schindler, D.E. Climatic effects on the phenology of lake processes. Glob. Chang. Biol. 2004, 10, 1844–1856. [Google Scholar] [CrossRef]
- Kratina, P.; Watt, T.J.; Green, D.S.; Kordas, R.L.; O’Gorman, E.J. Interactive effects of warming and microplastics on metabolism but not feeding rates of a key freshwater detritivore. Environ. Pollut. 2019, 255, 113259. [Google Scholar] [CrossRef] [PubMed]
- Conley, D.J.; Paerl, H.W.; Howarth, R.W. Controlling eutrophication: Nitrogen and phosphorus. Science 2009, 323, 1014–1015. [Google Scholar] [CrossRef] [PubMed]
- Smith, V.H. Eutrophication of freshwater and coastal marine ecosystems a global problem. Environ. Sci. Pollut. Res. 2003, 10, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ma, F.; Xu, W.; Wang, J.; Zhu, J.; Li, Y.; Wu, Z. Study progress of steady-state transformation theory of phytoplankton and macrophyte in shallow lakes. Express Water Resour. Hydropower Inf. 2022, 43, 121–127. [Google Scholar]
- Bonmatin, J.M.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; Long, E.; Marzaro, M.; Mitchell, E.A.D.; et al. Environmental fate and exposure; neonicotinoids and fipronil. Environ. Sci. Pollut. Res. 2015, 22, 35–67. [Google Scholar] [CrossRef] [PubMed]
- Smit, C.E.; Posthuma-Doodeman, C.J.A.M.; Van Vlaardingen, P.L.A.; De Jong, F.M.W. Ecotoxicity of imidacloprid to aquatic organisms: Derivation of water quality standards for peak and long-term exposure. Hum. Ecol. Risk Assess. 2015, 21, 1608–1630. [Google Scholar] [CrossRef]
- Chará-Serna, A.M.; Epele, L.B.; Morrissey, C.A.; Richardson, J.S. Nutrients and sediment modify the impacts of a neonicotinoid insecticide on freshwater community structure and ecosystem functioning. Sci. Total Environ. 2019, 692, 1291–1303. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, P.Y.; Molinos, J.G.; Xie, J.Y.; Zhang, H.; Wang, H.; Xu, X.Q.; Wang, K.; Feng, M.J.; Cheng, H.W.; et al. Interactions between climate warming, herbicides, and eutrophication in the aquatic food web. J. Environ. Manag. 2023, 345, 118753. [Google Scholar] [CrossRef]
- Parry, J.D. Protozoan grazing of freshwater biofilms. Adv. Appl. Microbiol. 2004, 54, 167–196. [Google Scholar] [PubMed]
- Reisser, W. Endosymbiotic associations of freshwater protozoa and algae. Prog. Protistol. 1986, 1, 195–214. [Google Scholar]
- Kathol, M.; Fischer, H.; Weitere, M. Contribution of biofilm-dwelling consumers to pelagic-benthic coupling in a large river. Freshw. Biol. 2011, 56, 1017–1230. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, W.; Jiang, Y.; Zhu, M.; Al-Rasheid, K.A.S. An approach to analyzing influence of enumeration time periods on detecting ecological features of microperiphyton communities for marine bioassessment. Ecol. Indic. 2012, 18, 50–57. [Google Scholar] [CrossRef]
- Kiss, Á.K.; Ács, É.; Kiss, K.T.; Török, J.K. Structure and seasonal dynamics of the protozoan community (heterotrophic flagellates, ciliates, amoeboid protozoa) in the plankton of a large river (River Danube, Hungary). Eur. J. Protistol. 2009, 45, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Kathol, M.; Norf, H.; Arndt, H.; Weitere, M. Effects of temperature increase on the grazing of planktonic bacteria by biofilm-dwelling consumers. Aquat. Microb. Ecol. 2009, 55, 65–79. [Google Scholar] [CrossRef]
- Tan, X.; Shi, X.; Liu, G.; Xu, H.; Nie, P. An approach to analyzing taxonomic patterns of protozoan communities for monitoring water quality in Songhua River, northeast China. Hydrobiologia 2010, 638, 193–201. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, W.; Jiang, Y.; Zhu, M.; Al-Rasheid, K.A.S.; Warren, A.; Song, W. An approach to determining the sampling effort for analyzing biofilm-dwelling ciliate colonization using an artificial substratum in coastal waters. Biofouling 2011, 27, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Montagnes, D.; Roberts, E.; Luke, S.J.; Lowe, C. The rise of model protozoa. Trends Microbiol. 2012, 20, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Wagener, S. Abundance and distribution of anaerobic protozoa and their contribution to methane production in Lake Cadagno (Switzerland). FEMS Microbiol. Lett. 2017, 74, 39–48. [Google Scholar] [CrossRef]
- Jiang, J.; Wu, S.; Shen, Y. Effects of seasonal succession and water pollution on the protozoan community structure in a eutrophic lake. Chemosphere 2007, 66, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Foissner, W. Protists as bioindicators in activated sludge: Identification, ecology and future needs. Eur. J. Protistol. 2016, 55, 75–94. [Google Scholar] [CrossRef] [PubMed]
- Shukla, U.; Gupta, P.K. Assemblage of ciliated protozoan community in a polluted and non-polluted environment in a tropical lake of central himalaya: Lake Naini Tal, India. J. Plankton Res. 2001, 23, 571–584. [Google Scholar] [CrossRef]
- Pratt, J.R.; Cairns, J. Functional groups in the protozoa: Roles in differing ecosystems1,2. J. Protozool. 1985, 32, 415–423. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, Z.; Gong, X.; Yan, M.; Shi, Z.; Wei, Y. Modern Biomonitoring Techniques Using Freshwater Microbiota; China Architecture & Building Press: Beijing, China, 1992; pp. 152–175. [Google Scholar]
- Chen, Q.; Wu, L. Relationship between zooplankton functional groups and phytoplankton in summer in Chaohu Lake. J. Anhui Univ. Sci. Technol. (Nat. Sci.) 2021, 41, 42–50. [Google Scholar]
- Zhang, J.; Liu, Z.; Li, J.; Guo, N.; Liang, Y.; Wu, L. Spatial-temporal variations of planktonic crustacean functional groups and analysis for the impact factors in Chaohu Lake. J. Anhui Agric. Univ. 2023, 50, 463–472. [Google Scholar]
- Wang, S.; Shi, Z.; Geng, H.; Wu, L.; Cao, Y. Effects of environmental factors on the functional diversity of crustacean zooplankton community. J. Lake Sci. 2021, 33, 1220–1229. [Google Scholar]
- Liu, Z.; Wang, Y.; Zhao, C.; Chang, C.; Dou, H.; Chen, X.; Tao, Y.; Ma, C. The relationship between the characteristics of zooplankton functional groups and water environmental factors in Hulun Lake. J. Dalian Ocean. Univ. 2023, 38, 689–697. [Google Scholar]
- Liu, H.; Ma, L.; Min, X.; An, L.; Sang, J.; Ma, X.; Luo, C.; Min, X.; Jin, P. Analysis of community structure and functional-trophic group of Protozoa in Gahai Lake in Winter. Ecol. Sci. 2012, 31, 631–635. [Google Scholar]
- Qin, B.; Gao, G.; Zhu, G.; Zhang, Y.; Song, Y.; Tang, X.; Xu, H.; Deng, J. Lake eutrophication and its ecosystem response. Chin. Sci. Bull. 2013, 58, 961–970. [Google Scholar] [CrossRef]
- Hu, B.; Pan, M.; Shi, P.; Xu, J.; Zhang, M. Effect of climate warming and eutrophication on N2O flux at water-air interface of shallow lakes. Acta Hydrobiol. Sin. 2021, 45, 625–635. [Google Scholar]
- Yu, Q.; Wang, H.Z.; Jeppesen, E. Reply to Cao et al.’s comment on “Does the responses of Vallisneria natans (Lour.) Hara to high nitrogen loading differ between the summer high-growth season and the low-growth season? Sci. Total Environ. 2018, 615, 1093–1094. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, P.; Wang, H.; Molinos, J.G.; Xu, J. Synergistic effects of warming and eutrophication alert zooplankton predator-prey interactions along the benthic-pelagic interface. Glob. Chang. Biol. 2021, 27, 5907–5919. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Su, G.; Zhang, P.; Wang, T.; Zhao, K.; Zhang, H.; Huang, J.; Wang, H.; Kong, X.; Xu, J.; et al. Effects of multiple environmental stressors on zoobenthos communities in shallow lakes: Evidence from a mesocosm experiment. Animals 2023, 13, 3722. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, H.; Yan, S.; Wang, T.; Zhang, P.; Zhang, H.; Wang, H.; Hansson, L.S.; Xu, J. Chemo diversity of cyanobacterial toxins driven by future scenarios of climate warming and eutrophication. Environ. Sci. Technol. 2023, 55, 11767–11778. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xu, J.; Molinos, J.G. A dynamic temperature difference control recording system in shallow lake mesocosm. Methods X 2020, 7, 100930. [Google Scholar] [CrossRef] [PubMed]
- HJ 897-2017; National Environmental Protection Standard of the People’s Republic of China: Water Quality, Determination of Chlorophylla by Spectrophotometric Method. China Environment Press: Beijing, China, 2017.
- Foissner, W.; Berger, H. A user-friendly guide to the ciliates (Protozoa, Ciliophora) commonly used by hydrobiologists as bioindicators in rivers, lakes, and waste waters, with notes on their ecology. Freshw. Biol. 1996, 35, 375–482. [Google Scholar] [CrossRef]
- Hang, X.F. Quantitative methods for freshwater zooplankton. Reserv. Fish. 1982, 4, 52–59. [Google Scholar]
- Bai, H.; Wang, Y.; Song, J.; Xu, W.; Wu, Q.; Zhang, Y. Spatio-temporal characteristics and driving factors of zooplankton community structure in the Shaanxi section of Weihe River, China. Chin. J. Ecol. 2022, 41, 1602–1610. [Google Scholar]
- Zhang, P.; Wang, T.; Zhang, H. Heat waves rather than continuous warming exacerbate impacts of nutrient loading and herbicides on aquatic ecosystems. Environ. Int. 2022, 168, 107478. [Google Scholar] [CrossRef]
- He, M. Effects of Climate Warming and Water Eutrophication on Zooplankton Community Structure in Shallow Lakes. Master’s Thesis, South-Central Minzu University, Wuhan, China, 2019. [Google Scholar]
- Spilling, K.; Tedesco, L.; Klais, R.; Olli, K. Changing plankton communities: Causes, effects and consequences. Front. Mar. Sci. 2019, 6, 00272. [Google Scholar] [CrossRef]
- López-Abbate, M.C. Microzooplankton communities in a Changing Ocean: A risk assessment. Diversity 2021, 13, 82. [Google Scholar] [CrossRef]
- De Garidel-Thoron, T.; Chaabane, S.; Giraud, X.; Meilland, J.; Jonkers, L.; Kucera, M.; Brummer, G.J.A.; Grigoratou, M.; Monteiro, F.M.; Greco, M.; et al. Theforaminiferal response to climate stressors project: Tracking the community response of planktonic foraminifera to historical climate change. Front. Mar. Sci. 2022, 9, 827962. [Google Scholar] [CrossRef]
- Allen, A.P.; Gillooly, J.F.; Brown, J.H. Linking the global carbon cycle to individual metabolism. Funct. Ecol. 2005, 19, 202–213. [Google Scholar] [CrossRef]
- Rose, J.M.; Caron, D.A. Does low temperature constrain the growth rates of heterotrophic protists? Evidence and implications for algal blooms in cold waters. Limnol. Oceanogr. 2007, 52, 886–895. [Google Scholar] [CrossRef]
- Meunier, M.; Danelian, T. No dramatic changes observed in subtropical radiolarian plankton assemblages during the Middle Eocene Climatic Optimum (MECO); evidence from the North Atlantic ODP Site 1051. Mar. Micropaleontol. 2023, 184, 102272. [Google Scholar] [CrossRef]
- Molina-Grima, E.; García-Camacho, F.; Acién-Fernández, F.G.; Sánchez-Mirón, A.; Plouviez, M.; Shene, C.; Chisti, Y. Pathogens and predators impacting commercial production of microalgae and cyanobacteria. Biothchnol. Adv. 2022, 55, 107884. [Google Scholar] [CrossRef] [PubMed]
- Kratina, P.; Greig, H.S.; Thompson, P.L. Warming modifies trophic cascades and eutrophication in experimental freshwater communities. Ecology 2012, 93, 1421–1430. [Google Scholar] [CrossRef]
- Wilken, S.; Huisman, J.; Naus-wiezer, S.; Donk, E.V. Mixotrophic organisms become more heterotrophic with rising temperature. Ecol. Lett. 2013, 16, 225–233. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, L.; Zhu, X.X.; Wang, J.; Montagnes, D.J.S. An evidence-based framework for predicting the impact of differing autotroph-heterotroph thermal sensitivities on consumer-prey dynamics. ISME J. 2016, 10, 1767–1778. [Google Scholar] [CrossRef]
- Xu, R.L. Study on Protozoan Productions in Donghu Lake; Knowledge output from Institute of Hydrology, Chinese Academy of Sciences: Wuhan, China, 1988. [Google Scholar]
- Zhang, L.; Wang, Z.; Wang, N.; Gu, L.; Yang, Z. Mixotrophic ochromonas addition improves the harmful Microcystis-dominated phytoplankton community in in-situ microcosms. Environ. Sci. Technol. 2020, 54, 4609–4620. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Li, G.; Huang, R.; Cui, S.; Diao, X. Initial study on acute toxicity of two typical persistent organic pollutants (POPs) against Paramecium caudatum. Asian J. Ecotoxicol. 2011, 6, 37–42. [Google Scholar]
- Gutierrez, M.F.; Battauz, Y.; Caisso, B. Disruption of the hatching dynamics of zooplankton egg banks due to glyphosate application. Chemosphere 2017, 171, 644–653. [Google Scholar] [CrossRef]
- Tresnakova, N.; Stara, A.; Velisek, J. Effects of glyphosate and its metabolite AMPA on aquatic organisms. Appl. Sci. 2021, 11, 9004. [Google Scholar] [CrossRef]
- Hou, C.Y.; Shi, T.Y.; Wang, W.Y. Toxicological sensitivity of protozoa to pesticides and nanomaterials: A prospect review. Chemosphere 2023, 339, 139749. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Pan, X. Study on toxicity of different pesticides to unicellular animals-Paramecium. J. Zhoukou Norm. Univ. 2019, 36, 70–73. [Google Scholar]
- Wang, Z.; Huang, W.; Liu, Z.; Zeng, J.; He, Z.; Shu, L. The neonicotinoid insecticide imidacloprid has unexpected effects on the growth and development of soil amoebae. Sci. Total Environ. 2023, 869, 161884. [Google Scholar] [CrossRef] [PubMed]
- Tomizawa, M.; Millar, N.S.; Casida, J.E. Pharmacological profiles of recombinant and native insect nicotinic acetylcholine receptors. Insect Biochem. Mol. Biol. 2005, 35, 1347–1355. [Google Scholar] [CrossRef] [PubMed]
- Camp, A.A.; Buchwalter, D.B. Can’t take the heat: Temperature-enhanced toxicity in the mayfly Isonychia bicolor exposed to the neonicotinoid insecticide imidacloprid. Aquat. Toxicol. 2016, 178, 49–57. [Google Scholar] [CrossRef]
- Sumon, K.A.; Ritika, A.K.; Peeters, E.T.H.M.; Rashid, H.; Bosma, R.H.; Rahman, M.S.; Fatema, M.K.; Van den Brink, P.J. Effects of imidacloprid on the ecology of sub-tropical freshwater microcosms. Environ. Pollut. 2018, 236, 432–441. [Google Scholar] [CrossRef]
- Gao, Y.; He, J.; Chen, M.; Li, L.; Zhang, F. The spatial distribution of abundance and production of bacterioplankton in and near Chukchi Sea, Arctic Ocean. Haiyang Xuebao 2015, 37, 96–104. [Google Scholar]
- Shen, Y. Bacterial Dominated Population Abundance of Lake Taihu, Detected by FISH. Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, 2011. [Google Scholar]
- Bohatová, M.; Vdacny, P. Locomotory behaviour of two phylogenetically distant predatory ciliates: Does evolutionary history matter? Ethol. Ecol. Evol. 2018, 30, 195–219. [Google Scholar] [CrossRef]
- Lou, W.; Ni, B.; Yu, T.; Gu, F. The acute toxicity effect of acephate to Paramecium caudatum and Paramecium bursaria. J. Fudan Univ. (Nat. Sci.) 2008, 47, 370–373+386. [Google Scholar]
- Van, D.P.D.; Kai, S.Y.; Dan, L.; Hao, J.L.; Paul, J.; Van, D.B.; Guang, G.Y. Imidacloprid treatments induces cyanobacteria blooms in freshwater communities under sub-tropical conditions. Aquat. Toxicol. 2021, 240, 105992. [Google Scholar]
- Huang, H.; Liu, H.; Wang, G.; Han, D.; Zhang, H.; Dong, X.; Zhang, X. Study progress of the toxicity, detection methods and residues of carbamate insecticides in aquatic environment. Chin. Fish. Qual. Stand. 2016, 6, 23–30. [Google Scholar]
- Liu, Z.; Tang, Y. Acute toxicity of 5 kinds of insecticides to the 3 fresh-water aquatic zooplanktons. J. Saf. Environ. 2017, 17, 793–799. [Google Scholar]
- DeLong, J.P.; Al-Ameeli, Z.; Lyon, S.; Van Etten, J.L.; Dunigan, D.D. Size-dependent catalysis of chlorovirus population growth by a messy feeding predator. Microb. Ecol. 2018, 75, 847–853. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | TN (mg/L) | TP (mg/L) | DO (mg/L) | pH | Cond. (μS/cm) |
|---|---|---|---|---|---|
| C | 1.66 ± 0.58 | 0.03 ± 0.02 | 8.96 ± 0.51 | 7.56 ± 0.18 | 179.35 ± 18.86 |
| W | 1.98 ± 0.41 | 0.03 ± 0.02 | 8.51 ± 0.39 | 7.85 ± 0.51 | 214.48 ± 11.73 |
| E | 1.90 ± 0.70 | 0.09 ± 0.10 | 9.18 ± 0.49 | 8.11 ± 0.46 | 206.95 ± 20.94 |
| P | 1.71 ± 0.44 | 0.05 ± 0.04 | 9.87 ± 0.40 | 8.61 ± 0.67 | 159.35 ± 8.65 |
| WE | 2.47 ± 0.53 | 0.14 ± 0.14 | 8.11 ± 0.20 | 7.97 ± 0.56 | 254.17 ± 19.58 |
| EP | 3.98 ± 1.60 | 0.36 ± 0.20 | 9.38 ± 1.11 | 8.81 ± 0.65 | 206.95 ± 21.73 |
| WP | 1.78 ± 0.29 | 0.06 ± 0.04 | 8.54 ± 0.49 | 8.29 ± 0.45 | 187.47 ± 11.11 |
| WEP | 3.90 ± 0.87 | 0.35 ± 0.19 | 8.25 ± 2.55 | 8.96 ± 0.59 | 261.00 ± 13.29 |
| Scientific Name | Functional Group | Scientific Name | Functional Group |
|---|---|---|---|
| Vorticella sp. | B | Raphidiophrys sp. | A&B |
| Cyclidium sp. | B | Acanthocystis sp. | A&B |
| Strobilidium gyrans | A | Podophrya sp. | Pr |
| Strobilidium sp. | A | Prorodon sp. | A |
| Halteria sp. | B | Order Hymenostomata | N |
| Askenasia sp. | B | Holophrya sp. | B |
| Mesodinium sp. | B | Coleps sp. | A&B |
| Tintinnidium sp. | A&B | Order Prostomatida | N |
| Tintinnidium pusillum | A&B | Lagynaphrya conifera | A |
| Pseudodifflugia sp. | A | Cyrtolophosis sp. | A&B |
| Difflugia acuminata | A | Pseedoglaucoma sp. | B |
| Difflugia sp. | A | Lacrymaria sp. | Pr |
| Centropyxis aculeata | A&B | Dileptus sp. | B |
| Centropyxis hemisphaerica | B | Nassula sp. | A |
| Tintinopsis sp. | A&B | Trachelophyllum sp. | B |
| Tintinnopsis niei | A&B | Order Hypotrichida | N |
| Tintinnopsis wangi | A&B | Didinium sp. | Pr |
| Leprotintinnus sp. | A&B | Didinium balbianii nanum | Pr |
| Cyclopyxis eurostoma | A | Didinium nasutum | Pr |
| Arcella hemisphaerica | A&B | Litonotus sp. | Pr |
| Pontigulasia incisa | A | Urotricha sp. | A |
| Enchelys sp. | A&B | Trochilia palustris | A&B |
| Family Amoebidae | N | Strombidium sp. | Pr |
| Family Vahlkampfiidae | N | Actinobolina sp. | Pr |
| Nuclearia sp. | A&B | Aspidicca costatas | B |
| Treatments | Abundance | Biomass | ||||
|---|---|---|---|---|---|---|
| F | Df | p | F | Df | p | |
| W | 1.54 | 1 | 0.2198 | 1.01 | 1 | 0.3183 |
| E | 5.78 | 1 | 0.0195 * | 9.85 | 1 | 0.0027 ** |
| P | 8.67 | 1 | 0.0047 ** | 7.26 | 1 | 0.0093 ** |
| WE | 0.03 | 1 | 0.8664 | 4.22 | 1 | 0.0447 * |
| EP | 1.24 | 1 | 0.2695 | 0.01 | 1 | 0.9093 |
| WP | 2.81 | 1 | 0.0994 | 12.08 | 1 | 0.001 *** |
| WEP | 0.46 | 1 | 0.4984 | 1.95 | 1 | 0.1682 |
| Treatments | Simpson Index | Shannon-Wiener Index | Richness Index | Pielou Index | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | Df | p | F | Df | p | F | Df | p | F | Df | p | |
| W | 0.11 | 1 | 0.7380 | 0.69 | 1 | 0.4088 | 2.47 | 1 | 0.1214 | 1.54 | 1 | 0.2194 |
| E | 9.37 | 1 | 0.0034 ** | 11.24 | 1 | 0.0014 ** | 28.18 | 1 | <0.001 *** | 5.20 | 1 | 0.0264 * |
| P | 1.00 | 1 | 0.3227 | 0.07 | 1 | 0.7925 | 0.79 | 1 | 0.3783 | 9.07 | 1 | 0.0039 ** |
| WE | 0.03 | 1 | 0.8538 | 0.01 | 1 | 0.9161 | 0.39 | 1 | 0.5358 | 0.18 | 1 | 0.6711 |
| EP | 0.52 | 1 | 0.4749 | 0.01 | 1 | 0.9192 | 0.04 | 1 | 0.8431 | 4.12 | 1 | 0.0471 * |
| WP | 3.98 | 1 | 0.0509 | 5.53 | 1 | 0.0222 * | 6.19 | 1 | 0.0158 * | 6.52 | 1 | 0.0134 * |
| WEP | 0.04 | 1 | 0.8338 | 0.05 | 1 | 0.8291 | 0.43 | 1 | 0.5147 | 0.14 | 1 | 0.7104 |
| Functional Group | A | A | B | B | A&B | A&B | A&B | Pr | Pr |
|---|---|---|---|---|---|---|---|---|---|
| Dominant Species | Strobilidium sp. | Difflugia sp. | Cyclidium sp. | Halteria sp. | Cyrtolophosis sp. | Tintinnidium sp. | Tintinopsis sp. | Didinium sp. | Podophrya sp. |
| April | + | + | + | ||||||
| May | + | + | |||||||
| June | + | + | + | ||||||
| July | + | + | + | ||||||
| August | + | + | + | ||||||
| September | + | + | + | + | + | ||||
| October | + | + | + | + | |||||
| November | + | + | + | + | + |
| Functional Group | Scientific Name | Treatment | |||||||
|---|---|---|---|---|---|---|---|---|---|
| C | W | E | P | WE | EP | WP | WEP | ||
| A | Strobilidium sp. | 0.3826 | 0.7636 | 0.6667 | 0.4103 | 0.5951 | 0.7500 | 0.3605 | 0.5152 |
| B | Cyclidium sp. | 0.1818 | 0.0815 | 0.2662 | |||||
| A&B | Tintinopsis sp | 0.0696 | |||||||
| A&B | Tintinnidium sp. | 0.0522 | 0.2099 | 0.2699 | 0.1084 | ||||
| R | Didinium sp. | 0.5400 | 0.3691 | ||||||
| R | Podophrya sp. | 0.1034 | 0.1545 | ||||||
| Functional Group | A | B | A&B | Pr | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatments | F | Df | p | F | Df | p | F | Df | p | F | Df | p |
| W | 6.85 | 1 | 0.0151 * | 0.91 | 1 | 0.3494 | 0.56 | 1 | 0.4583 | 0.32 | 1 | 0.7463 |
| E | 3.69 | 1 | 0.0664 | 3.85 | 1 | 0.0612 | 0.06 | 1 | 0.8386 | 0.26 | 1 | 0.7968 |
| P | 7.88 | 1 | 0.0097 ** | 0.05 | 1 | 0.9369 | 6.86 | 1 | 0.0164 * | 8.88 | 1 | 0.0081 ** |
| WE | 2.08 | 1 | 0.1614 | 4.20 | 1 | 0.0513 | 0.79 | 1 | 0.3816 | 0.60 | 1 | 0.5489 |
| EP | 0.62 | 1 | 0.4386 | 0.61 | 1 | 0.4424 | 7.02 | 1 | 0.0158 * | 0.60 | 1 | 0.5532 |
| WP | 22.16 | 1 | <0.001 *** | 4.78 | 1 | 0.0386 * | 2.38 | 1 | 0.0255 * | 10.30 | 1 | 0.0029 ** |
| WEP | 0.21 | 1 | 0.6482 | 13.15 | 1 | 0.0013 ** | 2.28 | 1 | 0.1436 | 6.77 | 1 | 0.0105 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, G.; Chen, Y.; Wang, Y.; Zhang, H.; Wang, H.; Jiang, M.; Zhang, X.; Gong, Y.; Yuan, S. Responses of Protozoan Communities to Multiple Environmental Stresses (Warming, Eutrophication, and Pesticide Pollution). Animals 2024, 14, 1293. https://doi.org/10.3390/ani14091293
Yuan G, Chen Y, Wang Y, Zhang H, Wang H, Jiang M, Zhang X, Gong Y, Yuan S. Responses of Protozoan Communities to Multiple Environmental Stresses (Warming, Eutrophication, and Pesticide Pollution). Animals. 2024; 14(9):1293. https://doi.org/10.3390/ani14091293
Chicago/Turabian StyleYuan, Guoqing, Yue Chen, Yulu Wang, Hanwen Zhang, Hongxia Wang, Mixue Jiang, Xiaonan Zhang, Yingchun Gong, and Saibo Yuan. 2024. "Responses of Protozoan Communities to Multiple Environmental Stresses (Warming, Eutrophication, and Pesticide Pollution)" Animals 14, no. 9: 1293. https://doi.org/10.3390/ani14091293
APA StyleYuan, G., Chen, Y., Wang, Y., Zhang, H., Wang, H., Jiang, M., Zhang, X., Gong, Y., & Yuan, S. (2024). Responses of Protozoan Communities to Multiple Environmental Stresses (Warming, Eutrophication, and Pesticide Pollution). Animals, 14(9), 1293. https://doi.org/10.3390/ani14091293