
Differential Gene Expression in Porcine Lung Compartments after Experimental Infection with Mycoplasma hyopneumoniae
 , , , , , , and
, , , , , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Inoculum
2.2. Animals and Experimental Desing
2.3. Experimental Infection
2.4. Serum Sample Collection and Mhyo Specific ELISA
2.5. Gross Pathology and Microscopic Evaluation of Lung Lesions
2.6. Laser Capture Microdissection
2.7. DNA Extraction and Mhyo Quantification
2.8. RNA Extraction and qPCR Gene Expression Analysis
2.9. Statistical Analysis
3. Results
3.1. Mhyo Humoral Response Analysis
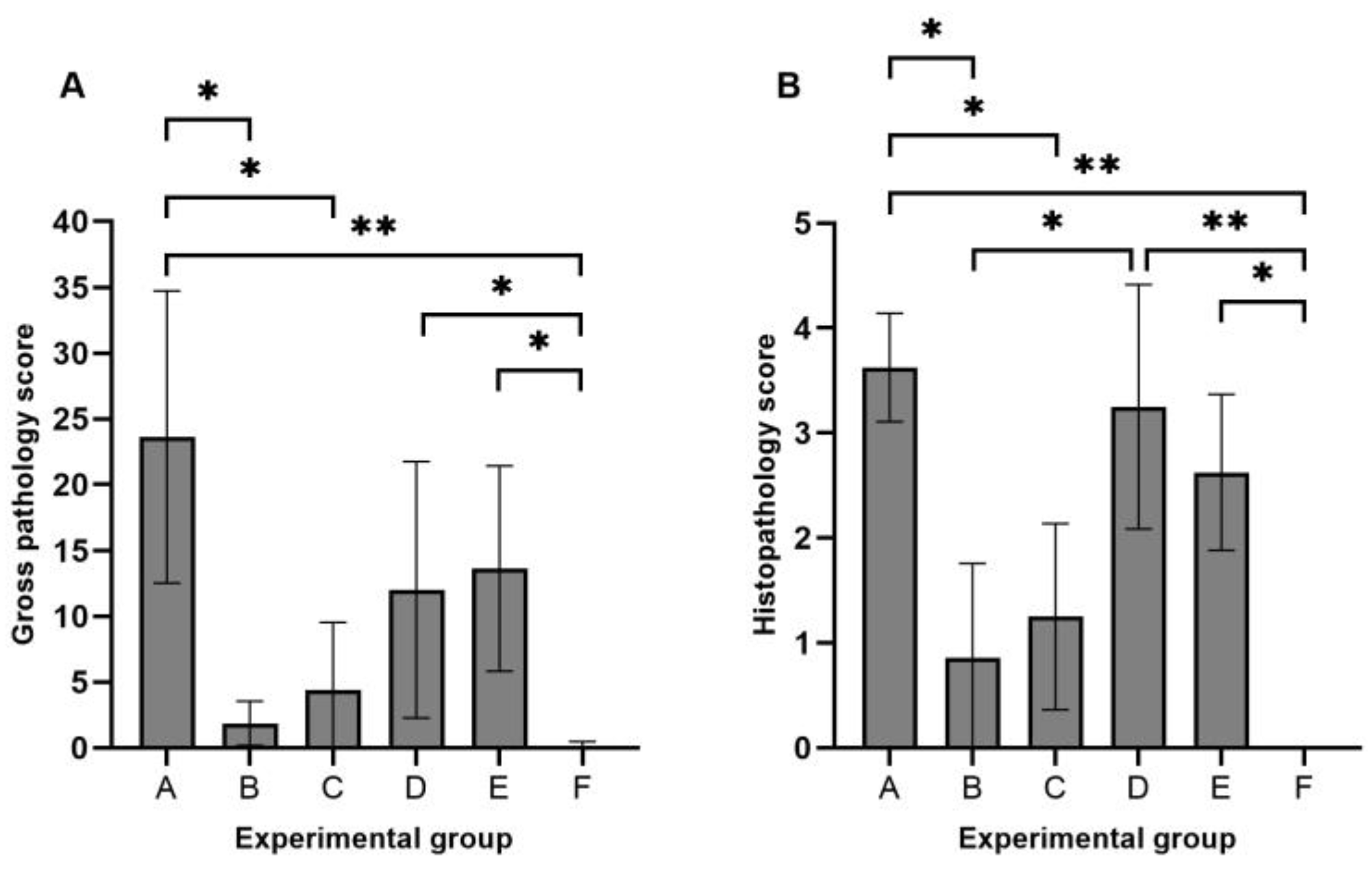
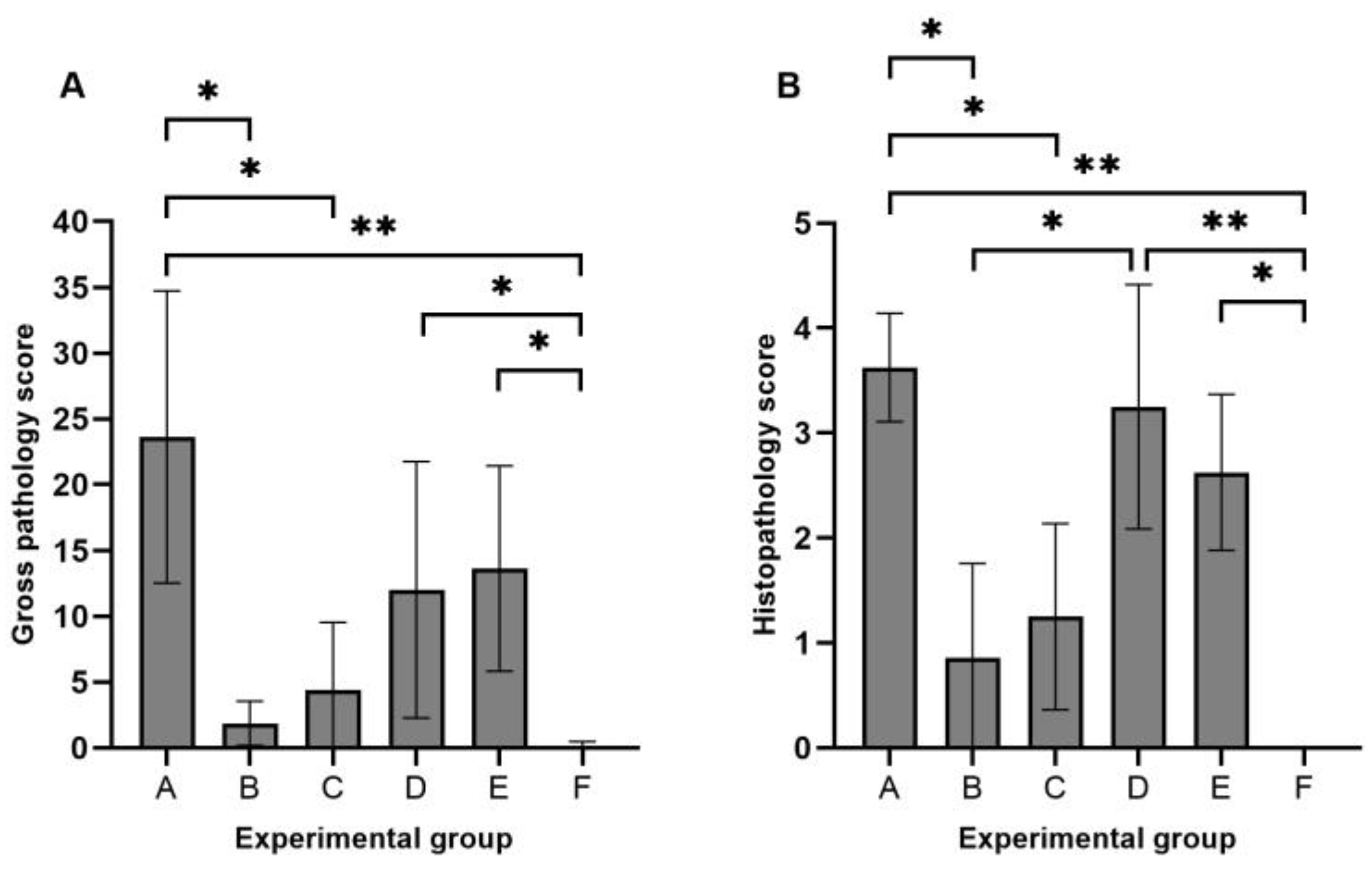
3.2. Gross Pathology and Histopathology
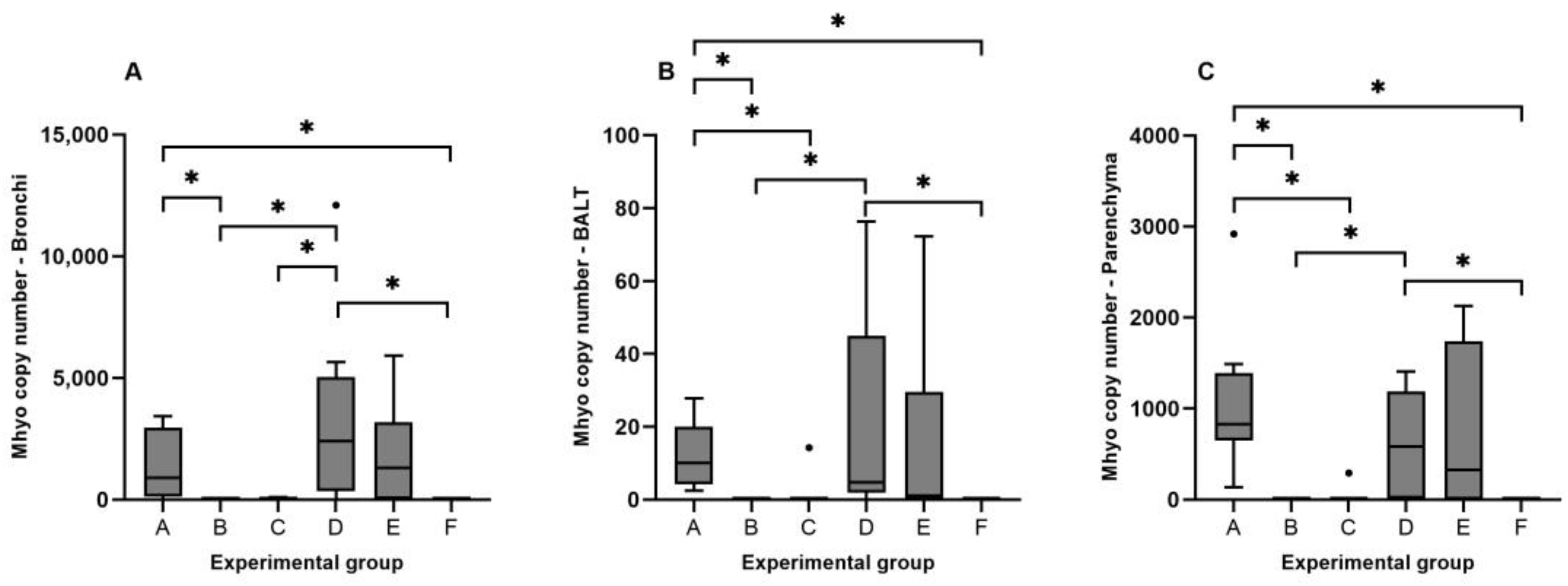
3.3. Detection of Mhyo Nucleic Acids by qPCR
3.4. Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pieters, M.G.; Maes, D. Mycoplasmosis. In Diseases of Swine; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; Wiley: Hoboken, NJ, USA, 2019; pp. 863–883. [Google Scholar]
- Holst, S.; Yeske, P.; Pieters, M. Elimination of Mycoplasma hyopneumoniae from Breed-to-Wean Farms: A Review of Current Protocols with Emphasis on Herd Closure and Medication. J. Swine Health Prod. 2015, 23, 321–330. [Google Scholar]
- DeBey, M.C.; Ross, R.F. Ciliostasis and Loss of Cilia Induced by Mycoplasma hyopneumoniae in Porcine Tracheal Organ Cultures. Infect Immun. 1994, 62, 5312–5318. [Google Scholar] [CrossRef] [PubMed]
- Bustamante-Marin, X.M.; Ostrowski, L.E. Cilia and Mucociliary Clearance. Cold Spring Harb. Perspect. Biol. 2017, 9, a028241. [Google Scholar] [CrossRef] [PubMed]
- Leal Zimmer, F.M.A.; Paes, J.A.; Zaha, A.; Ferreira, H.B. Pathogenicity & Virulence of Mycoplasma hyopneumoniae. Virulence 2020, 11, 1600–1622. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Oh, Y.; Han, K.; Seo, H.W.; Kim, D.; Park, C.; Kang, I.; Chae, C. Expression of Secreted and Membrane-Bound Mucins in the Airways of Piglets Experimentally Infected with Mycoplasma hyopneumoniae. Vet. J. 2012, 192, 120–122. [Google Scholar] [CrossRef]
- Raymond, B.B.A.; Djordjevic, S. Exploitation of Plasmin(Ogen) by Bacterial Pathogens of Veterinary Significance. Vet. Microbiol. 2015, 178, 1–13. [Google Scholar] [CrossRef]
- Bai, F.; Ni, B.; Liu, M.; Feng, Z.; Xiong, Q.; Shao, G. Mycoplasma hyopneumoniae-Derived Lipid-Associated Membrane Proteins Induce Inflammation and Apoptosis in Porcine Peripheral Blood Mononuclear Cells in Vitro. Vet. Microbiol. 2015, 175, 58–67. [Google Scholar] [CrossRef]
- Browning, G.F.; Marenda, M.S.; Noormohammadi, A.H.; Markham, P.F. The Central Role of Lipoproteins in the Pathogenesis of Mycoplasmoses. Vet. Microbiol. 2011, 153, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Livingston, C.W.; Stair, E.L.; Underdahl, N.R.; Mebus, C.A. Pathogenesis of Mycoplasmal Pneumonia in Swine. Am. J. Vet. Res. 1972, 33, 2249–2258. [Google Scholar]
- Huang, Y.T.; Chu, R.M.; Liu, R.S.; Weng, C.N. Morphologic Studies of Intrapulmonary Airway Mucosa-Associated Lymphoid Tissues in Swine. Vet. Immunol. Immunopathol. 1990, 25, 13–22. [Google Scholar] [CrossRef]
- Maes, D.; Sibila, M.; Kuhnert, P.; Segalés, J.; Haesebrouck, F.; Pieters, M. Update on Mycoplasma hyopneumoniae Infections in Pigs: Knowledge Gaps for Improved Disease Control. Transbound. Emerg. Dis. 2018, 65, 110–124. [Google Scholar] [CrossRef]
- Asai, T.; Okada, M.; Ono, M.; Irisawa, T.; Mori, Y.; Yokomizo, Y.; Sato, S. Increased Levels of Tumor Necrosis Factor and Interleukin 1 in Bronchoalveolar Lavage Fluids from Pigs Infected with Mycoplasma hyopneumoniae. Vet. Immunol. Immunopathol. 1993, 38, 253–260. [Google Scholar] [CrossRef]
- Asai, T.; Okada, M.; Ono, M.; Mori, Y.; Yokomizo, Y.; Sato, S. Detection of Interleukin-6 and Prostaglandin E2 in Bronchoalveolar Lavage Fluids of Pigs Experimentally Infected with Mycoplasma hyponeumoniae. Vet. Immunol. Immunopathol. 1994, 44, 97–102. [Google Scholar] [CrossRef]
- Thanawongnuwech, R.; Thacker, B.; Halbur, P.; Thacker, E.L. Increased Production of Proinflammatory Cytokines Following Infection with Porcine Reproductive and Respiratory Syndrome Virus and Mycoplasma hyopneumoniae. Clin. Diagn. Lab. Immunol. 2004, 11, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Muneta, Y.; Uenishi, H.; Kikuma, R.; Yoshihara, K.; Shimoji, Y.; Yamamoto, R.; Hamashima, N.; Yokomizo, Y.; Mori, Y. Porcine TLR2 and TLR6: Identification and Their Involvement in Mycoplasma hyopneumoniae Infection. J. Interferon. Cytokine Res. 2003, 23, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, F.; Ramírez, G.A.; Sarradell, J.; Andrada, M.; Lorenzo, H. Immunohistochemical Labelling of Cytokines in Lung Lesions of Pigs Naturally Infected with Mycoplasma hyopneumoniae. J. Comp. Pathol. 2004, 130, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Meyns, T.; Maes, D.; Calus, D.; Ribbens, S.; Dewulf, J.; Chiers, K.; de Kruif, A.; Cox, E.; Decostere, A.; Haesebrouck, F. Interactions of Highly and Low Virulent Mycoplasma hyopneumoniae Isolates with the Respiratory Tract of Pigs. Vet. Microbiol. 2007, 120, 87–95. [Google Scholar] [CrossRef]
- Garcia-Morante, B.; Segalés, J.; Fraile, L.; Llardén, G.; Coll, T.; Sibila, M. Potential Use of Local and Systemic Humoral Immune Response Parameters to Forecast Mycoplasma hyopneumoniae Associated Lung Lesions. PLoS ONE 2017, 12, e0175034. [Google Scholar] [CrossRef]
- Pieters, M.; Pijoan, C.; Fano, E.; Dee, S. An Assessment of the Duration of Mycoplasma hyopneumoniae Infection in an Experimentally Infected Population of Pigs. Vet. Microbiol. 2009, 134, 261–266. [Google Scholar] [CrossRef]
- Raymond, B.B.A.; Turnbull, L.; Jenkins, C.; Madhkoor, R.; Schleicher, I.; Uphoff, C.C.; Whitchurch, C.B.; Rohde, M.; Djordjevic, S.P. Mycoplasma hyopneumoniae Resides Intracellularly within Porcine Epithelial Cells. Sci. Rep. 2018, 8, 17697. [Google Scholar] [CrossRef]
- Tacchi, J.L.; Raymond, B.B.A.; Haynes, P.A.; Berry, I.J.; Widjaja, M.; Bogema, D.R.; Woolley, L.K.; Jenkins, C.; Minion, F.C.; Padula, M.P.; et al. Post-Translational Processing Targets Functionally Diverse Proteins in Mycoplasma hyopneumoniae. Open Biol. 2016, 6, 150210. [Google Scholar] [CrossRef] [PubMed]
- Bonner, R.F.; Emmert-Buck, M.; Cole, K.; Pohida, T.; Chuaqui, R.; Goldstein, S.; Liotta, L.A. Laser Capture Microdissection: Molecular Analysis of Tissue. Science 1997, 278, 1481–1483. [Google Scholar] [CrossRef] [PubMed]
- García-Nicolás, O.; Rosales, R.S.; Pallarés, F.J.; Risco, D.; Quereda, J.J.; Graham, S.P.; Frossard, J.-P.; Morgan, S.B.; Steinbach, F.; Drew, T.W.; et al. Comparative Analysis of Cytokine Transcript Profiles within Mediastinal Lymph Node Compartments of Pigs after Infection with Porcine Reproductive and Respiratory Syndrome Genotype 1 Strains Differing in Pathogenicity. Vet. Res. 2015, 46, 34. [Google Scholar] [CrossRef] [PubMed]
- Aranday-Cortes, E.; Bull, N.C.; Villarreal-Ramos, B.; Gough, J.; Hicks, D.; Ortiz-Peláez, A.; Vordermeier, H.M.; Salguero, F.J. Upregulation of IL-17A, CXCL9 and CXCL10 in Early-Stage Granulomas Induced by Mycobacterium bovis in Cattle. Transbound. Emerg. Dis. 2013, 60, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Minion, F.C.; Lefkowitz, E.J.; Madsen, M.L.; Cleary, B.J.; Swartzell, S.M.; Mahairas, G.G. The Genome Sequence of Mycoplasma hyopneumoniae Strain 232, the Agent of Swine Mycoplasmosis. J. Bacteriol. 2004, 186, 7123–7133. [Google Scholar] [CrossRef]
- Friis, N.F. Some Recommendations Concerning Primary Isolation of Mycoplasma suipneumoniae and Mycoplasma flocculare a Survey. Nord. Vet. Med. 1975, 27, 337–339. [Google Scholar] [PubMed]
- McAuliffe, L.; Ellis, R.J.; Lawes, J.R.; Ayling, R.D.; Nicholas, R.A. 16S RDNA PCR and Denaturing Gradient Gel Electrophoresis; a Single Generic Test for Detecting and Differentiating Mycoplasma Species. J. Med. Microbiol. 2005, 54, 731–739. [Google Scholar] [CrossRef]
- Goodwin, R.F.; Whittlestone, P. Enzootic Pneumonia of Pigs: Immunization Attempts Inoculating Mycoplasma suipneumoniae Antigen by Various Routes and with Different Adjuvants. Br. Vet. J. 1973, 129, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Marois, C.; Dory, D.; Fablet, C.; Madec, F.; Kobisch, M. Development of a Quantitative Real-Time TaqMan PCR Assay for Determination of the Minimal Dose of Mycoplasma hyopneumoniae Strain 116 Required to Induce Pneumonia in SPF Pigs. J. Appl. Microbiol. 2010, 108, 1523–1533. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhang, J.; Tang, Z.; Wang, N.; Long, L.; Li, K. Evaluating a Set of Reference Genes for Expression Normalization in Multiple Tissues and Skeletal Muscle at Different Development Stages in Pigs Using Quantitative Real-Time Polymerase Chain Reaction. DNA Cell Biol. 2012, 31, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Dawson, H.D.; Chen, C.; Gaynor, B.; Shao, J.; Urban, J.F. The Porcine Translational Research Database: A Manually Curated, Genomics and Proteomics-Based Research Resource. BMC Genom. 2017, 18, 643. [Google Scholar] [CrossRef] [PubMed]
- Dawood, A.; Algharib, S.A.; Zhao, G.; Zhu, T.; Qi, M.; Delai, K.; Hao, Z.; Marawan, M.A.; Shirani, I.; Guo, A. Mycoplasmas as Host Pantropic and Specific Pathogens: Clinical Implications, Gene Transfer, Virulence Factors, and Future Perspectives. Front. Cell Infect. Microbiol. 2022, 12, 855731. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, H.; Quesada, Ó.; Assunçao, P.; Castro, A.; Rodríguez, F. Cytokine Expression in Porcine Lungs Experimentally Infected with Mycoplasma hyopneumoniae. Vet. Immunol. Immunopathol. 2006, 109, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.; Kwon, D.; Jung, K.; Ha, Y.; Lee, Y.-H.; Kim, O.; Park, H.-K.; Kim, S.-H.; Hwang, K.-K.; Chae, C. Expression of Inflammatory Cytokines in Pigs Experimentally Infected with Mycoplasma hyopneumoniae. J. Comp. Pathol. 2006, 134, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Thanawongnuwech, R.; Thacker, E.L. Interleukin-10, Interleukin-12, and Interferon- γ Levels in the Respiratory Tract Following Mycoplasma hyopneumoniae and PRRSV Infection in Pigs. Viral Immunol. 2003, 16, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, F.; Rosales, R.S.; Ramírez, A.S.; Poveda, J.B. Vaccination Upregulates Th1 Cytokines in the Lung of Pigs Experimentally Infected with Mycoplasma hyopneumoniae. Animals 2023, 13, 520. [Google Scholar] [CrossRef]
- Almeida, H.M.S.; Mechler-Dreibi, M.L.; Sonálio, K.; Ferraz, M.E.S.; Storino, G.Y.; Barbosa, F.O.; Maes, D.; Montassier, H.J.; de Oliveira, L.G. Cytokine Expression and Mycoplasma hyopneumoniae Burden in the Development of Lung Lesions in Experimentally Inoculated Pigs. Vet. Microbiol. 2020, 244, 108647. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lunney, J.K.; Baker, R.B.; Opriessnig, T. Cytokine and Chemokine MRNA Expression Profiles in Tracheobronchial Lymph Nodes from Pigs Singularly Infected or Coinfected with Porcine Circovirus Type 2 (PCV2) and Mycoplasma hyopneumoniae (MHYO). Vet. Immunol. Immunopathol. 2011, 140, 152–158. [Google Scholar] [CrossRef]
- Bin, L.; Luping, D.; Bing, S.; Zhengyu, Y.; Maojun, L.; Zhixin, F.; Yanna, W.; Haiyan, W.; Guoqing, S.; Kongwang, H. Transcription Analysis of the Porcine Alveolar Macrophage Response to Mycoplasma hyopneumoniae. PLoS ONE 2014, 9, e101968. [Google Scholar] [CrossRef]
- Zhang, J.-M.; An, J. Cytokines, Inflammation, and Pain. Int. Anesth. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef]
- Christodoulides, A.; Gupta, N.; Yacoubian, V.; Maithel, N.; Parker, J.; Kelesidis, T. The Role of Lipoproteins in Mycoplasma-Mediated Immunomodulation. Front. Microbiol. 2018, 9, 1682. [Google Scholar] [CrossRef] [PubMed]
- Thanawongnuwech, R.; Young, T.F.; Thacker, B.J.; Thacker, E.L. Differential Production of Proinflammatory Cytokines: In Vitro PRRSV and Mycoplasma hyopneumoniae Co-Infection Model. Vet. Immunol. Immunopathol. 2001, 79, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Redondo, E.; Masot, A.J.; Fernández, A.; Gázquez, A. Histopathological and Immunohistochemical Findings in the Lungs of Pigs Infected Experimentally with Mycoplasma hyopneumoniae. J. Comp. Pathol. 2009, 140, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Hu, W.; Wei, Y.; Feng, Z.; Yang, Q. Effects of Mycoplasma hyopneumoniae on Porcine Nasal Cavity Dendritic Cells. Vet. Microbiol. 2017, 198, 1–8. [Google Scholar] [CrossRef]
- Fan, F.; Lv, J.; Yang, Q.; Jiang, F. Clinical Characteristics and Serum Inflammatory Markers of Community-Acquired Mycoplasma Pneumonia in Children. Clin. Respir. J. 2023, 17, 607–617. [Google Scholar] [CrossRef]
- Morrison, D.F.; Foss, D.L.; Murtaugh, M.P. Interleukin-10 Gene Therapy-Mediated Amelioration of Bacterial Pneumonia. Infect. Immun. 2000, 68, 4752–4758. [Google Scholar] [CrossRef]
- Beuckelaere, L.; Haspeslagh, M.; Biebaut, E.; Boyen, F.; Haesebrouck, F.; Krejci, R.; Meyer, E.; Gleerup, D.; De Spiegelaere, W.; Devriendt, B.; et al. Different Local, Innate and Adaptive Immune Responses Are Induced by Two Commercial Mycoplasma hyopneumoniae Bacterins and an Adjuvant Alone. Front. Immunol. 2022, 13, 1015525. [Google Scholar] [CrossRef]
- Vignali, D.A.A.; Kuchroo, V.K. IL-12 Family Cytokines: Immunological Playmakers. Nat. Immunol. 2012, 13, 722–728. [Google Scholar] [CrossRef]
- Wu, Q.; Martin, R.J.; Rino, J.G.; Breed, R.; Torres, R.M.; Chu, H.W. IL-23-Dependent IL-17 Production Is Essential in Neutrophil Recruitment and Activity in Mouse Lung Defense against Respiratory Mycoplasma pneumoniae Infection. Microbes Infect. 2007, 9, 78–86. [Google Scholar] [CrossRef]
- Sadler, A.J.; Williams, B.R.G. Interferon-Inducible Antiviral Effectors. Nat. Rev. Immunol. 2008, 8, 559–568. [Google Scholar] [CrossRef]
- Hasslung, F.C.; Berg, M.; Allan, G.M.; Meehan, B.M.; McNeilly, F.; Fossum, C. Identification of a Sequence from the Genome of Porcine Circovirus Type 2 with an Inhibitory Effect on IFN-Alpha Production by Porcine PBMCs. J. Gen. Virol. 2003, 84, 2937–2945. [Google Scholar] [CrossRef]
- Li, P.; Zhang, Y.; Li, X.; Zhou, W.; Li, X.; Jiang, F.; Wu, W. Mycoplasma hyopneumoniae Mhp597 Is a Cytotoxicity, Inflammation and Immunosuppression Associated Nuclease. Vet. Microbiol. 2019, 235, 53–62. [Google Scholar] [CrossRef]
- Kang, S.; Brown, H.M.; Hwang, S. Direct Antiviral Mechanisms of Interferon-Gamma. Immune Netw. 2018, 18, e33. [Google Scholar] [CrossRef]
- Ider, M.; Maden, M. Biomarkers of Infectious Pneumonia in Naturally Infected Calves. Am. J. Vet. Res. 2022, 83, 1–8. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Probe (5′-3′) | Reference |
|---|---|---|---|---|
| ICAM1 | GGACCATGGGCGAAGCTT | TGGGCAATCCCTCTCGTCTA | [6FAM]-ACATGCCCAGCCACCGTCAGG-[TAM] | [33] |
| IL-8 | CCGTGTCAACATGACTTCCAA | GCCTCACAGAGAGCTGCAGAA | [6FAM]-CTGTTGCCTTCTTGGCAGTTTTCCTGC-[TAM] | [33] |
| IL-10 | TGAGAACAGCTGCATCCACTTC | TCTGGTCCTTCGTTTGAAAGAAA | [6FAM]-CAACCAGCCTGCCCCACATGC-[TAM] | [33] |
| IL-23 | AGAAGAGGGAGATGATGAGAC | AGCAGGACTGACTGCCGTCC | [6FAM]-CTGAGGATCACAGCCATCCCCGC-[TAM] | [24] |
| IFN-α | TCAGCTGCAATGCCATCTG | AGGGAGAGATTCTCCTCATTTGTG | [6FAM]-TGACCTGCCTCAGACCCACAGCC-[TAM] | [33] |
| IFN-γ | GAAAAGCTGATTAAATTCCGGTAG | AGGTTAGATCTTGGTGACAGATC | [6FAM]-TCTGCAGATCCAGCGCAAAGCCATCAG-[TAM] | [24] |
| TGF-β | AGGGCTACCATGCCAATTT | CCGGGTTGTGCTGGTTGT | [6FAM]-CACTCAGTACAGCAAGGTCCTGGCTCTGTA-[TAM] | [33] |
| TNF-α | TGGCCCCTTGAGCATCA | CGGGCTTATCTGAGGTTTGAGA | [6FAM]-CCCTCTGGCCCAAGGACTCAGATCA-[TAM] | [33] |
| Rpl32 | TGGAAGAGACGTTGTGAGCAA | CGGAAGTTTCTGGTACACAATGTAA | [6FAM]-ATTTGTTGCACATTAGCAGCACTTCAAGCTC-[TAM] | [33] |
| Gross Pathology Score | Histopathology Score | Mhyo Load—Bronchi | Mhyo Load—BALT | Mhyo Load—Parenchyma | |
|---|---|---|---|---|---|
| Gross pathology score | 0.901 | 0.699 | 0.733 | 0.75 | |
| Histopathology score | 0.901 | 0.769 | 0.753 | 0.791 | |
| Mhyo load—bronchi | 0.699 | 0.769 | 0.68 | 0.732 | |
| Mhyo load- BALT | 0.733 | 0.753 | 0.68 | 0.879 | |
| Mhyo load—parenchyma | 0.75 | 0.791 | 0.732 | 0.879 |
| ICAM1 | IL-8 | IL-10 | IL-23 | IFN-α | IFN-γ | TGF-β | TNF-α | ||
|---|---|---|---|---|---|---|---|---|---|
| Mhyo load | BR | 0.147 | 0.663 ** | 0.194 | −0.215 | −0.294 | −0.279 | −0.028 | −0.373 * |
| BT | 0.065 | 0.329 | 0.037 | −0.156 | −0.05 | 0.381 | 0.273 | −0.077 | |
| PA | −0.323 | 0.663 ** | 0.099 | −0.483 ** | −0.571 ** | −0.022 | −0.131 | −0.241 | |
| ICAM1 | BR | 0.380 * | 0.514 ** | 0.350 * | 0.285 | 0.399 * | 0.741 ** | 0.559 ** | |
| BT | 0.028 | −0.259 | 0.097 | 0.534 * | 0.206 | 0.578 * | 0.544 * | ||
| PA | 0.018 | 0.2 | 0.497 ** | 0.633 ** | 0.496 ** | 0.647 ** | 0.726 ** | ||
| IL-8 | BR | 0.514 ** | 0.350 * | 0.285 | 0.399 * | 0.741 ** | 0.559 * | ||
| BT | −0.383 | −0.318 | 0.058 | −0.258 | 0.061 | −0.548 | |||
| PA | 0.192 | −0.059 | −0.268 | −0.008 | 0.054 | −0.06 | |||
| IL-10 | BR | 0.374 | −0.071 | 0.165 | 0.412 * | 0.269 | |||
| BT | 0.168 | −0.274 | −0.059 | 0.009 | −0.106 | ||||
| PA | 0.202 | −0.065 | 0.610 ** | 0.371 * | 0.463 ** | ||||
| IL-23 | BR | 0.362 * | 0.15 * | 0.405 * | 0.706 ** | ||||
| BT | 0.606 * | 0.041 | 0.512 * | 0.021 | |||||
| PA | 0.512 ** | 0.294 | 0.540 ** | 0.462 * | |||||
| IFN-α | BR | 0.314 | 0.392 * | 0.497 ** | |||||
| BT | −0.015 | 0.446 | 0.243 | ||||||
| PA | 0.169 | 0.474 ** | 0.485 ** | ||||||
| IFN-γ | BR | 0.519 ** | 0.735 ** | ||||||
| BT | 0.211 | 0.608 ** | |||||||
| PA | 0.517 ** | 0.783 ** | |||||||
| TGF-β | BR | 0.573 ** | |||||||
| BT | 0.318 | ||||||||
| PA | 0.708 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosales, R.S.; Risco, D.; García-Nicolás, O.; Pallarés, F.J.; Ramírez, A.S.; Poveda, J.B.; Nicholas, R.A.J.; Salguero, F.J. Differential Gene Expression in Porcine Lung Compartments after Experimental Infection with Mycoplasma hyopneumoniae. Animals 2024, 14, 1290. https://doi.org/10.3390/ani14091290
Rosales RS, Risco D, García-Nicolás O, Pallarés FJ, Ramírez AS, Poveda JB, Nicholas RAJ, Salguero FJ. Differential Gene Expression in Porcine Lung Compartments after Experimental Infection with Mycoplasma hyopneumoniae. Animals. 2024; 14(9):1290. https://doi.org/10.3390/ani14091290
Chicago/Turabian StyleRosales, Rubén S., David Risco, Obdulio García-Nicolás, Francisco J. Pallarés, Ana S. Ramírez, José B. Poveda, Robin A. J. Nicholas, and Francisco J. Salguero. 2024. "Differential Gene Expression in Porcine Lung Compartments after Experimental Infection with Mycoplasma hyopneumoniae" Animals 14, no. 9: 1290. https://doi.org/10.3390/ani14091290
APA StyleRosales, R. S., Risco, D., García-Nicolás, O., Pallarés, F. J., Ramírez, A. S., Poveda, J. B., Nicholas, R. A. J., & Salguero, F. J. (2024). Differential Gene Expression in Porcine Lung Compartments after Experimental Infection with Mycoplasma hyopneumoniae. Animals, 14(9), 1290. https://doi.org/10.3390/ani14091290