
Characterizing the Effect of Campylobacter jejuni Challenge on Growth Performance, Cecal Microbiota, and Cecal Short-Chain Fatty Acid Concentrations in Broilers
 , ,
, ,  , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Birds and Experimental Setup
2.2. C. jejuni Challenge Preparation
2.3. Cecal Sample Collection and DNA Extraction
2.4. DNA Sequencing and Bioinformatics Analysis
2.5. Short-Chain Fatty Acid Analysis
2.6. Statistical Analysis
3. Results and Discussion
3.1. Performance Parameters
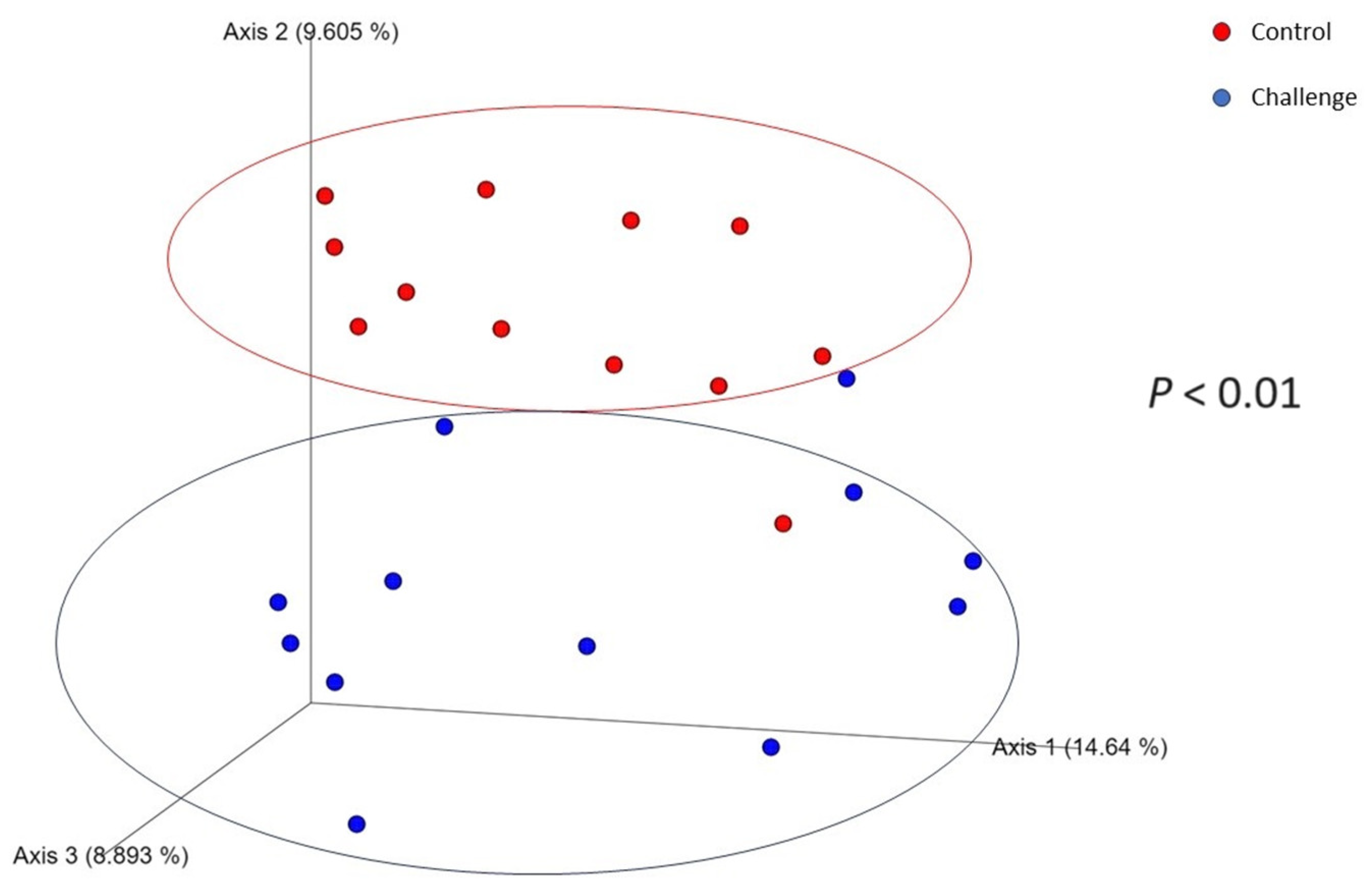
3.2. Microbial Diversity
3.3. Bacterial Diversity at the Phylum Level
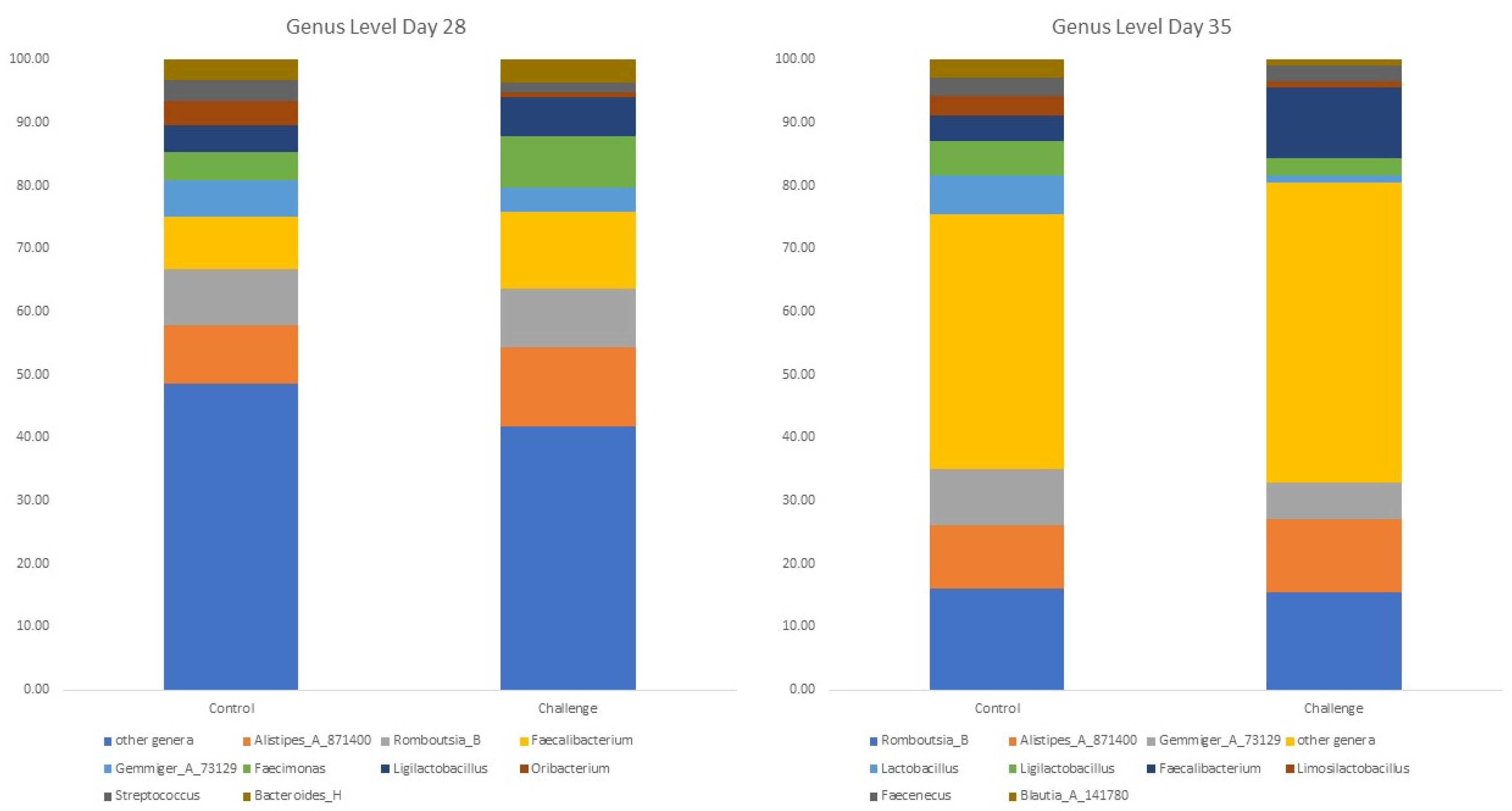
3.4. Bacterial Diversity at the Genera and Species Level
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global epidemiology of Campylobacter infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [PubMed]
- Ben Romdhane, R.; Merle, R. The data behind risk analysis of Campylobacter jejuni and Campylobacter coli infections. In Fighting Campylobacter Infections: Towards a One Health Approach; Springer: Berlin/Heidelberg, Germany, 2021; pp. 25–58. [Google Scholar]
- Hoffmann, S.A.; Maculloch, B.; Batz, M. Economic Burden of Major Foodborne Illnesses Acquired in the United States; United States Department of Agriculture: Washington, DC, USA, 2015. [Google Scholar]
- Oh, E.; McMullen, L.; Jeon, B. High prevalence of hyper-aerotolerant Campylobacter jejuni in retail poultry with potential implication in human infection. Front. Microbiol. 2015, 6, 1263. [Google Scholar] [CrossRef] [PubMed]
- Olvera-Ramírez, A.M.; McEwan, N.R.; Stanley, K.; Nava-Diaz, R.; Aguilar-Tipacamú, G. A Systematic Review on the Role of Wildlife as Carriers and Spreaders of Campylobacter spp. Animals 2023, 13, 1334. [Google Scholar] [CrossRef] [PubMed]
- Cawthraw, S.; Newell, D. Investigation of the presence and protective effects of maternal antibodies against Campylobacter jejuni in chickens. Avian Dis. 2010, 54, 86–93. [Google Scholar] [CrossRef]
- Skarp, C.; Hänninen, M.-L.; Rautelin, H. Campylobacteriosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 103–109. [Google Scholar] [CrossRef]
- Pielsticker, C.; Glünder, G.; Rautenschlein, S. Colonization properties of Campylobacter jejuni in chickens. Eur. J. Microbiol. Immunol. 2012, 2, 61–65. [Google Scholar] [CrossRef]
- Gruntar, I.; Biasizzo, M.; Kušar, D.; Pate, M.; Ocepek, M. Campylobacter jejuni contamination of broiler carcasses: Population dynamics and genetic profiles at slaughterhouse level. Food Microbiol. 2015, 50, 97–101. [Google Scholar] [CrossRef]
- Laban, S.E.; Khalefa, H.S. Impact of Microbial Load of Slaughterhouse Environment on the Degree of Broiler Chicken Carcass Contamination, with a Focus on Campylobacter Prevalence. J. Adv. Vet. Res. 2023, 13, 1551–1559. [Google Scholar]
- Bolton, D.J. Campylobacter virulence and survival factors. Food Microbiol. 2015, 48, 99–108. [Google Scholar] [CrossRef]
- Ortega-Sanz, I.; García, M.; Bocigas, C.; Megías, G.; Melero, B.; Rovira, J. Genomic characterization of Campylobacter jejuni associated with perimyocarditis: A family case report. Foodborne Pathog. Dis. 2023, 20, 368–373. [Google Scholar] [CrossRef]
- Mortada, M.; Cosby, D.E.; Akerele, G.; Ramadan, N.; Oxford, J.; Shanmugasundaram, R.; Ng, T.T.; Selvaraj, R.K. Characterizing the immune response of chickens to Campylobacter jejuni (Strain A74C). PLoS ONE 2021, 16, e0247080. [Google Scholar] [CrossRef]
- Looft, T.; Cai, G.; Choudhury, B.; Lai, L.X.; Lippolis, J.D.; Reinhardt, T.A.; Sylte, M.J.; Casey, T.A. Avian Intestinal Mucus Modulates Campylobacter jejuni Gene Expression in a Host-Specific Manner. Front. Microbiol. 2018, 9, 3215. [Google Scholar] [CrossRef]
- Alemka, A.; Corcionivoschi, N.; Bourke, B. Defense and adaptation: The complex inter-relationship between Campylobacter jejuni and mucus. Front. Cell. Infect. Microbiol. 2012, 2, 15. [Google Scholar] [CrossRef]
- Duangnumsawang, Y.; Zentek, J.; Goodarzi Boroojeni, F. Development and functional properties of intestinal mucus layer in poultry. Front. Immunol. 2021, 12, 745849. [Google Scholar] [CrossRef]
- Struwe, W.B.; Gough, R.; Gallagher, M.E.; Kenny, D.T.; Carrington, S.D.; Karlsson, N.G.; Rudd, P.M. Identification of O-glycan Structures from Chicken Intestinal Mucins Provides Insight into Campylobactor jejuni Pathogenicity*[S]. Mol. Cell. Proteom. 2015, 14, 1464–1477. [Google Scholar] [CrossRef]
- Munoz, L.R.; Bailey, M.A.; Krehling, J.T.; Bourassa, D.V.; Hauck, R.; Pacheco, W.J.; Chaves-Cordoba, B.; Chasteen, K.S.; Talorico, A.A.; Escobar, C.; et al. Effects of dietary yeast cell wall supplementation on growth performance, intestinal Campylobacter jejuni colonization, innate immune response, villus height, crypt depth, and slaughter characteristics of broiler chickens inoculated with Campylobacter jejuni at d 21. Poult. Sci. 2023, 102, 102609. [Google Scholar] [CrossRef]
- Humphrey, S.; Chaloner, G.; Kemmett, K.; Davidson, N.; Williams, N.; Kipar, A.; Humphrey, T.; Wigley, P. Campylobacter jejuni is not merely a commensal in commercial broiler chickens and affects bird welfare. MBio 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Awad, W.A.; Molnár, A.; Aschenbach, J.R.; Ghareeb, K.; Khayal, B.; Hess, C.; Liebhart, D.; Dublecz, K.; Hess, M. Campylobacter infection in chickens modulates the intestinal epithelial barrier function. Innate Immun. 2015, 21, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, T.; Meredith, H.; Vigors, S.; McDonnell, M.J.; Ryan, M.; Thornton, K.; O’Doherty, J.V. Extracts of laminarin and laminarin/fucoidan from the marine macroalgal species Laminaria digitata improved growth rate and intestinal structure in young chicks, but does not influence Campylobacter jejuni colonisation. Anim. Feed Sci. Technol. 2017, 232, 71–79. [Google Scholar] [CrossRef]
- Chagneau, S.; Gaucher, M.-L.; Fravalo, P.; Thériault, W.P.; Thibodeau, A. Intestinal Colonization of Campylobacter jejuni and Its Hepatic Dissemination Are Associated with Local and Systemic Immune Responses in Broiler Chickens. Microorganisms 2023, 11, 1677. [Google Scholar] [CrossRef] [PubMed]
- Thibodeau, A.; Fravalo, P.; Yergeau, É.; Arsenault, J.; Lahaye, L.; Letellier, A. Chicken Caecal Microbiome Modifications Induced by Campylobacter jejuni Colonization and by a Non-Antibiotic Feed Additive. PLoS ONE 2015, 10, e0131978. [Google Scholar] [CrossRef] [PubMed]
- Sofka, D.; Pfeifer, A.; Gleiß, B.; Paulsen, P.; Hilbert, F. Changes within the intestinal flora of broilers by colonisation with Campylobacter jejuni. Berl. Münch. Tierärztl. Wochensch. 2015, 128, 104–110. [Google Scholar]
- Skånseng, B.; Kaldhusdal, M.; Rudi, K. Comparison of chicken gut colonisation by the pathogens Campylobacter jejuni and Clostridium perfringens by real-time quantitative PCR. Mol. Cell. Probes 2006, 20, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; Grinevich, D.; Thakur, S.; Balamotis, M.A.; Yehezkel, T.B. Ultra-accurate microbial amplicon sequencing with synthetic long reads. Microbiome 2021, 9, 130. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.B.; Holton, M.P.; Callaway, T.R.; Lourenco, J.M.; Fontes, P.L.P. Differences in Microbial Community Composition between Uterine Horns Ipsilateral and Contralateral to the Corpus Luteum in Beef Cows on Day 15 of the Estrous Cycle. Microorganisms 2023, 11, 2117. [Google Scholar] [CrossRef] [PubMed]
- Cason, E.; Al Hakeem, W.; Adams, D.; Shanmugasundaram, R.; Selvaraj, R. Effects of synbiotic supplementation as an antibiotic growth promoter replacement on cecal Campylobacter jejuni load in broilers challenged with C. jejuni. J. Appl. Poult. Res. 2023, 32, 100315. [Google Scholar] [CrossRef]
- Rothrock, M.J., Jr.; Hiett, K.L.; Gamble, J.; Caudill, A.C.; Cicconi-Hogan, K.M.; Caporaso, J.G. A hybrid DNA extraction method for the qualitative and quantitative assessment of bacterial communities from poultry production samples. JoVE (J. Vis. Exp.) 2014, 94, e52161. [Google Scholar]
- Abellan-Schneyder, I.; Siebert, A.; Hofmann, K.; Wenning, M.; Neuhaus, K. Full-Length SSU rRNA Gene Sequencing Allows Species-Level Detection of Bacteria, Archaea, and Yeasts Present in Milk. Microorganisms 2021, 9, 1251. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High resolution sample inference from amplicon data. BioRxiv 2015, 13, 024034. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V. Scikit-learn: Machine learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundaram, R.; Lourenco, J.; Hakeem, W.A.; Dycus, M.M.; Applegate, T.J. Subclinical doses of dietary fumonisins and deoxynivalenol cause cecal microbiota dysbiosis in broiler chickens challenged with Clostridium perfringens. Front. Microbiol. 2023, 14, 1106604. [Google Scholar] [CrossRef] [PubMed]
- Al Hakeem, W.G.; Fathima, S.; Shanmugasundaram, R.; Selvaraj, R.K. Campylobacter jejuni in poultry: Pathogenesis and control strategies. Microorganisms 2022, 10, 2134. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Jiang, Q.; Tang, H.; Wang, Z.; Yin, Y.; Ren, F.; Kong, L.; Jiao, X.; Huang, J. Characterization and prevalence of Campylobacter spp. from broiler chicken rearing period to the slaughtering process in Eastern China. Front. Vet. Sci. 2020, 7, 227. [Google Scholar] [CrossRef]
- Sahin, O.; Zhang, Q.; Meitzler, J.C.; Harr, B.S.; Morishita, T.Y.; Mohan, R. Prevalence, antigenic specificity, and bactericidal activity of poultry anti-Campylobacter maternal antibodies. Appl. Environ. Microbiol. 2001, 67, 3951–3957. [Google Scholar] [CrossRef]
- Gharib, N.K.; Rahimi, S.; Khaki, P. Comparison of the effects of probiotic, organic acid and medicinal plant on Campylobacter jejuni challenged broiler chickens. J. Agr. Sci. Tech. 2012, 14, 1485–1496. [Google Scholar]
- Al Hakeem, W.G.; Acevedo Villanueva, K.Y.; Selvaraj, R.K. The Development of Gut Microbiota and Its Changes Following C. jejuni Infection in Broilers. Vaccines 2023, 11, 595. [Google Scholar]
- Indikova, I.; Humphrey, T.J.; Hilbert, F. Survival with a helping hand: Campylobacter and microbiota. Front. Microbiol. 2015, 6, 1266. [Google Scholar] [CrossRef]
- Sciuto, K.; Moro, I. Cyanobacteria: The bright and dark sides of a charming group. Biodivers. Conserv. 2015, 24, 711–738. [Google Scholar] [CrossRef]
- Codd, G.A.; Morrison, L.F.; Metcalf, J.S. Cyanobacterial toxins: Risk management for health protection. Toxicol. Appl. Pharmacol. 2005, 203, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-C.; Huang, M.-Y.; Balasubramanian, B.; Jha, R. Heat stress affects jejunal immunity of Yellow-Feathered broilers and is potentially mediated by the microbiome. Front. Physiol. 2022, 13, 1022. [Google Scholar] [CrossRef] [PubMed]
- Hermans, D.; Pasmans, F.; Heyndrickx, M.; Van Immerseel, F.; Martel, A.; Van Deun, K.; Haesebrouck, F. A tolerogenic mucosal immune response leads to persistent Campylobacter jejuni colonization in the chicken gut. Crit. Rev. Microbiol. 2012, 38, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Michaelsen, T.Y.; Singleton, C.M.; Dottorini, G.; Kirkegaard, R.H.; Albertsen, M.; Nielsen, P.H.; Dueholm, M.S. Novel syntrophic bacteria in full-scale anaerobic digesters revealed by genome-centric metatranscriptomics. ISME J. 2020, 14, 906–918. [Google Scholar] [CrossRef] [PubMed]
- Barton, L.L.; Ritz, N.L.; Fauque, G.D.; Lin, H.C. Sulfur cycling and the intestinal microbiome. Dig. Dis. Sci. 2017, 62, 2241–2257. [Google Scholar] [CrossRef] [PubMed]
- Stahl, M.; Friis, L.M.; Nothaft, H.; Liu, X.; Li, J.; Szymanski, C.M.; Stintzi, A. L-fucose utilization provides Campylobacter jejuni with a competitive advantage. Proc. Natl. Acad. Sci. USA 2011, 108, 7194–7199. [Google Scholar] [CrossRef] [PubMed]
- Gilroy, R.; Ravi, A.; Getino, M.; Pursley, I.; Horton, D.L.; Alikhan, N.-F.; Baker, D.; Gharbi, K.; Hall, N.; Watson, M. Extensive microbial diversity within the chicken gut microbiome revealed by metagenomics and culture. PeerJ 2021, 9, e10941. [Google Scholar] [CrossRef]
- Leylabadlo, H.E.; Ghotaslou, R.; Feizabadi, M.M.; Farajnia, S.; Moaddab, S.Y.; Ganbarov, K.; Khodadadi, E.; Tanomand, A.; Sheykhsaran, E.; Yousefi, B. The critical role of Faecalibacterium prausnitzii in human health: An overview. Microb. Pathog. 2020, 149, 104344. [Google Scholar] [CrossRef]
- Auger, S.; Kropp, C.; Borras-Nogues, E.; Chanput, W.; Andre-Leroux, G.; Gitton-Quent, O.; Benevides, L.; Breyner, N.; Azevedo, V.; Langella, P. Intraspecific diversity of microbial anti-inflammatory molecule (MAM) from Faecalibacterium prausnitzii. Int. J. Mol. Sci. 2022, 23, 1705. [Google Scholar] [CrossRef]
- Muñoz, M.; Guerrero-Araya, E.; Cortés-Tapia, C.; Plaza-Garrido, A.; Lawley, T.D.; Paredes-Sabja, D. Comprehensive genome analyses of Sellimonas intestinalis, a potential biomarker of homeostasis gut recovery. Microb. Genom. 2020, 6. [Google Scholar] [CrossRef]
- Luethy, P.M.; Huynh, S.; Ribardo, D.A.; Winter, S.E.; Parker, C.T.; Hendrixson, D.R. Microbiota-Derived Short-Chain Fatty Acids Modulate Expression of Campylobacter jejuni Determinants Required for Commensalism and Virulence. mBio 2017, 8. [Google Scholar] [CrossRef]
- Stahl, M.; Butcher, J.; Stintzi, A. Nutrient acquisition and metabolism by Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2012, 2, 5. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Basal Diet (0–35 d) | % |
|---|---|
| Corn | 58.47 |
| Soybean meal | 35.15 |
| Soybean oil | 2.269 |
| Limestone | 1.593 |
| Biofos | 1.387 |
| NaCl | 0.35 |
| Vitamins premix 1 | 0.35 |
| Dl-methionine | 0.21 |
| Lysine HCL | 0.137 |
| Trace mineral premix 2 | 0.08 |
| Total | 100 |
| Calculated Nutrient Composition | |
| ME, kcal/kg | 3050 |
| Crude protein, % | 21.44 |
| Crude fat, % | 4.55 |
| Lysine, % | 1.31 |
| Calcium, % | 0.95 |
| TSAA, % | 0.91 |
| Threonine, % | 0.87 |
| Methionine, % | 0.56 |
| Available phosphorus, % | 0.45 |
| Body Weight Gain(g) | Control | Challenge | SEM 1 | p-Value |
|---|---|---|---|---|
| 0–21 | 697 | 683 | 12.78 | 0.28 |
| 0–28 | 1271 | 1189 | 33.57 | 0.11 |
| 0–35 | 1848 | 1727 | 54.69 | 0.14 |
| 21–28 | 574 | 517 | 24.79 | 0.13 |
| 21–35 | 1151 | 1053 | 46.10 | 0.16 |
| 28–35 | 577 | 537 | 34.15 | 0.42 |
| Feed Intake (g) | Control | Challenge | SEM 1 | p-Value |
| 0–21 | 1331 | 1267 | 57.31 | 0.44 |
| 0–28 | 2244 | 2075 | 112.53 | 0.31 |
| 0–35 | 3257 | 3053 | 132.19 | 0.30 |
| 21–28 | 913 | 808 | 74.19 | 0.34 |
| 21–35 | 1925 | 1786 | 92.37 | 0.31 |
| 28–35 | 1012 | 977 | 39.20 | 0.54 |
| Feed Conversion Ratio | Control | Challenge | SEM 1 | p-Value |
| 0–21 | 1.90 | 1.88 | 0.09 | 0.89 |
| 0–28 | 1.76 | 1.75 | 0.09 | 0.92 |
| 0–35 | 1.76 | 1.77 | 0.08 | 0.90 |
| 21–28 | 1.6 | 1.58 | 0.14 | 0.93 |
| 21–35 | 1.68 | 1.70 | 0.10 | 0.88 |
| 28–35 | 1.77 | 1.84 | 0.10 | 0.64 |
| Alpha Index | Day 28 | Day 35 | ||||
|---|---|---|---|---|---|---|
| Control | Challenge | p-Value | Control | Challenge | p-Value | |
| Observed features | 183 | 141 | 0.03 | 166 | 148 | 0.50 |
| Faith Index | 9.1 | 8.7 | 0.87 | 10.94 | 13.24 | 0.42 |
| Evenness index | 0.84 | 0.81 | 0.14 | 0.83 | 0.81 | 0.14 |
| Shannon index | 6.31 | 5.79 | 0.05 | 6.11 | 5.82 | 0.26 |
| Phylum (%) | Day 28 | Phylum (%) | Day 35 | ||||
|---|---|---|---|---|---|---|---|
| Control | Challenge | p-Value | Control | Challenge | p-Value | ||
| Firmicutes | 86.08 | 80.68 | 0.17 | Firmicutes | 86.15 | 78.42 | 0.78 |
| Bacteroidota | 13.43 | 16.66 | 0.69 | Bacteroidota | 12.20 | 14.83 | 0.93 |
| Cyanobacteria | 0.21 b | 1.73 a | 0.01 | Cyanobacteria | 0.26 | 0.95 | 0.78 |
| Patescibacteria | 0.04 | 0.03 | 0.73 | Patescibacteria | 0.16 | 2.99 | 0.19 |
| Desulfobacterota | 0.00 | 0.05 | 0.40 | Desulfobacterota | 0.00 | 0.54 | 0.02 |
| Proteobacteria | 0.21 | 0.54 | 0.92 | Proteobacteria | 0.00 | 0.36 | 0.66 |
| Actinobacteriota | 0.04 | 0.03 | 0.78 | Actinobacteriota | 0.05 | 0.07 | 0.17 |
| Other phyla | 0.00 | 0.27 | 0.17 | Other phyla | 1.17 | 1.85 | 0.67 |
| Genera | Day 28 | Genera | Day 35 | ||||
|---|---|---|---|---|---|---|---|
| Control | Challenge | p-Value | Control | Challenge | p-Value | ||
| Acutalibacter | 1.54 a | 0.32 b | 0.004 | Acutalibacter | 1.08 a | 0.34 b | 0.01 |
| UBA644 | 0.09 a | 0.00 b | 0.009 | Oribacterium | 2.45 a | 1.63 b | 0.01 |
| Stercorousia | 0.21 b | 1.73 a | 0.01 | Mailhella | 0.00 | 0.54 | 0.02 |
| UBA1417 | 0.49 | 0.02 | 0.02 | RUG13038 | 0.05 | 0.42 | 0.04 |
| Sellimonas | 0.89 | 0.24 | 0.02 | Faecalibacterium | 3.99 | 11.21 | 0.04 |
| Oribacterium | 3.73 | 0.80 | 0.04 | ||||
| Species | Day 28 | Species | Day 35 | ||||
|---|---|---|---|---|---|---|---|
| Control | Challenge | p-Value | Control | Challenge | p-Value | ||
| UBA644 sp002299265 | 0.09 a | 0 b | 0.009 | Onthocola_B sp000437355 | 0.15 b | 0.61 a | 0.01 |
| Sellimonas intestinalis | 0.89 | 0.24 | 0.02 | Mailhella massiliensis | 0.00 | 0.54 | 0.02 |
| Evtepia gabavorous | 0.31 | 0.10 | 0.03 | Faecalibacterium sp002160895 | 3.99 | 11.17 | 0.04 |
| Day 28 | Day 35 | |||||||
|---|---|---|---|---|---|---|---|---|
| SCFAs 1 | Control | Challenge | SEM 2 | p-Value | Control | Challenge | SEM 2 | p-Value |
| Acetate (mM) | 27.58 | 29.22 | 2.08 | 0.59 | 30.38 | 33.69 | 3.14 | 0.47 |
| Propionate (mM) | 1.18 | 1.15 | 0.27 | 0.94 | 1.20 | 1.13 | 0.16 | 0.76 |
| Butyrate (mM) | 3.35 | 2.27 | 0.66 | 0.28 | 4.70 | 5.67 | 0.78 | 0.40 |
| Isovalerate (mM) | 0.02 | 0.07 | 0.05 | 0.44 | 0.13 | 0.07 | 0.04 | 0.37 |
| Valerate (mM) | 0.14 | 0.17 | 0.05 | 0.70 | 0.28 | 0.33 | 0.05 | 0.51 |
| Total VFA (mM) | 32.30 | 32.91 | 2.69 | 0.87 | 36.72 | 40.91 | 3.97 | 0.47 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Hakeem, W.G.; Cason, E.E.; Adams, D.; Fathima, S.; Shanmugasundaram, R.; Lourenco, J.; Selvaraj, R.K. Characterizing the Effect of Campylobacter jejuni Challenge on Growth Performance, Cecal Microbiota, and Cecal Short-Chain Fatty Acid Concentrations in Broilers. Animals 2024, 14, 473. https://doi.org/10.3390/ani14030473
Al Hakeem WG, Cason EE, Adams D, Fathima S, Shanmugasundaram R, Lourenco J, Selvaraj RK. Characterizing the Effect of Campylobacter jejuni Challenge on Growth Performance, Cecal Microbiota, and Cecal Short-Chain Fatty Acid Concentrations in Broilers. Animals. 2024; 14(3):473. https://doi.org/10.3390/ani14030473
Chicago/Turabian StyleAl Hakeem, Walid G., Emily E. Cason, Daniel Adams, Shahna Fathima, Revathi Shanmugasundaram, Jeferson Lourenco, and Ramesh K. Selvaraj. 2024. "Characterizing the Effect of Campylobacter jejuni Challenge on Growth Performance, Cecal Microbiota, and Cecal Short-Chain Fatty Acid Concentrations in Broilers" Animals 14, no. 3: 473. https://doi.org/10.3390/ani14030473
APA StyleAl Hakeem, W. G., Cason, E. E., Adams, D., Fathima, S., Shanmugasundaram, R., Lourenco, J., & Selvaraj, R. K. (2024). Characterizing the Effect of Campylobacter jejuni Challenge on Growth Performance, Cecal Microbiota, and Cecal Short-Chain Fatty Acid Concentrations in Broilers. Animals, 14(3), 473. https://doi.org/10.3390/ani14030473