

Short- and Long-Term Storage of Non-Domesticated European Mouflon (Ovis aries musimon) Cumulus–Oocyte Complexes Recovered in Field Conditions
 ,
,  , ,
, ,  ,
,  , , , and
, , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Chemicals
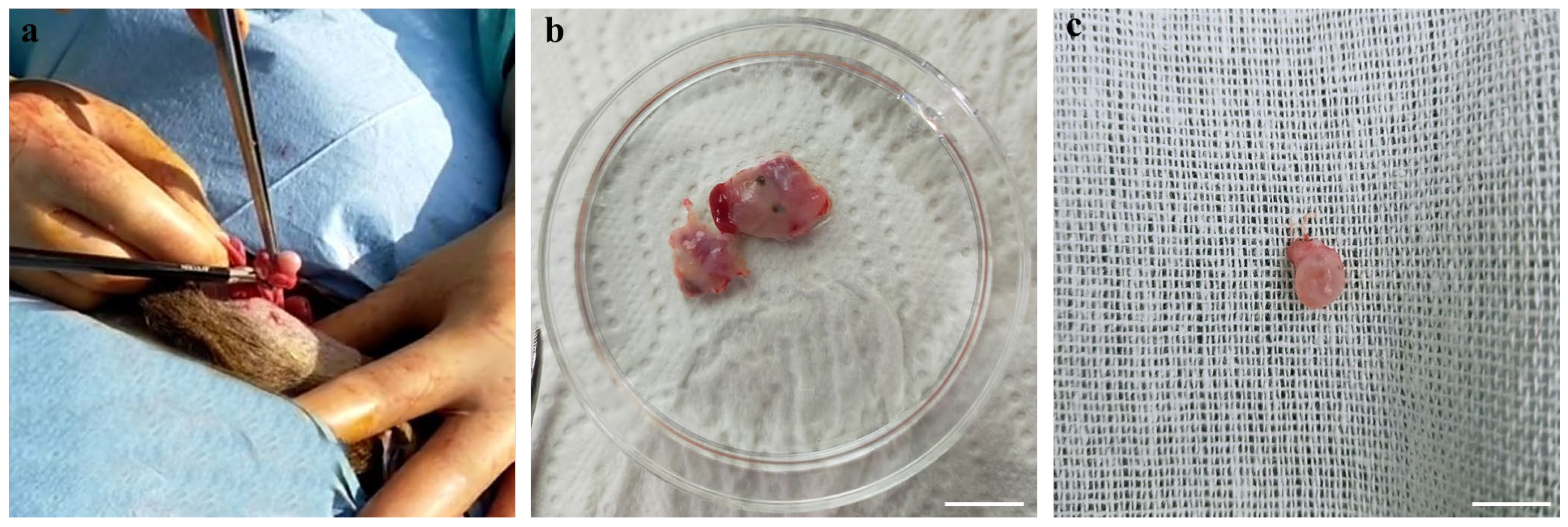
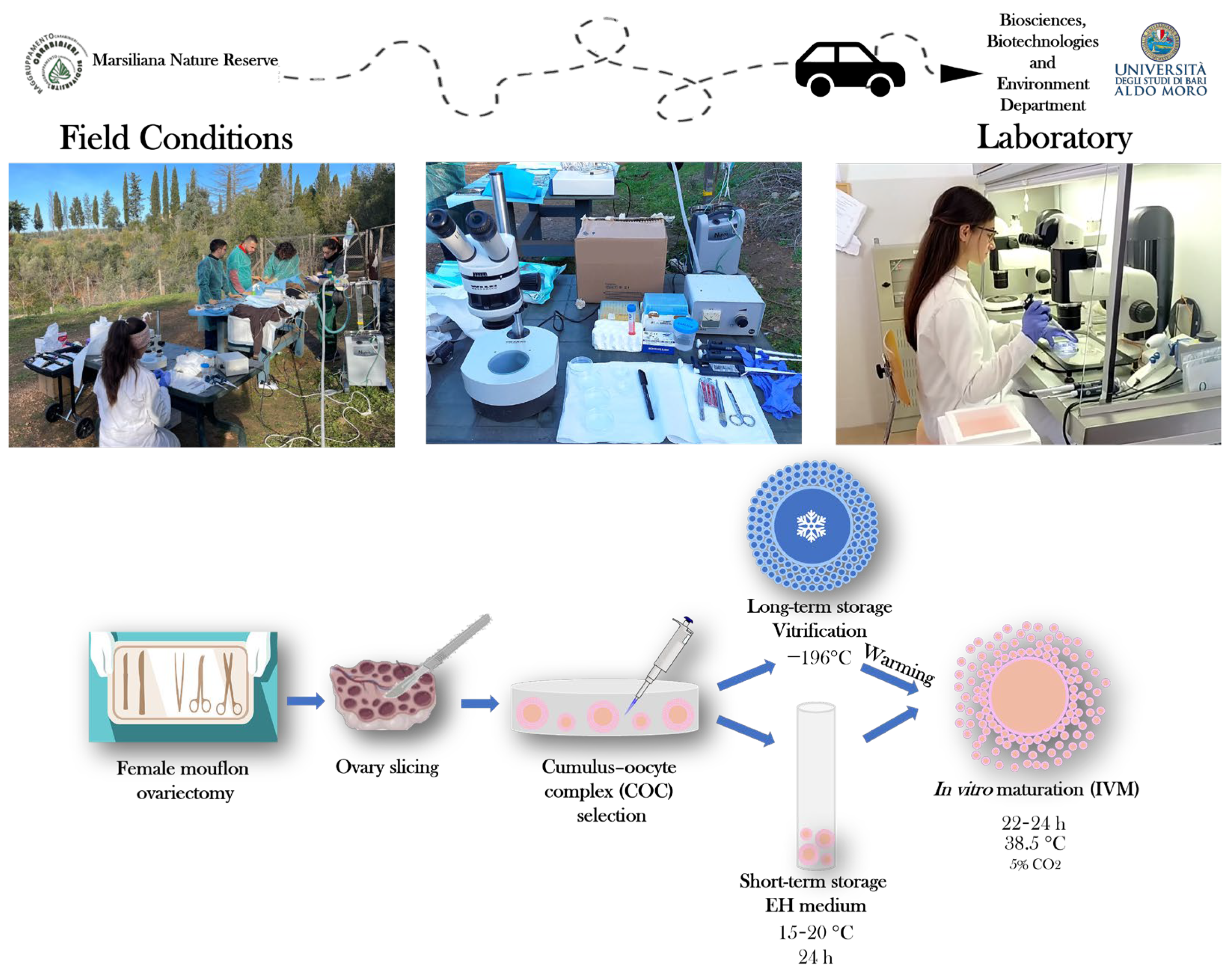
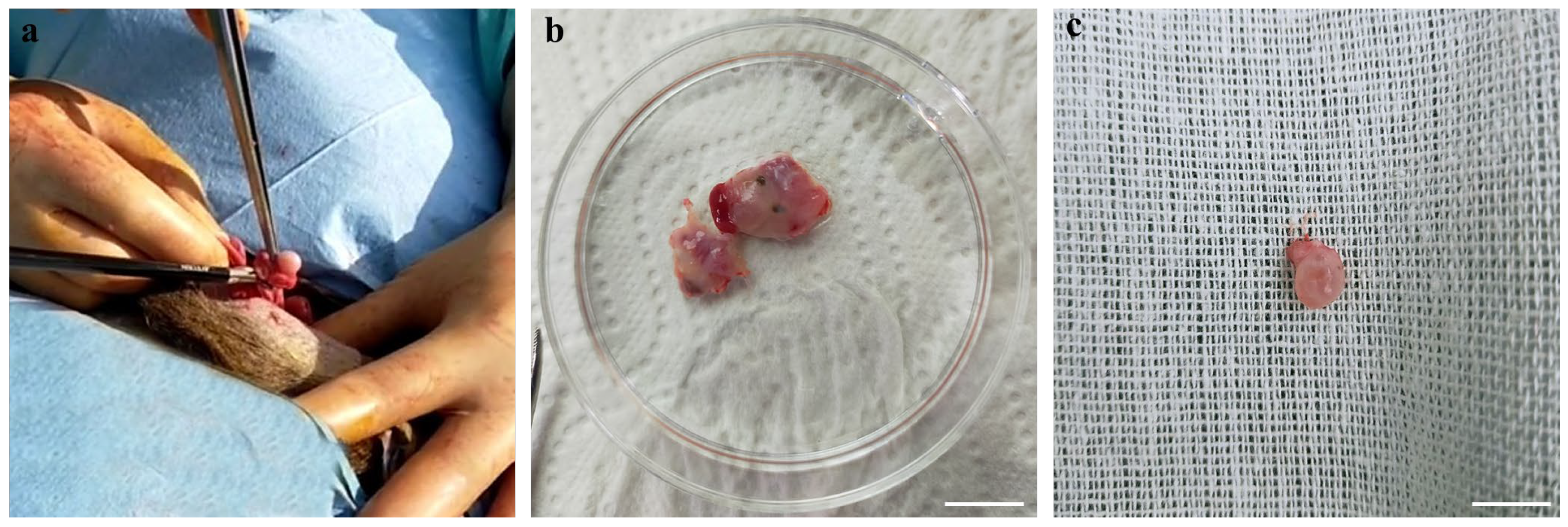
2.3. Ovary Processing and COC Retrieval
2.4. Short-Term Storage of COCs in EH Medium
2.5. Long-Term Storage of COCs by Vitrification
2.6. Warming of Vitrified COCs
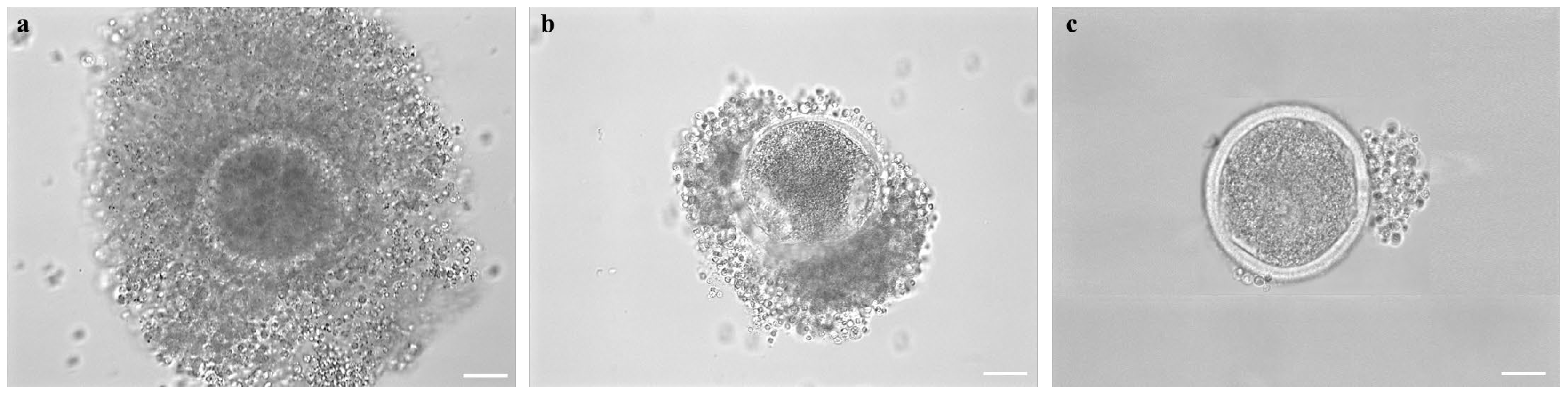
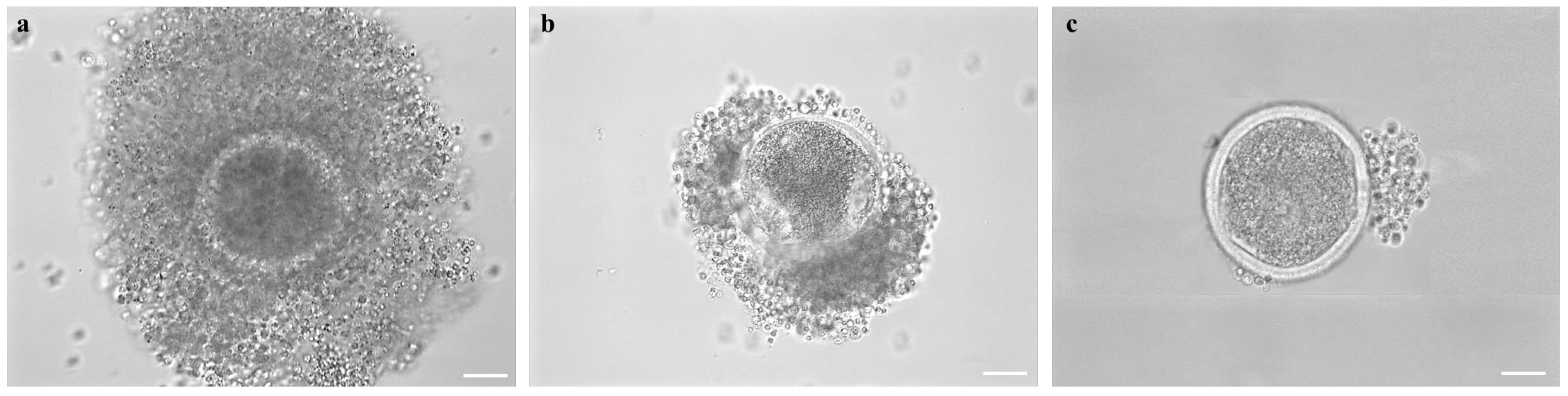
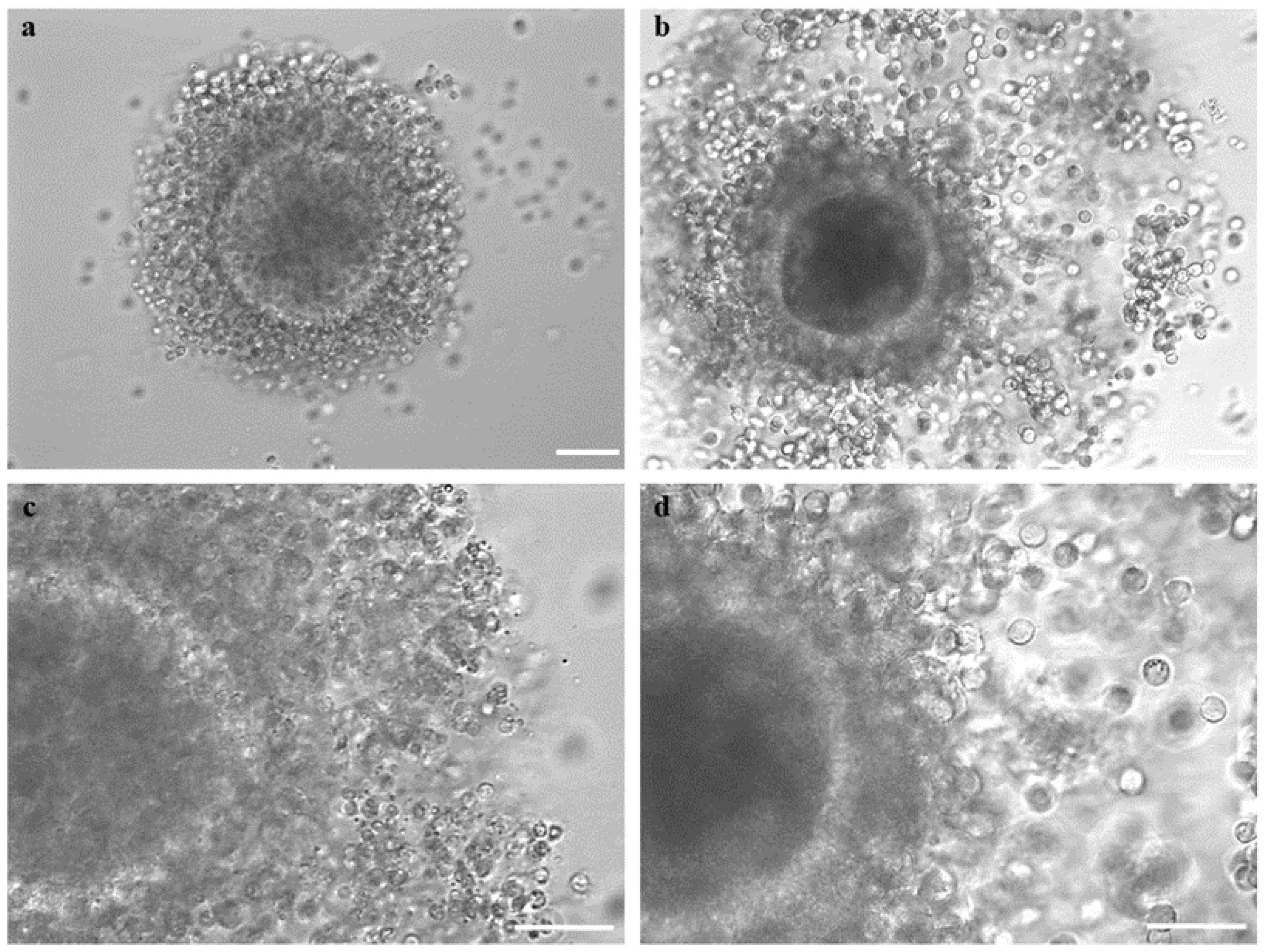
2.7. Assessment of COC Morphology after Short- or Long-Term Storage
2.8. In Vitro Maturation (IVM)
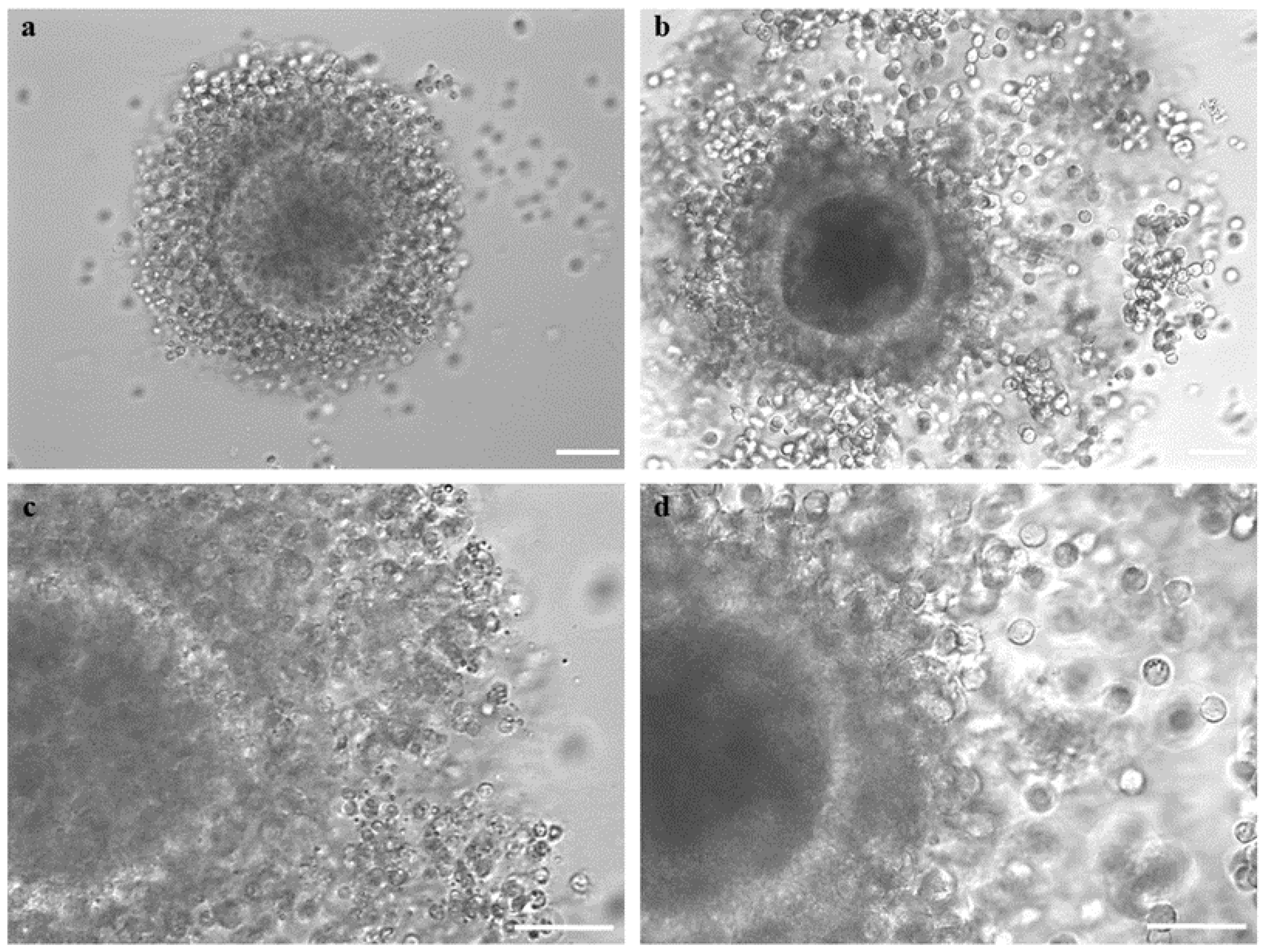
2.9. Assessment of Cumulus Expansion and Oocyte Denuding
2.10. Oocyte Mitochondria and ROS Staining
2.11. Oocyte Nuclear Chromatin Evaluation
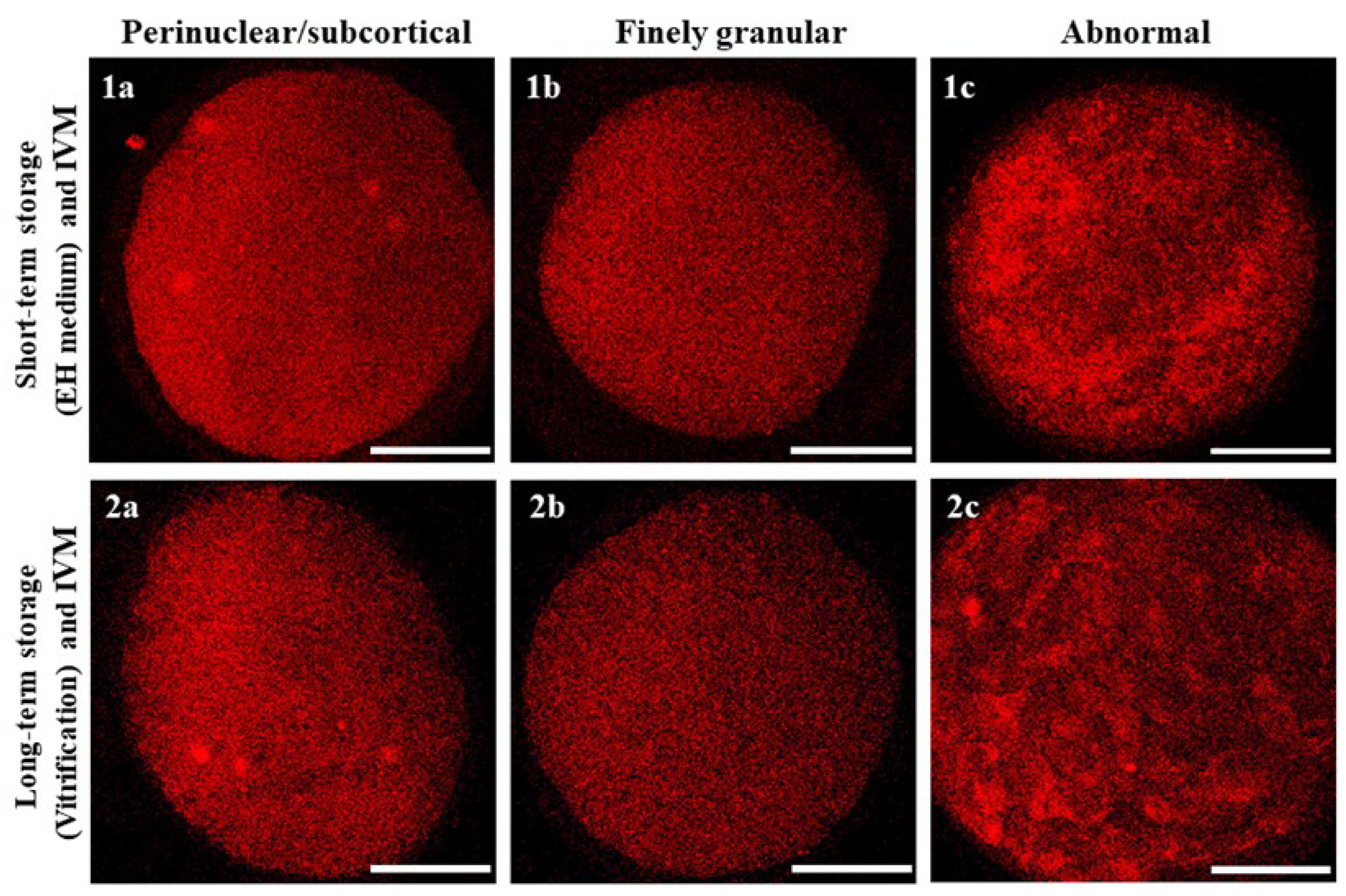
2.12. Assessment of Mitochondrial Distribution Pattern and Intracellular ROS Localization
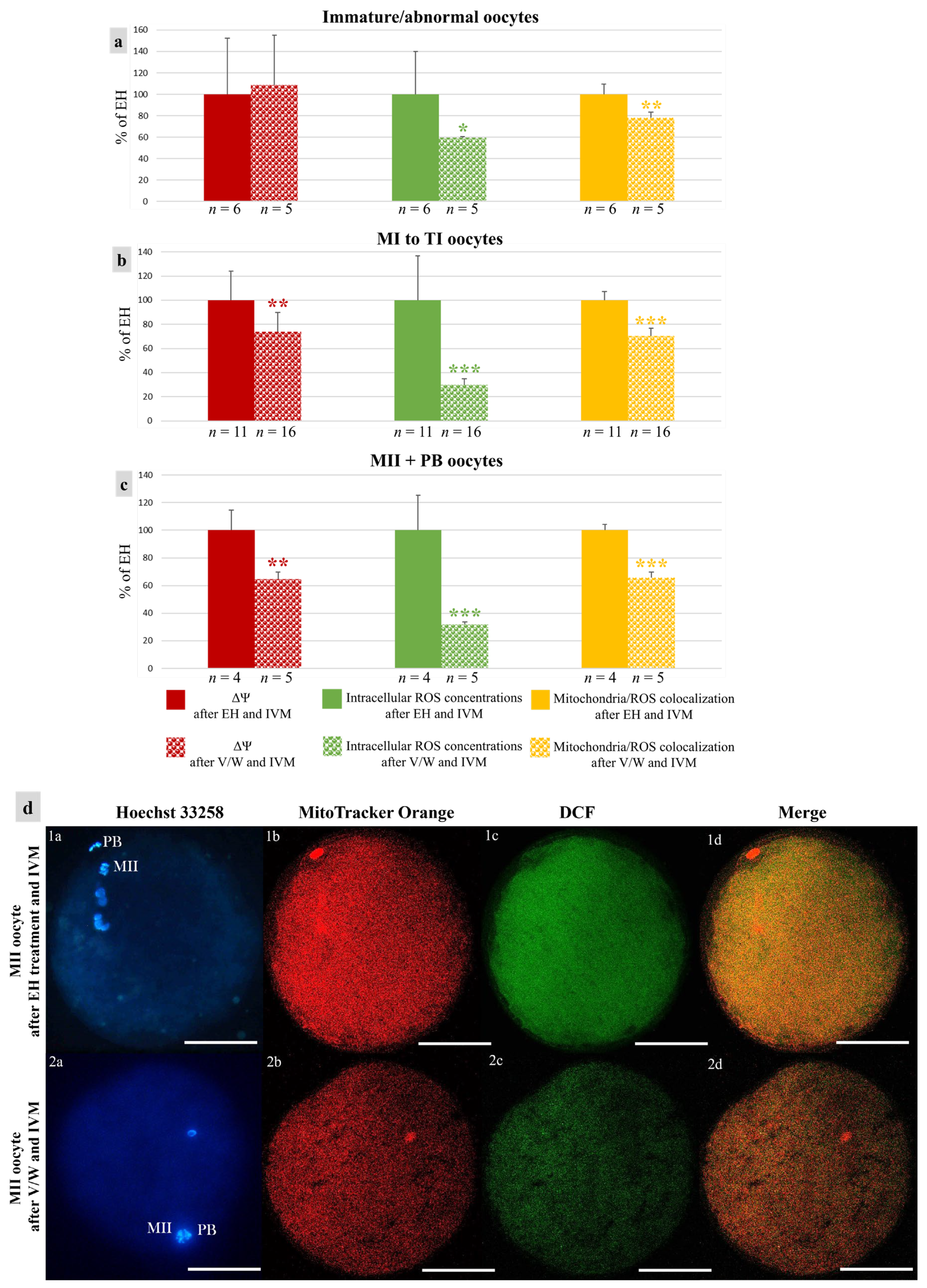
2.13. Quantification of Bioenergetic/Oxidative Variables
2.14. Statistical Analysis
3. Results
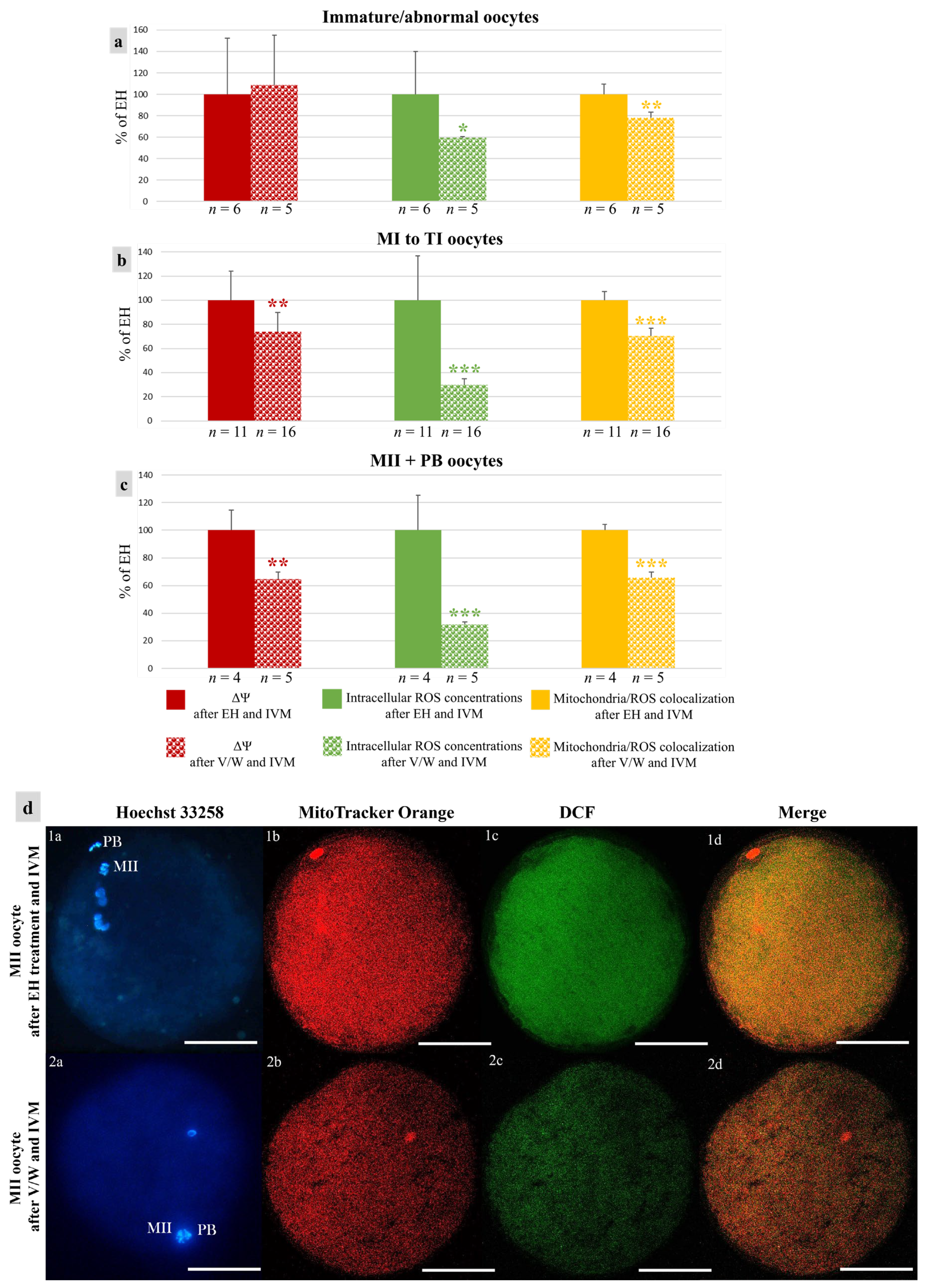
3.1. Short- and Long-Term Storage Preserved Oocyte Meiotic Competence in Mouflons
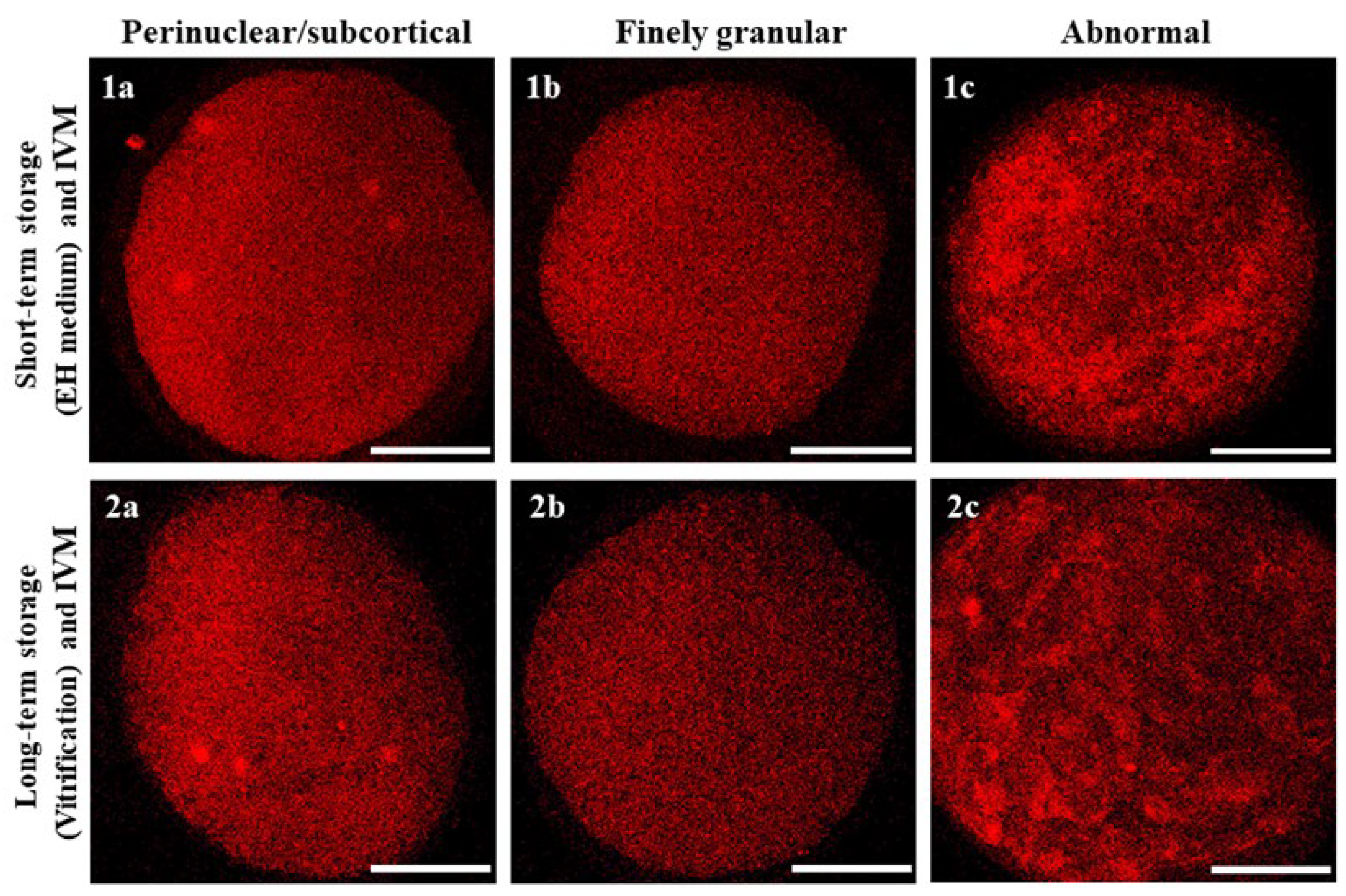
3.2. Short- and Long-Term Storage Did Not Affect Mitochondria Distribution Pattern in Mouflon Oocytes
3.3. Vitrification Affects Quantitative Bioenergetic Parameters in Mouflon Oocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Comizzoli, P.; Songsasen, N.; Wildt, D.E. Protecting and extending fertility for females of wild and endangered mammals. Cancer Treat. Res. 2010, 156, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Bhat, G.R.; Sofi, K.A. Oocyte and embryo preservation in wild animals: An update. Cryo Lett. 2021, 42, 251–260. [Google Scholar]
- de Oliveira Santos, M.V.; Rodrigues Silva, A.; Fernandes Pereira, A. Embryo production by in vitro fertilization in wild ungulates: Progress and perspectives—A review. Ann. Anim. Sci. 2022, 22, 1151–1162. [Google Scholar] [CrossRef]
- Cicirelli, V.; Carbonari, A.; Burgio, M.; Giannini, F.; Rizzo, A. Ovariectomy in Mouflons (Ovis aries) in the Field: Application of Innovative Surgical and Anaesthesiological Techniques. Animals 2023, 13, 491. [Google Scholar] [CrossRef] [PubMed]
- Cerri, D.; Ambrogi, C.; Ebani, V.V.; Poli, A.; Cappelli, F.; Cardini, G.; Andreani, E. Experimental Brucella ovis infection in mouflon (Ovis musimon). J. Wildl. Dis. 2002, 38, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Ptak, G.; Clinton, M.; Barboni, B.; Muzzeddu, M.; Cappai, P.; Tischner, M.; Loi, P. Preservation of the wild European mouflon: The first example of genetic management using a complete program of reproductive biotechnologies. Biol. Reprod. 2002, 66, 796–801. [Google Scholar] [CrossRef] [PubMed]
- Barbato, M.; Masseti, M.; Pirastru, M.; Columbano, N.; Scali, M.; Vignani, R.; Mereu, P. Islands as Time Capsules for Genetic Diversity Conservation: The Case of the Giglio Island Mouflon. Diversity 2022, 14, 609. [Google Scholar] [CrossRef]
- Choi, Y.H.; Love, L.B.; Varner, D.D.; Hinrichs, K. Holding immature equine oocytes in the absence of meiotic inhibitors: Effect on germinal vesicle chromatin and blastocyst development after intracytoplasmic sperm injection. Theriogenology 2006, 66, 955–963. [Google Scholar] [CrossRef]
- Barberino, R.S.; Silva, J.R.V.; Figueiredo, J.R.; Matos, M.H.T. Transport of Domestic and Wild Animal Ovaries: A Review of the Effects of Medium, Temperature, and Periods of Storage on Follicular Viability. Biopreserv. Biobank. 2019, 17, 84–90. [Google Scholar] [CrossRef]
- Hinrichs, K. Advances in Holding and Cryopreservation of Equine Oocytes and Embryos. J. Equine Vet. Sci. 2020, 89, 102990. [Google Scholar] [CrossRef]
- Martino, N.A.; Dell’Aquila, M.E.; Filioli Uranio, M.; Rutigliano, L.; Nicassio, M.; Lacalandra, G.M.; Hinrichs, K. Effect of holding equine oocytes in meiosis inhibitor-free medium before in vitro maturation and of holding temperature on meiotic suppression and mitochondrial energy/redox potential. Reprod. Biol. Endocrinol. 2014, 12, 99. [Google Scholar] [CrossRef] [PubMed]
- Martino, N.A.; Marzano, G.; Mastrorocco, A.; Lacalandra, G.M.; Vincenti, L.; Hinrichs, K.; Dell Aquila, M.E. Use of time-lapse imaging to evaluate morphokinetics of in vitro equine blastocyst development after oocyte holding for two days at 15 °C versus room temperature before intracytoplasmic sperm injection. Reprod. Fertil. Dev. 2019, 31, 1862–1873. [Google Scholar] [CrossRef] [PubMed]
- Alm, H.; Choi, Y.H.; Love, L.; Heleil, B.; Torner, H.; Hinrichs, K. Holding bovine oocytes in the absence of maturation inhibitors: Kinetics of in vitro maturation and effect on blastocyst development after in vitro fertilization. Theriogenology 2008, 70, 1024–1029. [Google Scholar] [CrossRef] [PubMed]
- Pascottini, O.B.; Catteeuw, M.; Van Soom, A.; Opsomer, G. Holding immature bovine oocytes in a commercial embryo holding medium: High developmental competence for up to 10 h at room temperature. Theriogenology 2018, 107, 63–69. [Google Scholar] [CrossRef]
- Yang, C.R.; Miao, D.Q.; Zhang, Q.H.; Guo, L.; Tong, J.S.; Wei, Y.; Huang, X.; Hou, Y.; Schatten, H.; Liu, Z.; et al. Short-term preservation of porcine oocytes in ambient temperature: Novel approaches. PLoS ONE 2010, 5, e14242. [Google Scholar] [CrossRef] [PubMed]
- Macías-García, B.; González-Fernández, L.; Matilla, E.; Hernández, N.; Mijares, J.; Sánchez-Margallo, F.M. Oocyte holding in the Iberian red deer (Cervus elaphus hispanicus): Effect of initial oocyte quality and epidermal growth factor addition on in vitro maturation. Reprod. Domest. Anim. 2018, 53, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Prentice, J.R.; Anzar, M. Cryopreservation of Mammalian oocyte for conservation of animal genetics. Vet. Med. Int. 2010, 2011, 146405. [Google Scholar] [CrossRef]
- Temerario, L.; Monaco, D.; Mastrorocco, A.; Martino, N.A.; Cseh, S.; Lacalandra, G.M.; Ciani, E.; Dell'Aquila, M.E. New Strategies for Conservation of Gentile di Puglia Sheep Breed, an Autochthonous Capital of Millennial Tradition in Southern Italy. Animals 2023, 13, 2371. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.C.; Carrageta, D.F.; Bernardino, R.L.; Alves, M.G.; Oliveira, P.F. Aquaporins and Animal Gamete Cryopreservation: Advances and Future Challenges. Animals 2022, 12, 359. [Google Scholar] [CrossRef]
- Vieira, A.D.; Forell, F.; Feltrin, C.; Rodrigues, J.L. Calves born after direct transfer of vitrified bovine in vitro-produced blastocysts derived from vitrified immature oocytes. Reprod. Domest. Anim. 2008, 43, 314–318. [Google Scholar] [CrossRef]
- Somfai, T.; Yoshioka, K.; Tanihara, F.; Kaneko, H.; Noguchi, J.; Kashiwazaki, N.; Nagai, T.; Kikuchi, K. Generation of live piglets from cryopreserved oocytes for the first time using a defined system for in vitro embryo production. PLoS ONE 2014, 9, e97731. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Escribano, N.; Bogado Pascottini, O.; Woelders, H.; Vandenberghe, L.; De Schauwer, C.; Govaere, J.; Van den Abbeel, E.; Vullers, T.; Ververs, C.; Roles, K.; et al. An improved vitrification protocol for equine immature oocytes, resulting in a first live foal. Equine Vet. J. 2018, 50, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Clérico, G.; Taminelli, G.; Veronesi, J.C.; Polola, J.; Pagura, N.; Pinto, C.; Sansinena, M. Mitochondrial function, blastocyst development and live foals born after ICSI of immature vitrified/warmed equine oocytes matured with or without melatonin. Theriogenology 2021, 160, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Tharasanit, T.; Manee-In, S.; Buarpung, S.; Chatdarong, K.; Lohachit, C.; Techakumphu, M. Successful pregnancy following transfer of feline embryos derived from vitrified immature cat oocytes using ‘stepwise’ cryoprotectant exposure technique. Theriogenology 2011, 76, 1442–1449. [Google Scholar] [CrossRef] [PubMed]
- Rao, B.S.; Mahesh, Y.U.; Suman, K.; Charan, K.V.; Lakshmikantan, U.; Gibence, H.R.; Shivaji, S. Meiotic maturation of vitrified immature chousingha (Tetracerus quadricornis) oocytes recovered postmortem. Cryobiology 2011, 62, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Zahmel, J.; Jänsch, S.; Jewgenow, K.; Sandgreen, D.M.; Skalborg Simonsen, K.; Colombo, M. Maturation and fertilization of African lion (Panthera leo) oocytes after vitrification. Cryobiology 2021, 98, 146–151. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Santos, M.V.; de Queiroz Neta, L.B.; Avezedo Borges, A.; Fernandes Pereira, A. Influence of commercially available follicle stimulating hormone on the in vitro maturation of bovine oocytes. Semin. Ciênc. Agrár. 2017, 38, 1393–1402. [Google Scholar] [CrossRef]
- Yang, H.W.; Hwang, K.J.; Kwon, H.C.; Kim, H.S.; Choi, K.W.; Oh, K.S. Detection of reactive oxygen species (ROS) and apoptosis in human fragmented embryos. Hum. Reprod. 1998, 13, 998–1002. [Google Scholar] [CrossRef]
- Manders, E.M.M.; Verbeek, F.J.; Aten, J.A. Measurement of co-localization of objects in dual-colour confocal images. J. Microsc. 1993, 169, 375–382. [Google Scholar] [CrossRef]
- Bogliolo, L.; Ariu, F.; Fois, S.; Rosati, I.; Zedda, M.T.; Leoni, G.; Succu, S.; Pau, S.; Ledda, S. Morphological and biochemical analysis of immature ovine oocytes vitrified with or without cumulus cells. Theriogenology 2007, 68, 1138–1149. [Google Scholar] [CrossRef]
- Moawad, A.R.; Fisher, P.; Zhu, J.; Choi, I.; Polgar, Z.; Dinnyes, A.; Campbell, K.H. In vitro fertilization of ovine oocytes vitrified by solid surface vitrification at germinal vesicle stage. Cryobiology 2012, 65, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Quan, G.B.; Wu, G.Q.; Wang, Y.J.; Ma, Y.; Lv, C.R.; Hong, Q.H. Meiotic maturation and developmental capability of ovine oocytes at germinal vesicle stage following vitrification using different cryodevices. Cryobiology 2016, 72, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, E.; Shirazi, A.; Shams-Esfandabadi, N.; Nazari, H. Antioxidants and glycine can improve the developmental competence of vitrified/warmed ovine immature oocytes. Reprod. Domest. Anim. 2019, 54, 595–603. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COC Treatment | N. of Healthy Selected COCs (N. of Donors) | COC Morphology after Short- or Long-Term Storage N. (%) | ||
|---|---|---|---|---|
| Preserved | Partially Removed Cumulus | Completely Removed Cumulus | ||
| EH | 23 (3) | 15 (65.2) | 4 (17.4) | 4 (17.4) |
| Vitrification | 26 (8) | 21 (80.8) | 4 (15.4) | 1 (3.8) |
| COC Treatment | N. of Cultured COCs (N. of Donors) | Cumulus Expansion after Short- or Long-Term Storage and IVM N. (%) | ||
|---|---|---|---|---|
| Yes | No | Removed | ||
| EH | 23 (3) | 2 (8.7) | 12 (52.2) | 9 (39.1) |
| Vitrification | 26 (8) | 9 (34.6) | 14 (53.8) | 3 (11.5) |
| COC Treatment | N. of Cultured COCs (N. of Donors) | N. of Evaluated Oocytes | Nuclear Chromatin Characteristics N. (%) | ||
|---|---|---|---|---|---|
| Immature/ Abnormal | MI to TI | MII + PB | |||
| EH | 23 (3) | 21 | 6 (28.6) | 11 (52.4) | 4 (19.0) |
| Vitrification | 26 (8) | 26 | 5 (19.2) | 16 (61.5) | 5 (19.2) |
| COC Treatment | Meiotic Stage | N. of Evaluated Oocytes | Mitochondrial Distribution Pattern N. (%) | ||
|---|---|---|---|---|---|
| P/S | Finely Granular | Abnormal | |||
| EH | Immature/ Abnormal | 6 | 0 (0) | 5 (83.3) | 1 (16.7) |
| Vitrification | Immature/ Abnormal | 5 | 0 (0) | 2 (40.0) | 3 (60.0) |
| EH | MI-TI | 11 | 1 (10.0) | 10 (90.0) | 0 (0) |
| Vitrification | MI-TI | 16 | 2 (12.5) | 13 (81.3) | 1 (6.3) |
| EH | MII + PB | 4 | 1 (25.0) | 3 (75.0) | 0 (0) |
| Vitrification | MII + PB | 5 | 1 (20.0) | 3 (60.0) | 1 (20.0) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Temerario, L.; Cicirelli, V.; Martino, N.A.; Carbonari, A.; Burgio, M.; Frattina, L.; Lacalandra, G.M.; Rizzo, A.; Dell’Aquila, M.E. Short- and Long-Term Storage of Non-Domesticated European Mouflon (Ovis aries musimon) Cumulus–Oocyte Complexes Recovered in Field Conditions. Animals 2024, 14, 807. https://doi.org/10.3390/ani14050807
Temerario L, Cicirelli V, Martino NA, Carbonari A, Burgio M, Frattina L, Lacalandra GM, Rizzo A, Dell’Aquila ME. Short- and Long-Term Storage of Non-Domesticated European Mouflon (Ovis aries musimon) Cumulus–Oocyte Complexes Recovered in Field Conditions. Animals. 2024; 14(5):807. https://doi.org/10.3390/ani14050807
Chicago/Turabian StyleTemerario, Letizia, Vincenzo Cicirelli, Nicola Antonio Martino, Alice Carbonari, Matteo Burgio, Lorenza Frattina, Giovanni Michele Lacalandra, Annalisa Rizzo, and Maria Elena Dell’Aquila. 2024. "Short- and Long-Term Storage of Non-Domesticated European Mouflon (Ovis aries musimon) Cumulus–Oocyte Complexes Recovered in Field Conditions" Animals 14, no. 5: 807. https://doi.org/10.3390/ani14050807
APA StyleTemerario, L., Cicirelli, V., Martino, N. A., Carbonari, A., Burgio, M., Frattina, L., Lacalandra, G. M., Rizzo, A., & Dell’Aquila, M. E. (2024). Short- and Long-Term Storage of Non-Domesticated European Mouflon (Ovis aries musimon) Cumulus–Oocyte Complexes Recovered in Field Conditions. Animals, 14(5), 807. https://doi.org/10.3390/ani14050807