


A Special Relationship—Aspects of Human–Animal Interaction in Birds of Prey, Brown Bears, Beavers, and Elk in Prehistoric Europe


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Birds of Prey
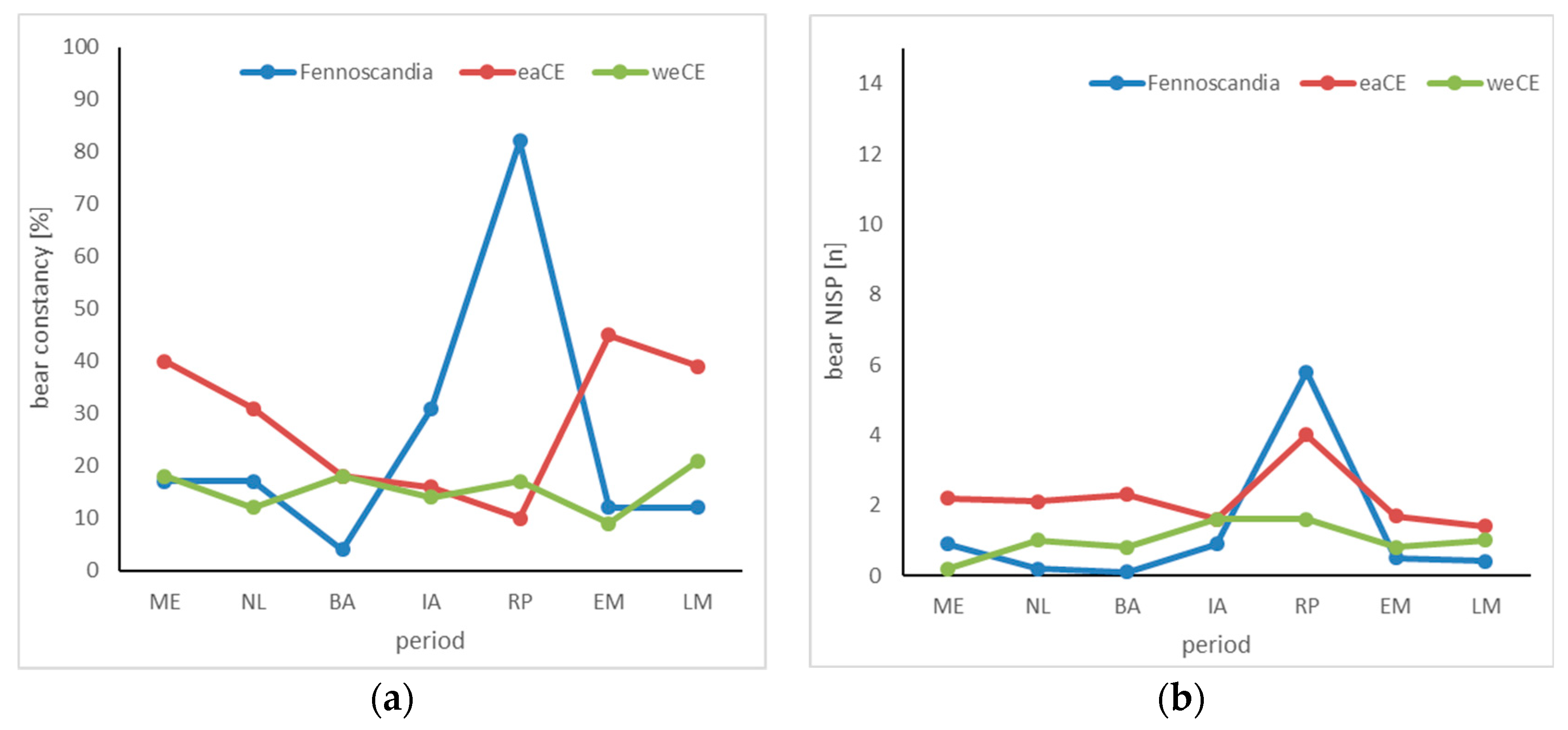
3.2. Brown Bear
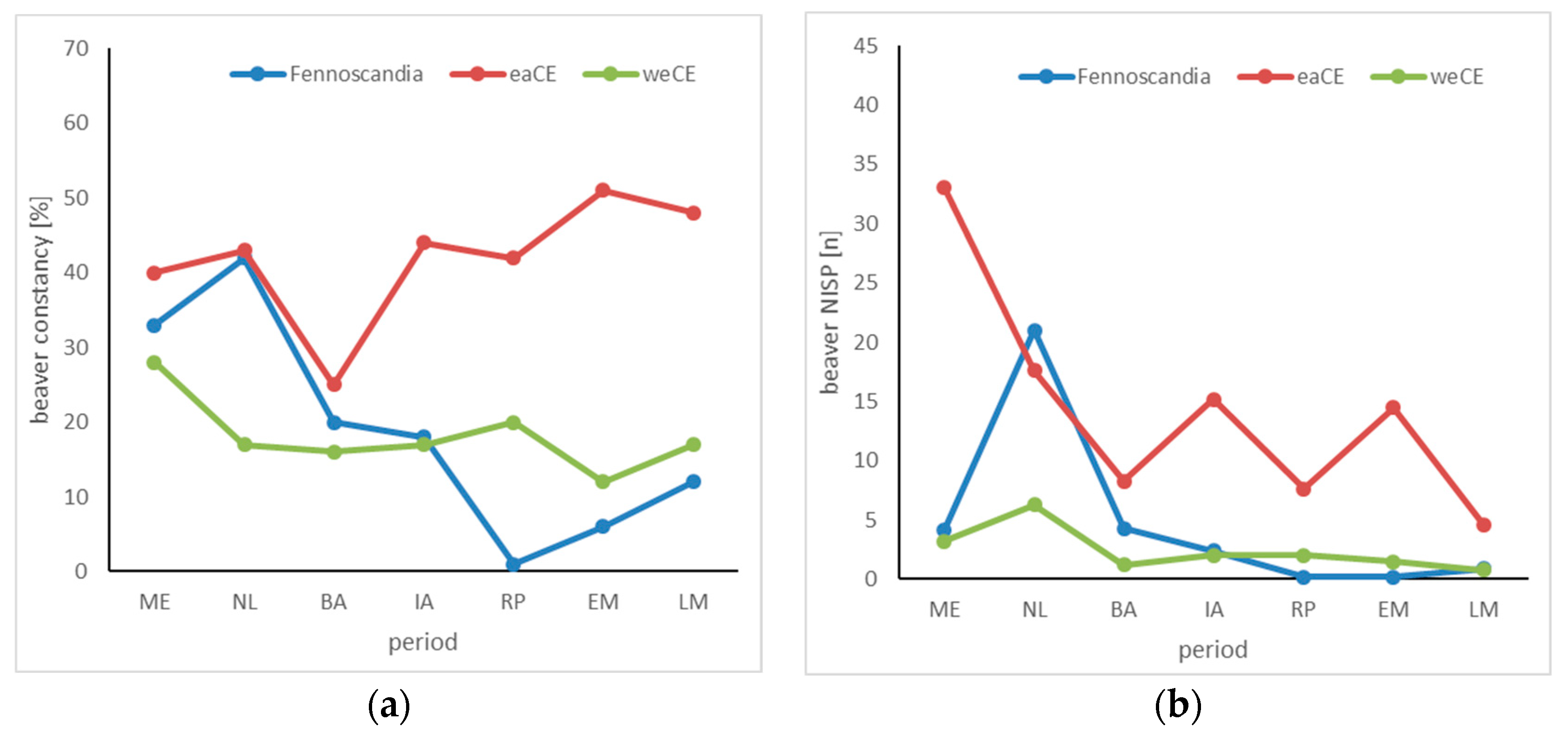
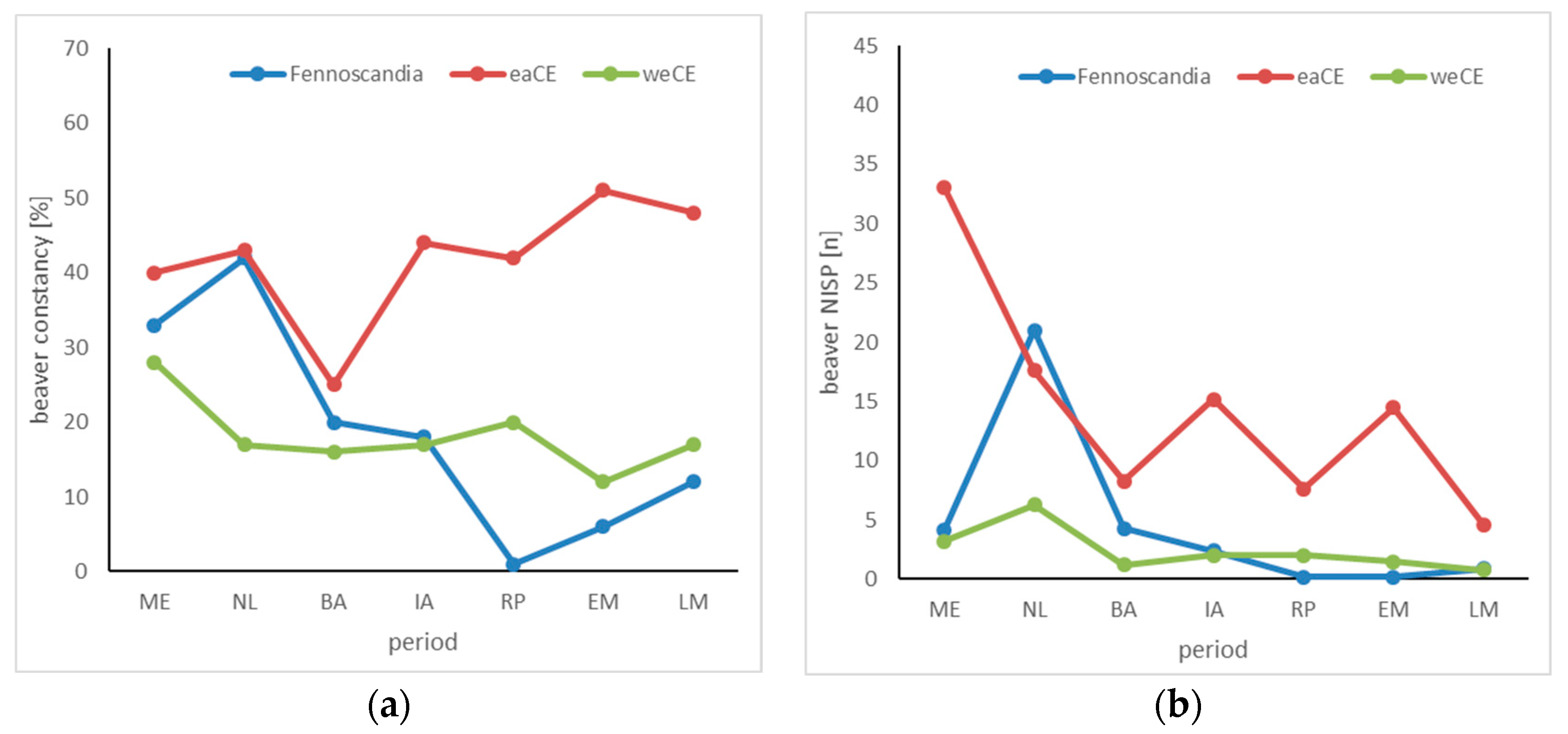
3.3. Beaver
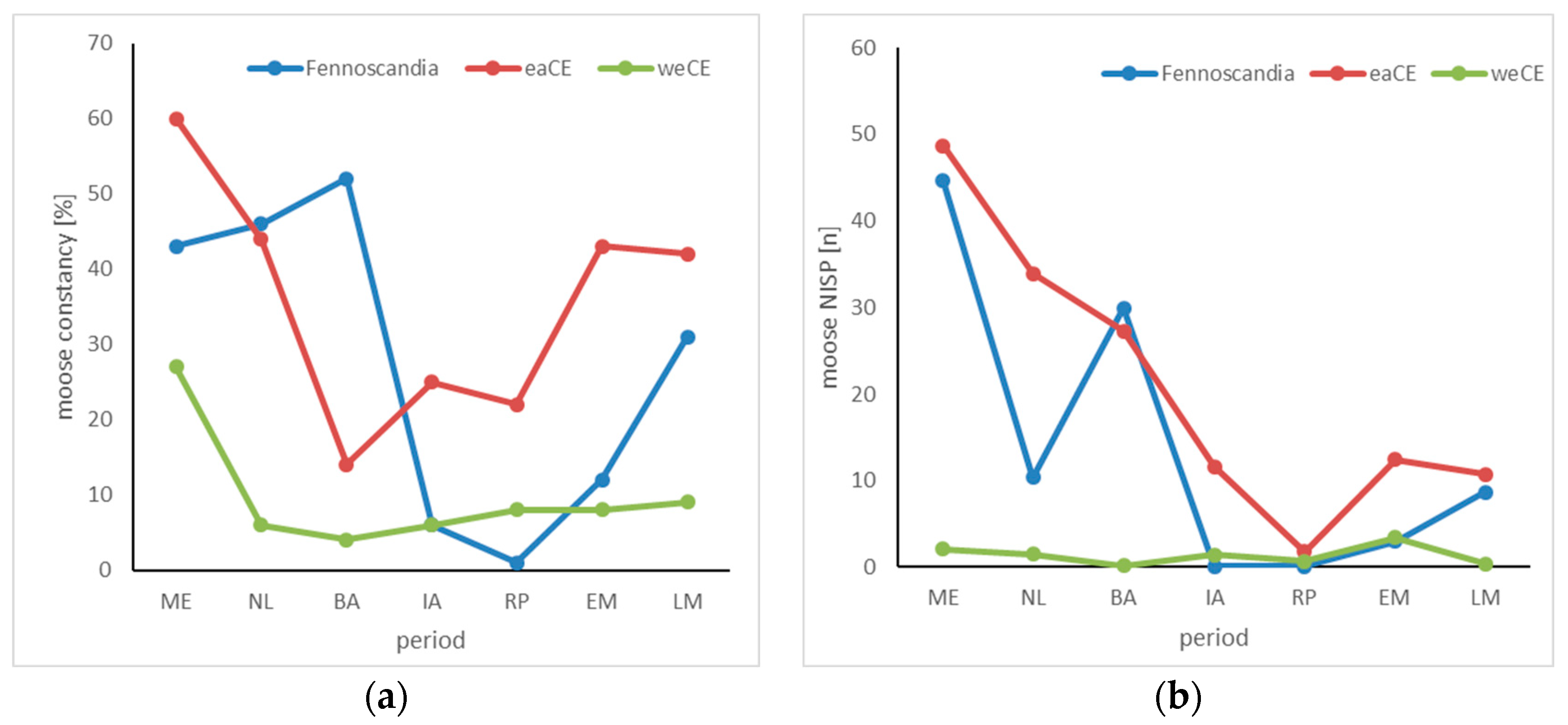
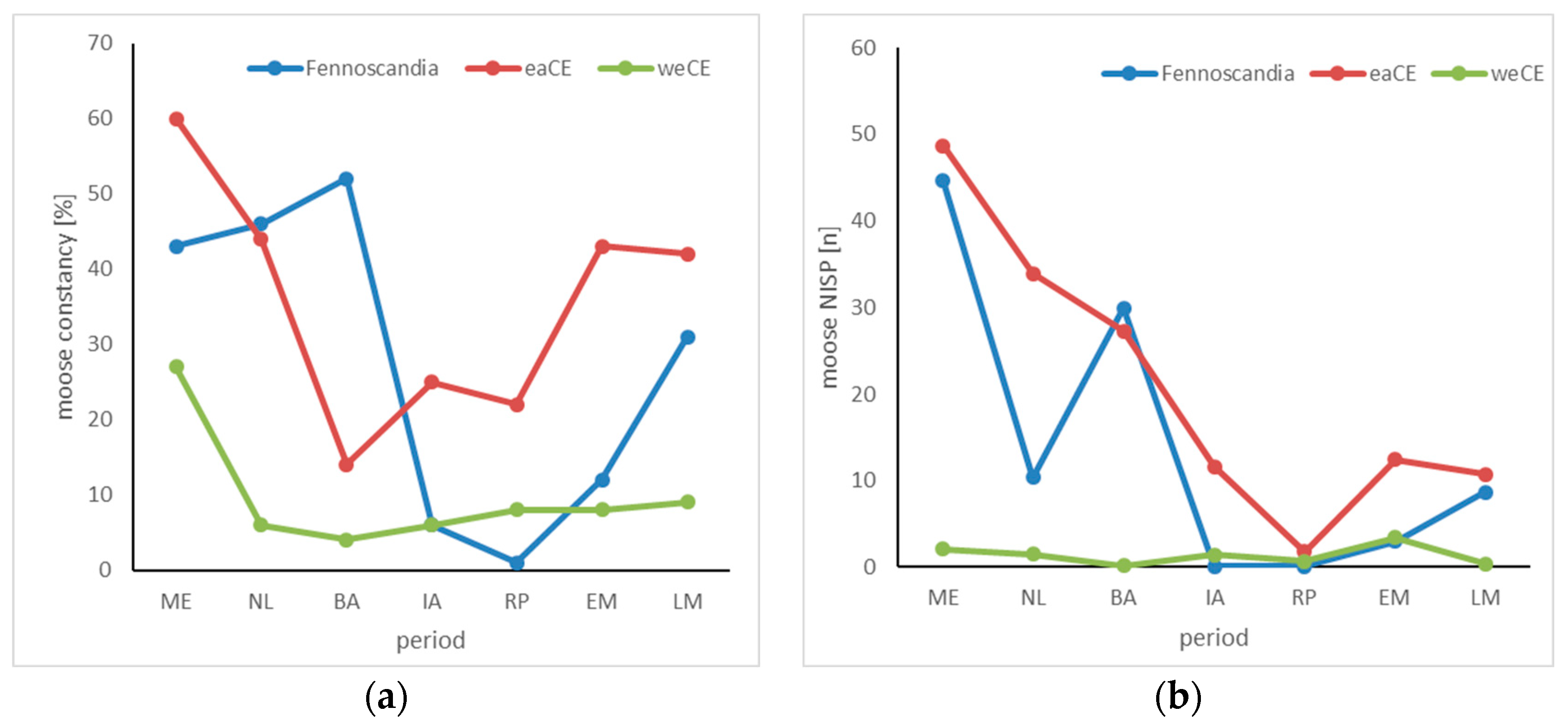
3.4. Elk
4. Discussion
4.1. Birds of Prey
4.2. Brown Bear
4.3. Beaver
4.4. Elk
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DeMello, M. Animals and Society. An introduction to Human-Animal Studies, 2nd ed.; Columbia University Press: New York, NY, USA, 2021. [Google Scholar]
- Kompatscher, G.; Spannring, R.; Schachinger, K. Human-Animal Studies. Eine Einführung für Studierende und Lehrende; Waxmann Verlag: Münster, Germany, 2021. [Google Scholar]
- Pesch, A. Die Kraft der Tiere. Völkerwanderungszeitliche Goldhalskragen und die Grundsätze Germanischer Kunst; Schnell & Steiner: Regensburg, Germany, 2015. [Google Scholar] [CrossRef]
- Singer, P. Animal Liberation; Harper Collins: New York, NY, USA, 1975. [Google Scholar]
- Regan, T. The Case for Animal Rights; University of California Press: Berkeley, CA, USA, 1983. [Google Scholar]
- Russell, N. Social Zooarchaeology: Humans and Animals in Prehistory; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar] [CrossRef]
- Overton, N.J.; Hamilakis, Y. A manifesto for a social zooarchaeology. Swans and other beings in the Mesolithic. Archaeol. Dialogues 2013, 20, 111–136. [Google Scholar] [CrossRef]
- Albarella, U.; Trentacoste, A. (Eds.) EthnoZooArchaeology: The Present and Past of Human-Animal Relationships; Oxbow: Oxford, UK, 2011. [Google Scholar]
- Armstrong-Oma, K.; Birke, L. Guest Editors’ Introduction Archaeology and Human-Animal Studies. Soc. Anim. 2013, 21, 113–119. [Google Scholar] [CrossRef]
- Boyd, B. Archaeology and Human–Animal Relations: Thinking Through Anthropocentrism. Annu. Rev. Anthropol. 2017, 46, 299–316. [Google Scholar] [CrossRef]
- Schmölcke, U.; Groß, D.; Nikulina, E.A. Bears and beavers. ‘The Browns’ in daily life and spiritual world. In Interaction without Borders. Exemplary Archaeological Research at the Beginning of the 21st Century; Eriksen, B.V., Abegg-Wigg, A., Bleile, R., Ickerodt, U., Eds.; Stiftung Schleswig-Holsteinische Landesmuseen: Schleswig, Germany, 2017; pp. 901–916. [Google Scholar]
- Schmölcke, U.; Nikulina, E.A. Humans and their Relationship to Large Carnivores in Central Europe from Foragers to Modern Times: A Survey. In The Homotherium Finds from Schöningen 13II-4: Man and Big Cats of the Ice Age; Conard, N.J., Hillgruber, K.-F., Serangeli, J., Hassmann, H., Terberger, T., Eds.; Propylaeum: Heidelberg, Germany, 2022; pp. 129–150. [Google Scholar]
- Gersmann, K.-H.; Grimm, O. (Eds.) Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Wachholtz: Kiel/Hamburg, Germany, 2018. [Google Scholar]
- Grimm, O. Raptor on the fist: Falconry, its imagery and similar motifs throughout the millennia on a global scale—A synthesis against a broader background. In Raptor on the Fist—Falconry, Its Imagery and Similar Motifs throughout the Millennia on a Global Scale; Grimm, O., Ed.; In Cooperation with Gersmann, K.-H.; Tropato, A.-L.; Wachholtz: Kiel/Hamburg, Germany, 2020; pp. 17–33. [Google Scholar]
- Grimm, O. “Bear and human”—Introduction, discussion and synthesis. In Bear and Human—Facets of a Multi-Layered Relationship from Past to Recent Times with an Emphasis on Northern Europe; Grimm, O., Ed.; In Cooperation with Groß, D.; Pesch, A.; Sundqvist, O.; Zedrosser, A.; Brepols Publishers: Turnhout, Belgium, 2023; pp. 19–46. [Google Scholar]
- Braudel, F. Histoire et sciences sociales: La longue durée. Ann. Écon. Soc. Civ. 1958, 13, 725–753. [Google Scholar] [CrossRef]
- Burke, P. The French Historical Revolution. The Annales School, 1929–1989, 2nd ed.; Polity Press: Cambridge, UK, 2004. [Google Scholar]
- Heinrich, D.; von den Driesch, A.; Benecke, N. Holozängeschichte der Tierwelt Europas.; IANUS Forschungsdatenzentrum Archäologie & Altertumswissenschaften: Berlin, Germany, 2016. [Google Scholar] [CrossRef]
- Schmölcke, U. Die Stetigkeit als archäozoologische Bewertungsmethode: Beispiele aus Paläoichthyologie (frühmittelalterlicher Seehandelsplatz Groß Strömkendorf) und Paläoökologie (Neolithikum Schleswig-Holsteins). Beiträge Archäozoologie Prähistorischer Anthropol. 2003, 4, 195–203. [Google Scholar]
- Schmölcke, U. Once more: Sample size estimation in archaeozoology. Offa 2013, 69, 239–247. [Google Scholar]
- Schmölcke, U. The history of the brown bear (Ursus arctos L.) in the northern German lowlands In Bear and Human: Facets of a Multi-Layered Relationship from Past to Recent Times, with Emphasis on Northern Europe; Grimm, O., Ed.; Brepols Publishers: Turnhout, Belgium, 2023; pp. 265–290. [Google Scholar]
- Ulrich, W.; Almeida-Neto, M.; Gotelli, N.J. A consumer’s guide to nestedness analysis. Oikos 2009, 118, 3–17. [Google Scholar] [CrossRef]
- Mannermaa, K.; Panteleyev, A.; Sablin, M. Birds in Late Mesolithic burials at Yuzhniy Oleniy ostrov (Lake Onega, western Russia)—What do they tell about humans and environment? Fennosc. Archaeol. 2008, 25, 3–25. [Google Scholar]
- Mannermaa, K. Powerful birds. The Eurasian jay (Garrulus glandarius) and the osprey (Pandion haliaetus) in hunter-gatherer burials at Zvejnieki, northern Latvia and Yuzhniy Oleniy Ostrov, northwestern Russia. Anthropozoologica 2013, 48, 189–205. [Google Scholar] [CrossRef]
- Kriiska, A.; Lõugas, L.; Lõhmus, M.; Mannermaa, K.; Johanson, K. New AMS dates from Estonian Stone Age burial sites. Est. J. Archaeol. 2007, 11, 83–121. [Google Scholar] [CrossRef]
- Ericson, P.G.P.; Tyrberg, T. The Early History of the Swedish Avifauna. A Review of the Subfossil Record and Early Written Sources; Kungl. Vitterhets historie och antikvitets Akademien: Stockholm, Sweden, 2004. [Google Scholar]
- Vretemark, M. Birds of Prey as Evidence for Falconry in Swedish Burials and Settlements (550–1500 AD). In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Grimm, O., Gersmann, K.-H., Eds.; Wachholtz Murmann: Kiel-Hamburg, Germany, 2018; pp. 827–839. [Google Scholar]
- Karpińska, K.D. On Wings to the Otherworld. Bird Remains in Viking Age Graves from Scandinavia; University of Oslo: Oslo, Norway, 2022. [Google Scholar]
- Grimm, O. The early 7th century AD Rickeby (Vallentuna) cremation grave from mid-east Sweden, understood as a three-dimensional painting/still life embodying three different kinds of falconer. In Raptor on the Fist—Falconry, Its Imagery and Similar Motifs throughout the Millennia on a Global Scale; Grimm, O., Ed.; In cooperation with Gersmann, K.-H.; Tropato, A.-L.; Wachholtz: Kiel/Hamburg, Germany, 2020; pp. 481–488. [Google Scholar]
- Gansum, T. The royal Viking Age ship grave from Gokstad in Vestfold, eastern Norway, and its link to falconry. In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz: Kiel/Hamburg, Germany, 2018; pp. 717–726. [Google Scholar]
- Ludovici, B. Chamber grave 41 from the Bockshornschanze near Quedlinburg (Central Germany): Evidence of the practice of falconry by women from the middle of the 1st century? In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz: Kiel/Hamburg, Germany, 2018; pp. 1293–1302. [Google Scholar]
- Schmölcke, U. Central European burials with birds of prey from the middle of the 1st millenium AD—A short survey of the early history of archaeology in connection with these burials. In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/Hamburg, Germany, 2018; pp. 495–505. [Google Scholar]
- Maldre, L.; Tomek, T.; Peets, J. Birds of prey from Vendel Age ship burials of Salme (c. 750 AD) and in Estonian archaeological material. In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/Hamburg, Germany, 2018; pp. 1229–1250. [Google Scholar]
- Girininkas, A.; Daugnora, L. Premodern hunting with birds of prey in the historical Lithuanian lands: Entertainment, politics or economic necessity? In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/ Hamburg, Germany, 2018; pp. 1215–1227. [Google Scholar]
- Grimm, O. From Aachen in the west to Birka in the north and Mikulčice in the east—Some archaeological remarks on bird of prey bones and falconry in premodern settlement contexts in parts of Europe (pre and post 1000 AD)’. In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/Hamburg, Germany, 2018; pp. 285–312. [Google Scholar]
- Vretemark, M. Kungahälla—Om handel och varuutbyte med animalie produkter In Kungahälla. Problem och Forskning kring Stadens Äldsta Historia; Andersson, H., Karlsson, C., Vretemark, M., Eds.; Lund Studies in Medieval Archaeology nr 28; Lunds Universitets Historiska Museum: Lund, Sweden, 2001; pp. 111–133. [Google Scholar]
- Prummel, W. Falconry in continental settlements as reflected by animal bones from the 6th to 12th centuries AD. In Hunting in Northern Europe until 1500 AD—Old Traditions and Regional Developments, Continental Sources and Continental Influences; Grimm, O., Schmölcke, U., Eds.; Wachholtz: Neumünster, Germany, 2013; pp. 357–377. [Google Scholar]
- Heinrich, D. Falconry in the Viking Age trading centre Haithabu and its successor, the medieval town of Schleswig?, with an appendix by Wolf-Rüdiger Teegen. In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/Hamburg, Germany, 2018; pp. 973–993. [Google Scholar]
- Bleile, B. Falconry among the Slavs of the Elbe? In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/Hamburg, Germany, 2018; pp. 1303–1370. [Google Scholar]
- Teegen, W.-R. The skeletons of a peregrine and a sparrowhawk and the spatial distribution of birds of prey in the Slavonic fortification of Starigard/Oldenburg (Schleswig-Holstein, northern Germany, 7th–13th centuries). In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/Hamburg, Germany, 2018; pp. 1371–1389. [Google Scholar]
- Mlíkovský, J. Die Vögel aus der frühmittelalterlichen Burg von Mikulčice, Mähren. In Studien zum Burgwall von Mikulčice 5; Poláček, L., Ed.; Spisy Archeologického Ústavu AV ČR Brno: Brno, Czech Republic, 2003; pp. 215–338. [Google Scholar]
- Mlíkovský, J. Evidence for hawking in early Medieval Czechia. Buteo 2005, 14, 53–56. [Google Scholar]
- Jost, C.A. A depiction of a falconer on a disc brooch of the 7th century from the cemetery of Münstermaifeld, District of Mayen-Koblenz, south-western Germany. In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/Hamburg, Germany, 2018; pp. 1421–1426. [Google Scholar]
- Profantova, N. Early medieval depictions of falconry from eastern Central Europe in the 8th–9th centuries. In Raptor on the Fist—Falconry, Its Imagery and Similar Motifs throughout the Millennia on a Global Scale; Grimm, O., Ed.; In Cooperation with Gersmann, K.-H.; Tropato, A.-L.; Wachholtz: Kiel/Hamburg, Germany, 2020; pp. 587–603. [Google Scholar]
- Oehrl, S. An overview of falconry in Northern Germanic and insular iconography, 6th/7th centuries AD to c. 1100 AD. In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/Hamburg, Germany, 2018; pp. 841–860. [Google Scholar]
- Oehrl, S. “I am Eagle”—Depictions of raptors and their meaning in the art of Late Iron Age and Viking Age Scandinavia (c. AD 400–1100). In Raptor on the Fist—Falconry, Its Imagery and Similar Motifs throughout the Millennia on a Global Scale; Grimm, O., Ed.; In Cooperation with Gersmann, K.-H.; Tropato, A.-L.; Wachholtz: Kiel/Hamburg, Germany, 2020; pp. 451–479. [Google Scholar]
- Hurka, F. Falconry and similar forms of hunting according to ancient Greco-Roman sources. In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/Hamburg, Germany, 2018; pp. 685–698. [Google Scholar]
- Schmölcke, U. What about Exotic Species? Significance of Remains of Strange and Alien Animals in the Baltic Sea Region, Focusing on the Period from the Viking Age to High Medieval Times (800–1300 CE). Heritage 2022, 5, 3864–3880. [Google Scholar] [CrossRef]
- Van den Abeele, B. “On the dunghill”: The dead hawk in medieval Latin and French moralising literature. In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/Hamburg, Germany, 2018; pp. 523–537. [Google Scholar]
- Reichstein, H.; Pieper, H. Untersuchungen an Skelettresten von Vögeln aus Haithabu (Ausgrabung 1966–1969) (Neue Ausgrabungen in Haithabu, Bericht 22); Wachholtz: Neumünster, Germany, 1986. [Google Scholar]
- Ehrlich, F.; Piličiauskienė, G.; Urbonaitė-Ubė, M.; Rannamäe, E. The Meaning of Eagles in the Baltic Region. A Case Study from the Castle of the Teutonic Order in Klaipėda, Lithuania (13th–14th Century). Archaeol. Litu. 2020, 21, 59–78. [Google Scholar] [CrossRef]
- Beermann, S. Bärenkrallen und Bärenfelle in Brand- und Körpergräbern der vorrömischen Eisenzeit bis Völkerwanderungszeit in Mittel- und Nordeuropa; Verlag Dr. Rudolf Steinert: Bonn, Germany, 2016. [Google Scholar]
- Ljungkjvist, J.; Lindholm, K.-J. Bears and the Viking Age transition in Sweden. In Bear and Human—Facets of a Multi-Layered Relationship from Past to Recent Times with an Emphasis on Northern Europe; Grimm, O., Ed.; In Cooperation with Groß, D.; Pesch, A.; Sundqvist, O.; Zedrosser, A.; Brepols Publishers: Turnhout, Belgium, 2023; pp. 387–404. [Google Scholar]
- Wamers, E. Bestattungen mit Bärenfell. In Franconofurd 2. Das Bi-Rituelle Kinderdoppelgrab der späten Merowingerzeit unter der Frankfurter Bartholomäuskirche (‘Dom‘); Wamers, E., Ed.; Schnell und Steiner: Regensburg, Germany, 2015; pp. 180–192. [Google Scholar]
- Wamers, E. Von Bären und Männern. Berserker, Bärenkämpfer und Bärenführer im frühen Mittelalter. Z. Archäologie Mittelalt. 2009, 37, 1–46. [Google Scholar]
- Grimm, O. Bear-skins in northern European burials and some remarks on other bear-related furnishings in the north and middle of Europe in the 1st millennium AD. In Hunting in Northern Europe until 1500 AD—Old Traditions and Regional Developments, Continental Sources and Continental Influences; Grimm, O., Schmölcke, U., Eds.; Wachholtz: Neumünster, Germany, 2013; pp. 277–296. [Google Scholar]
- Grimm, O. (Ed.) Bear and Human—Facets of a Multi-Layered Relationship from Past to Recent Times with an Emphasis on Northern Europe; In Cooperation with Groß, D.; Pesch, A.; Sundqvist, O.; Zedrosser, A.; Brepols Publishers: Turnhout, Belgium, 2023; Available online: https://www.brepolsonline.net/action/showBook?doi=10.1484/M.TANE-EB.5.133678 (accessed on 26 January 2024).
- Grimm, O. Bear skin burials revisited: Norway and Sweden, mainly Migration Period. In Bear and Human—Facets of a Multi-Layered Relationship from Past to Recent Times with an Emphasis on Northern Europe; Grimm, O., Ed.; In Cooperation with Groß, D.; Pesch, A.; Sundqvist, O.; Zedrosser, A.; Brepols Publishers: Turnhout, Belgium, 2023; pp. 533–546. [Google Scholar]
- Schönfelder, M. Bear-claws in Germanic graves. Oxf. J. Archaelogy 1994, 13, 217–227. [Google Scholar] [CrossRef]
- Henriksen, M.B. Brudager Mark—En Romertidsgravplads nær Gudme på Sydøstfyn 1. Fynske jernaldergrave 6 = Fynske studier 22; Forlaget Odensy byers Museer: Odense, Denmark, 2009. [Google Scholar]
- Iregren, E. Sámi bear graves—Results from archaeological and zooarchaeological excavations and analyses in the Swedish part of Sápmi. In Bear and Human—Facets of a Multi-Layered Relationship from Past to Recent Times with an Emphasis on Northern Europe; Grimm, O., Ed.; In Cooperation with Groß, D.; Pesch, A.; Sundqvist, O.; Zedrosser, A.; Brepols Publishers: Turnhout, Belgium, 2023; pp. 547–586. [Google Scholar]
- Sommerseth, I. Sámi bear graves in Norway—Hidden sites and rituals. In Bear and Human—Facets of a Multi-Layered Relationship from Past to Recent Times with an Emphasis on Northern Europe; Grimm, O., Ed.; In Cooperation with Groß, D.; Pesch, A.; Sundqvist, O.; Zedrosser, A.; Brepols Publishers: Turnhout, Belgium, 2023; pp. 587–602. [Google Scholar]
- Magnell, O. Bear bones from the Viking Age cult place at Frösö church—The unifying factor in bear-human relationships in Viking Age Jämtland, northern Sweden. In Bear and Human—Facets of a Multi-Layered Relationship from Past to Recent Times with an Emphasis on Northern Europe; Grimm, O., Ed.; In Cooperation with Groß, D.; Pesch, A.; Sundqvist, O.; Zedrosser, A.; Brepols Publishers: Turnhout, Belgium, 2023; pp. 429–452. [Google Scholar]
- Schmölcke, U.; Nikulina, E.A. Mesolithic beaver hunting station or base camp of supra-regional Stone Age fur trade? New archaeozoological and archaeogenetic results from Dąbki 9. In The Dąbki Site in Pomerania and the Neolithisation of the North European Lowlands (c. 5000-3000 calBC); Kabaciński, J., Hartz, S., Raemaekers, D.C.M., Terberger, T., Eds.; Marie Leidorf: Rahden/Westf, Germany, 2015; pp. 65–86. [Google Scholar]
- Ekman, J.; Iregren, E. Archaeozoological Investigations in Northern Sweden; Kungl. Vitterhets Historie och Antikvitets Akademien: Stockholm, Sweden, 1984. [Google Scholar]
- Ukkonen, P. Osteological analysis of the refuse fauna in the Lake Saimaa area. Hels. Pap. Archaeol. 1996, 8, 63–91. [Google Scholar]
- Ukkonen, P. Early in the North—Utilization of Animal Resources in Northern Finland during Prehistory. Iskos 2004, 13, 103–130. [Google Scholar]
- Ewersen, J. Die Tierknochenfunde aus der Neolithischen Siedlung Heidmoor, Kr. Segeberg, unter Besonderer Berücksichtigung Wirtschaftshistorischer Aspekte; Wachholtz: Neumünster, Germany, 2007. [Google Scholar]
- Hübner, K.-D.; Saur, R.; Reichstein, H. Die Säugetierknochen der neolithischen Seeufersiedlung Hüde I. In Palynologische und Säugetierkundliche Untersuchungen zum Siedlungsplatz Hüde I am Dümmer Landkreis Diepholz; Göttinger Schriften zur Vor- und Frühgeschichte 23; Wachholtz: Neumünster, Germany, 1988; pp. 35–142. [Google Scholar]
- Gurina, N.N. Oleneostrovski Mogil’nik. In Materialy i Issledovaniya po Arkheologgi SSSR; Akademia Nauk SSSR: Moscow, Sowjet Union, 1956. [Google Scholar]
- Møhl, U. 1978: Elsdyrskeletterne fra Skottemark og Favrbo. Sik og brug ved borealtidens jagter. Aarbøger Nord. Oldkynd. Hist. 1980, 1978, 5–32. [Google Scholar]
- Møller Hansen, K.; Buch Pedersen, K. With or without Bones. Late Palaeolithic Hunters in South Zealand. In Across the Western Baltic; Møller Hansen, K., Buch Pedersen, K., Eds.; Sydsjællands Museum: Vordingborg, Denmark, 2006; pp. 93–110. [Google Scholar]
- Møller Hansen, K.; Brinch Petersen, E.; Aaris-Sørensen, K. Filling the gap: Early Preboreal Maglemose elk deposits at Lundby, Sjælland, DK. In Hunters in a Changing World. Environment and Archaeology of the Pleistocene—Holocene Transition (ca. 11000–9000 B.C.) in Northern Central Europe; Terberger, T., Eriksen, B.V., Eds.; Marie Leidorf: Rahden, Germany, 2004; pp. 75–84. [Google Scholar]
- Leduc, C. A specialized Early Maglemosian site at Lundby Mose (Zealand, Denmark): A contribution to the understanding of Maglemosian patterns of animal resource exploitation. J. Archaeol. Sci. 2014, 41, 199–213. [Google Scholar] [CrossRef]
- Sjöström, A. 1998: Ringsjöholm. A Boreal—Early Atlantic Settlement in Central Scania, Sweden. Lund Archaeol. Rev. 1998, 1997, 5–20. [Google Scholar]
- Larsson, L.; Sázelová, S.; Novák, M.; Mizerová, A. On the trail of the elks: Finds from the preboreal in southern Sweden. In Forgotten Times and Spaces: New Perspectives in Paleoanthropological, Paleoetnological and Archeological Studies; Sázelová, S., Novák, M., Mizerová, A., Eds.; Institute of Archeology of the Czech Academy of Sciences, Masaryk University: Brno, Czech Republic, 2015; pp. 468–478. [Google Scholar] [CrossRef]
- Larsson, L. Animals and animal depictions in the early Holocene of Northern Europe based on an antler adze with deer depictions. In Tiere und Tierdarstellungen in der Archäologie. Beiträge zum Kolloquium in Gedenken an Torsten Capelle; Brieske, V., Dickers, A., Rind, M.M., Eds.; Aschendorff: Münster, Germany, 2017; pp. 63–73. [Google Scholar]
- Bolin, H. Animal magic: The mythological significance of elks, boats and humans in north Swedish rock art. J. Mater. Cult. 2000, 5, 153–176. [Google Scholar] [CrossRef]
- Sjöstrand, Y. Should I stay or should I go? On the meaning of variations among mobile and stable elk motifs at Nämforsen, Sweden. In Changing Pictures. Rock Art Traditions and Visions in Northern Europe; Goldhahn, J., Jones, A., Fuglestvedt, I., Eds.; Oxbow Books: Oxford, UK, 2010; pp. 139–154. [Google Scholar]
- Sognnes, K. Land of elks–sea of whales: Landscapes of the Stone Age rock-art in central Scandinavia. In European Landscapes of Rock-Art; Chippindale, C., Nash, G., Eds.; Routledge: London, UK, 2002; pp. 195–212. [Google Scholar]
- Mantere, V. The Relationship Between Humans and Elks (Alces alces) in Northern Europe c. 12000–1200 calBC; Department of Archaeology, University of Turku: Turku, Finland, 2023; Available online: https://www.utupub.fi/handle/10024/175889 (accessed on 26 January 2024).
- Kabaciński, J.; Hartz, S.; Terberger, T. Elks in the early Stone Age art of the northern Lowlands. Praehist. Z. 2011, 86, 151–164. [Google Scholar] [CrossRef]
- Deinet, S.; Ieronymidou, C.; McRae, L.; Burfield, I.J.; Foppen, R.P.; Collen, B.; Böhm, M. Wildlife Comeback in Europe: The Recovery of Selected Mammal and Bird Species; Zoological Society of London: London, UK, 2013; Available online: https://rewildingeurope.com/wp-content/uploads/2013/11/Wildlife-Comeback-in-Europe-the-recovery-of-selected-mammal-and-bird-species.pdf (accessed on 26 January 2024).
- Schmölcke, U.; Zachos, F.E. Holocene distribution and extinction of the elk (Alces alces, Cervidae) in Central Europe. Mamm. Biol. 2005, 70, 329–344. [Google Scholar] [CrossRef]
- Benecke, N. Nacheiszeitliche Vorkommen seltener Säugetierarten zwischen Elbe und Oder. In Animals and Man in the Past: Essays in Honour of Dr. A.T. Clason; Buitenhuis, H., Prummel, W., Eds.; ARC-Publicatie: Groningen, The Netherlands, 2001; pp. 192–208. [Google Scholar]
- Morin, E.; Laroulandie, V. Presumed symbolic use of diurnal raptors by Neanderthals. PLoS ONE 2012, 7, e32856. [Google Scholar] [CrossRef]
- Amkreutz, L.; Corbey, R. An eagle-eyed perspective. Haliaeetus albicilla in the Mesolithic and Neolithic of the Lower Rhine Area. In Between Foraging and Farming. An Extended Broad Spectrum of Papers Presented to Leendert Louwe Kooijmans; Fokkens, H., Coles, B.J., Van Gijn, A.L., Kleijne, J.P., Ponjee, H.H., Eds.; Leiden University: Leiden, The Netherlands, 2008; pp. 167–180. Available online: https://hdl.handle.net/1887/43060 (accessed on 26 January 2024).
- Zeiler, J.T. The white-tailed eagle (Haliaeetus albicilla) in the Netherlands: Changing landscapes, changing attitudes. Archaeol. Anthropol. Sci. 2019, 11, 6371–6375. [Google Scholar] [CrossRef]
- Bednarek, W. Emotions and motivation of the falconer and his relationship with the trained raptor—Attempt at an evolutionary-biological interpretation. In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel Hamburg, Germany, 2018; pp. 285–312. [Google Scholar]
- Richter, T. Practicalities of falconry, as seen by a present-day falconer. In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz: Kiel/Hamburg, Germany, 2018; pp. 55–86. [Google Scholar]
- Dusil, S. Falconry in the mirror of normative sources from Central Europe (5th to 19th centuries). In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/Hamburg, Germany, 2018; pp. 507–521. [Google Scholar]
- Giese, M. Legal regulations on hunting in the barbarian lawcodes of the early middle ages. In Hunting in Northern Europe until 1500 AD—Old Traditions and Regional Developments, Continental Sources and Continental Influences; Grimm, O., Schmölcke, U., Eds.; Wachholtz: Neumünster, Germany, 2013; pp. 480–504. [Google Scholar]
- Carstens, L. Land of the hawk: Old Norse literary sources about the knowledge and practice of falconry. In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/Hamburg, Germany, 2018; pp. 799–826. [Google Scholar]
- Haley-Halinski, K. Birds and Humans in the Old Norse World, c. 600-1500 AD. Ph.D. Thesis, University of Cambridge, Cambridge, UK, 2022. Available online: https://api.repository.cam.ac.uk/server/api/core/bitstreams/f0340561-f564-4106-a2d2-77ad18972d33/content (accessed on 26 January 2024).
- Boccassini, D. Falconry as royal ‘delectation’: Understanding the art of taming and its philosophical foundation in 12th and 13th--century Europe. In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/ Hamburg, Germany, 2018; pp. 367–387. [Google Scholar]
- Giese, M. The ‘de arte venandi cum avibus’ of Emperor Frederick II. In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/Hamburg, Germany, 2018; pp. 1459–1469. [Google Scholar]
- Hadjinicolaou, Y. Flying images. Falconry’s visual afterlife. In Raptor on the Fist—Falconry, Its Imagery and Similar Motifs throughout the Millennia on a Global Scale; Grimm, O., Ed.; In Cooperation with Gersmann, K.-H.; Tropato, A.-L.; Wachholtz: Kiel/Hamburg, Germany, 2020; pp. 855–878. [Google Scholar]
- Horobin, D. The pen and the peregrine: Literary influences on the development of British falconry (8th century to the present). In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/Hamburg, Germany, 2018; pp. 1055–1088. [Google Scholar]
- Karaskova-Hesry, O. Mary of Burgundy (1457–1482): Lady with a hawk. In Raptor on the Fist—Falconry, Its Imagery and Similar Motifs throughout the Millennia on a Global Scale; Grimm, O., Ed.; In Cooperation with Gersmann, K.-H.; Tropato, A.-L.; Wachholtz: Kiel/Hamburg, Germany, 2020; pp. 819–833. [Google Scholar]
- Lorenz, K. Der Kumpan in der Umwelt des Vogels. Der Artgenosse als auslösendes Moment sozialer Verhaltungsweisen [The companion in the bird’s world. The fellow-member of the species as releasing factor of social behavior]. J. Ornithol. 1935, 83, 137–213. [Google Scholar] [CrossRef]
- Spindler, K. Falknerei in Archäologie und Geschichte unter besonderer Berücksichtigung der Beizjagd in Tirol; Nearchos Sonderheft 3; Universitätsbuchhandlung Golf Verlag: Innsbruck, Austria, 1998. [Google Scholar]
- Grimm, O.; Gersmann, K.-H. Introduction, discussion and summary: Raptor and human—Falconry and bird symbolism throughout the millennia on a global scale. In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz Verlag: Kiel/Hamburg, Germany, 2018; pp. 18–25. [Google Scholar]
- Gersmann, K.-H. Some thoughts on the emergence and function of falconry from the perspective of a practicing falconer. In Raptor and Human—Falconry and Bird Symbolism throughout the Millennia on a Global Scale; Gersmann, K.-H., Grimm, O., Eds.; Wachholtz: Kiel/Hamburg, Germany, 2018; pp. 141–146. [Google Scholar]
- Ehrlich, F.; Piličiauskienė, G.; Blaževičius, P. Bird remains from Vilnius Lower Castle, Lithuania (13th–19th centuries) reveal changes in social status and unusual bird pathologies. Int J Osteoarch 2022, 33, 731–741. [Google Scholar] [CrossRef]
- Paulsen, H. Pfeil und Bogen in Haithabu (Berichte über die Ausgrabungen in Haithabu, Bericht 33); Wachholtz: Neumünster, Germany, 1999; pp. 93–143. [Google Scholar]
- Bieder, R.E. Bear; Reaction books: London, UK, 2005. [Google Scholar]
- Brunner, B. Bär und Mensch. Die Geschichte einer Beziehung; Wissenschaftliche Buchgesellschaft: Darmstadt, Germany, 2010. [Google Scholar]
- Oehrl, S. Bear hunting and its ideological context (as background for the interpretation of bear claws and other remains of bear in Germanic graves of the 1st millennium AD). In Hunting in Northern Europe until 1500 AD—Old Traditions and Regional Developments, Continental Sources and Continental Influences; Grimm, O., Schmölcke, U., Eds.; Wachholtz: Neumünster, Germany, 2013; pp. 297–332. [Google Scholar]
- Germonpré, M.; Hämäläinen, R. Fossil Bear Bones in the Belgian Upper Paleolithic: The Possibility of a Proto Bear-Ceremonialism. Arct. Anthropol. 2007, 44, 1–30. [Google Scholar] [CrossRef]
- Kunst, G.K.; Pacher, M. Brown bear remains in prehistoric and early historic societies: Case studies from Austria. Berichte Geol. Bundesanst. 2019, 132, 89–121. Available online: https://opac.geologie.ac.at/ais312/dokumente/BR0132.pdf (accessed on 26 January 2024).
- Schmölcke, U. Mesolithic hunters in mixed oak forests. Differences in hunting strategy and hunting behaviour. In Changing Identity in a Changing World. Current Studies on the Stone Age around 4000 BCE; Groß, D., Rothstein, M., Eds.; Sidestone Press: Leiden, The Netherlands, 2023; pp. 201–212. [Google Scholar] [CrossRef]
- Gramsch, B. Mesolithische Knochenartefakte von Friesack, Fundplatz 4, Lkr. Havelland. Veröffentlichungen Brand. Landesarchäologie 2012, 45, 7–59. [Google Scholar]
- Rydving, H. The ‘Bear Ceremonial’ and Bear Rituals among the Khanty and the Sámi. Temenos. Nord. J. Comp. Relig. 2010, 46, 31–52. [Google Scholar] [CrossRef]
- Piludu, V.M. The songs and rituals of the Finno-Karelian bear hunt: Gifts, seduction and mimesis in the forest. In Bear and Human—Facets of a Multi-Layered Relationship from Past to Recent Times with an Emphasis on Northern Europe; Grimm, O., Ed.; In cooperation with Groß, D.; Pesch, A.; Sundqvist, O.; Zedrosser, A.; Brepols Publishers: Turnhout, Belgium, 2023; pp. 693–722. [Google Scholar]
- Piludu, V.M. The Finno-Karelian bear feast and wedding: The bruin as a guest of honour of the village. In Bear and Human—Facets of a Multi-Layered Relationship from Past to Recent Times with an Emphasis on Northern Europe; Grimm, O., Ed.; In Cooperation with Groß, D.; Pesch, A.; Sundqvist, O.; Zedrosser, A.; Brepols Publishers: Turnhout, Belgium, 2023; pp. 723–744. [Google Scholar]
- Piludu, V.M. The Finno-Karelian bear skull rituals: Bringing the bruin home to ensure its regeneration. In Bear and Human—Facets of a Multi-Layered Relationship from Past to Recent Times with an Emphasis on Northern Europe; Grimm, O., Ed.; In Cooperation with Groß, D.; Pesch, A.; Sundqvist, O.; Zedrosser, A.; Brepols Publishers: Turnhout, Belgium, 2023; pp. 745–760. [Google Scholar]
- Hallowell, I.A. Bear Ceremonialism in the Northern Hemisphere. Am. Anthropol. 1926, 28, 1–175. [Google Scholar] [CrossRef]
- Nedoma, R. Germanic “bear” and Germanic personal names before c. AD 1000 with elements referring to “bear”. In Bear and Human—Facets of a Multi-Layered Relationship from Past to Recent Times with an Emphasis on Northern Europe; Grimm, O., Ed.; In Cooperation with Groß, D.; Pesch, A.; Sundqvist, O.; Zedrosser, A.; Brepols Publishers: Turnhout, Belgium, 2023; pp. 921–932. [Google Scholar]
- Udolph, J. The Slavic word for “bear”. In Bear and Human—Facets of a Multi-Layered Relationship from Past to Recent Times with an Emphasis on Northern Europe; Grimm, O., Ed.; In Cooperation with Groß, D.; Pesch, A.; Sundqvist, O.; Zedrosser, A.; Brepols Publishers: Turnhout, Belgium, 2023; pp. 933–938. [Google Scholar]
- Hussain, S.T.; Brusgaard, N.Ø. Human-beaver cohabitation in the Early and Mid-Holocene of Northern Europe: Re-visiting Mesolithic material culture and ecology through a multispecies lens. Holocene 2024, 34, 25–55. [Google Scholar] [CrossRef]
- Razmaitė, V.; Šveistienė, R.; Švirmickas, G.J. Compositional characteristics and nutritional quality of Eurasian beaver (Castor fiber) meat. Czech J. Food Sci. 2011, 29, 480–486. [Google Scholar] [CrossRef]
- Jankowska, B.; Żmijewski, T.; Kwiatkowska, A.; Korzeniowski, W. The composition and properties of beaver (Castor fiber) meat. Eur. J. Wildl. Res. 2005, 51, 283–286. [Google Scholar] [CrossRef]
- O’Shea, J.; Zvelebil, M. Oleneostrovski mogilnik: Reconstructing the social and economic organization of prehistoric foragers in northern Russia. J. Anthropol. Archaeol. 1984, 3, 1–40. [Google Scholar] [CrossRef]
- Grünberg, J.M. Animals in Mesolithic Burials in Europe. Anthropozoologica 2013, 48, 231–253. [Google Scholar] [CrossRef]
- Jacobs, K. Returning to Oleni’ostrov: Social, Economic, and Skeletal Dimensions of a Boreal Forest Mesolithic Cemetery. J. Anthropol. Archaeol. 1995, 4, 359–403. [Google Scholar] [CrossRef]
- Eriksson, G.; Lõugas, L.; Zagorska, I. Stone Age hunter–fisher–gatherers at Zvejnieki, northern Latvia. Before Farming 2003, 2003, 1–25. [Google Scholar] [CrossRef]
- Myrberget, S. The beaver in Norway. Acta Theriol. 1967, 12, 17–26. [Google Scholar] [CrossRef]
- Wróbel, M. Population of Eurasian beaver (Castor fiber) in Europe. Glob. Ecol. Conserv. 2020, 23, e01046. [Google Scholar] [CrossRef]
- Aaris-Sørensen, K. Dynamics of the mammalian fauna in Denmark. Foss. Strat. 2009, 57, 1–59. [Google Scholar]
- Schmölcke, U.; Ickerodt, U. Die Entstehung der schleswig-holsteinischen Kulturlandschaften—Eine Skizze. In Historische Kulturlandschaften in Schleswig-Holstein. Planung—Gestaltung—Vermittlung; Weltecke, C., Ickerodt, U., Eds.; Archäologisches Landesamt Schleswig-Holstein: Schleswig, Germany, 2018; pp. 13–33. [Google Scholar]
- Luik, H. Beaver in the Economy and Social Communication of the Inhabitants of South Estonia in the Viking Age (800–1050 AD). In Bestial Mirrors. Using Animals to Construct Human Identities in Medieval Europe; Pluskowski, A., Kunst, G.K., Kucera, M., Bietak, M., Hein, I., Eds.; Vienna Institute for Archaeological Science: Wien, Austria, 2010; pp. 46–54. [Google Scholar]
- Ebbesen, K. Terningspil i yngre stenalder. Kuml 1996, 1995/96, 101–112. [Google Scholar] [CrossRef]
- Nikulina, E.A.; Schmölcke, U. Spiel mit dem Knochen. Abent. Archäol. 2007, 5, 12–15. [Google Scholar]
- Nikulina, E.A.; Schmölcke, U. Les osselets, ancêtres du jeu de dés. Pour Sci. 2008, 365, 40–43. [Google Scholar]
- Jonuks, T. Archaeology of religion—Possibilities and prospects. Est. J. Archaeol. 2005, 9, 32–59. [Google Scholar] [CrossRef]
- Cole, B. Beavers in Britain’s Past; Oxbow: Oxford, UK, 2006. [Google Scholar]
- Blair, J. The Church in Anglo-Saxon Society; Oxbow: Oxford, UK, 2005. [Google Scholar]
- Wade, E. A Story About Beaver Slavery in Arabic, Northern European, and Persian Sources. ANQ A Q. J. Short Artic. Notes Rev. 2023, 1–4. [Google Scholar] [CrossRef]
- Speck, F.G. Mistassini hunting territories in the Labrador Peninsula. Am. Anthropol. 1923, 25, 452–471. Available online: https://www.jstor.org/stable/660044 (accessed on 26 January 2024). [CrossRef]
- Cox, B.A. Native People, Native Lands: Canadian Indians, Inuit and Metis; McGill-Queen’s University Press: Ottawa, ON, Canada, 1988. [Google Scholar]
- Tanner, A. Bringing Home Animals: Religious Ideology and Mode of Production of the Mistassini Cree Hunters; St. Martin’s Press: New York, NY, USA, 1979. [Google Scholar]
- Pentikäinen, J. Saamelaiset: Pohjoisen Kansan Mytologia; Suomalaisen Kirjallisuuden Seura: Helsinki, Finland, 1995. [Google Scholar]
- Tarkka, L. Rajarahvaan Laulu: Tutkimus Vuokkiniemen Kalevalamittaisesta Runokulttuurista 1821–1921 [The Song of the Border People: The Kalevala-Meter Rune-singing Culture in Vuokkiniemi 1821–1921]; Suomalaisen Kirjallisuuden Seura: Helsink, Finland, 2005. [Google Scholar]
- Morgan, R.G. Beaver Ecology/Beaver Mythology. Ph.D. Thesis, University of Alberta, Edmonton, AB, Canada, 1991. [Google Scholar]
- Zhilin, M. Beaver mandible tools in the Mesolithic of the forest zone of Eastern Europe and Urals. J. Archaeol. Sci. Rep. 2020, 30, 102199. [Google Scholar] [CrossRef]
- Kluge, F. Etymologisches Wörterbuch der Deutschen Sprache; De Gruyter: Berlin, Germany, 2002. [Google Scholar] [CrossRef]
- Pokorny, J. Indogermanisches Etymologisches Wörterbuch; Francke Verlag: Tübingen, Germany, 1959; Available online: https://indo-european.info/pokorny-etymological-dictionary/whnjs.htm (accessed on 26 January 2024).
- Dresler, P.; Dreslerová, G.; Doláková, N.; Kočár, P.; Kočárová, R. Beaver as proof of the change of natural environment and economy of the first half of the 10th century AD. Archaeol. Austriaca 2022, 106, 117–136. [Google Scholar] [CrossRef]
- Gorbachev, A.Y. Transmission of the Names of Mythical Creatures from Belarusian to English: Modern Approaches. In Proceedings of the 77th Scientific Conference of Students and Postgraduates of the Belarusian State University: Materials of the Conference, Minsk, Belarus, 17–22 April 2021; pp. 443–447. (In Russian). [Google Scholar]
- Jääskeläinen, E. Human-beaver Engagements Seen Through Multiperiod Settlement Sites at Rautalampi, Hämeenniemi and Kitee, Hiidenniemi, Finland. Fennosc. Archaeol. 2023, 40, 39–56. [Google Scholar] [CrossRef]
- Mykhailova, N.R.; Yanevich, O.O. Ritual on the Rock. Reflection of Totemic Rites of the Deer Cult in the Rock Art of Northern Eurasia. Strat. Plus 2022, 2, 233–246. [Google Scholar] [CrossRef]
- Mykhailova, N. Elk-boat Depictions in the Ethnoarchaeological Context. Archaeol. Litu. 2022, 23, 135–147. [Google Scholar] [CrossRef]
- Lindgren, B. Hällbilder. Kosmogoni och verklighet [Rock art. Cosmogony and reality]. In Tidsspår: Forntidsvärld och Gränslöst Kulturarv; Bergvall, M., George, O., Eds.; Länsmuseet Västernorrland: Härnösand, Sweden, 2001; pp. 43–81. [Google Scholar]
- Lindgren, B. Hällmålningar. Ett uttryck för materiella och immateriella dimensioner [Rock art. An expression of material and immaterial dimensions]. In Hällbilder & Hällbildernas Rum (Studier i Regional Arkeologi 2); Klang, L., Lindgren, B., Ramqvist, P.H., Eds.; Regional Arkeologi: Örnsköldsvik, Sweden, 2002; pp. 55–75. [Google Scholar]
- Mantere, V.N.; Kashina, E.A. Elk-Head Staffs in Prehistoric North-Eastern Europe and North-Western Russia—Signs of Power and Prestige? Oxf. J. Archaeol. 2020, 39, 2–18. [Google Scholar] [CrossRef]
- Mykhailova, N. The Antlered Mother: From the Paleolithic to the Modern Era. In The History and Environmental Impacts of Hunting Deities. Supernatural Gamekeepers and Animal Masters; Chacon, R.J., Ed.; Springer: Cham, Switzerland, 2023; pp. 9–25. [Google Scholar] [CrossRef]
- Kangas, V.-M.; Kvist, L.; Kholodova, M.; Nygrén, T.; Danilov, P.; Panchenko, D.; Fraimout, A.; Aspi, J. Evidence of post-glacial secondary contact and subsequent anthropogenic influence on the genetic composition of Fennoscandian moose (Alces alces). J. Biogeogr. 2015, 43, 2197–2208. [Google Scholar] [CrossRef]
- Kirkinen, T. Between Skins: Animal Skins in the Iron Age and Historical Burials in Eastern Fennoscandia. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2019. Available online: https://helda.helsinki.fi/items/597bb860-0ed6-4151-9dc6-7b1da79788b5 (accessed on 26 January 2024).
- Kirkinen, T. The role of wild animals in death rituals: Furs and animal skins in the late Iron Age inhumation burials in Southeastern Fennoscandia. Fennosc. Archaeol. 2015, 32, 101–120. Available online: https://journal.fi/fennoscandiaarchaeologica/article/view/126516 (accessed on 26 January 2024).
- Holmquist Olausson, L. ‘Älgmannen’ från Birka. Presentation av en nyligen undersökt krigargrav med människooffer. Fornvännen 1990, 85, 175–182. [Google Scholar]
- Eriksen, M.H. Doors to the dead. The power of doorways and thresholds in Viking Age Scandinavia. Archaeol. Dialogues 2013, 20, 187–214. [Google Scholar] [CrossRef]
- Shepherd, D.J. Bear, elk, and fish symbolism in Finnish contexts. In The Symbolic Role of Animals in Archaeology; Ryan, K., Crabtree, P.J., Eds.; University of Pennsylvania Press: Philadelphia, PA, USA, 1995; pp. 27–38. [Google Scholar]
- Bird-David, N. Us, Relatives: Scaling and Plural Life in a Forager World; Ethnographic Studies in Subjectivity, 12; University of California Press: Oakland, CA, USA, 2017. [Google Scholar]
- Heyd, V.M. Das Zeitalter der Ideologien: Migration, Interaktion & Expansion im prähistorischen Europa des 4. und 3. Jahrtausend v. Chr. In Transitional Landscapes? The 3rd Millennium BC in Europe; Furholt, M., Großmann, R., Szmyt, M., Eds.; Habelt: Bonn, Germany, 2016; pp. 53–85. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Elk | Bear | Beaver | Elk | Bear | Beaver | |||
|---|---|---|---|---|---|---|---|---|
| Period | Sites (n) | CON (%) | NISP 1 | NISP (%) | ||||
| Mesolithic age | 108 | 43 | 17 | 33 | 19,345 | 44.6 | 0.9 | 4.2 |
| Neolithic age | 103 | 46 | 17 | 42 | 29,840 | 10.4 | 0.2 | 21.0 |
| Bronze Age | 25 | 52 | 4 | 20 | 1459 | 29.9 | 0.1 | 4.3 |
| Iron Age | 49 | 6 | 31 | 18 | 4225 | 0.1 | 0.9 | 2.4 |
| Roman Period | 71 | 1 | 82 | 1 | 1001 | 0.1 | 5.8 | 0.2 |
| Early medieval period | 50 | 12 | 12 | 6 | 1208 | 3.0 | 0.5 | 0.2 |
| Late medieval period | 59 | 31 | 12 | 12 | 2787 | 8.7 | 0.4 | 0.9 |
| 465 | 59,865 | |||||||
| Elk | Bear | Beaver | Elk | Bear | Beaver | |||
|---|---|---|---|---|---|---|---|---|
| Period | Sites (n) | CON (%) | NISP 1 | NISP (%) | ||||
| Mesolithic age | 10 | 60 | 40 | 40 | 2041 | 48.7 | 2.2 | 33.0 |
| Neolithic age | 118 | 44 | 31 | 43 | 75,115 | 33.9 | 2.1 | 17.6 |
| Bronze Age | 87 | 14 | 18 | 25 | 13,089 | 27.3 | 2.3 | 8.3 |
| Iron Age | 64 | 25 | 16 | 44 | 7196 | 11.6 | 1.6 | 15.2 |
| Roman Period | 41 | 22 | 10 | 42 | 3569 | 1.8 | 4.0 | 7.6 |
| Early medieval period | 89 | 43 | 45 | 51 | 11,381 | 12.4 | 1.7 | 14.5 |
| Late medieval period | 150 | 42 | 39 | 48 | 40,371 | 10.7 | 1.4 | 4.6 |
| Early Modern period | 19 | 21 | 21 | 21 | 1521 | 18.6 | 2.5 | 2.1 |
| 578 | 154,283 | |||||||
| Elk | Bear | Beaver | Elk | Bear | Beaver | |||
|---|---|---|---|---|---|---|---|---|
| Period | Sites (n) | CON (%) | NISP 1 | NISP (%) | ||||
| Mesolithic age | 159 | 27 | 18 | 28 | 24,408 | 2.1 | 0.2 | 3.2 |
| Neolithic age | 409 | 6 | 12 | 17 | 56,900 | 1.5 | 1.0 | 6.3 |
| Bronze Age | 90 | 4 | 18 | 16 | 12,070 | 0.2 | 0.8 | 1.2 |
| Iron Age | 135 | 6 | 14 | 17 | 10,388 | 1.4 | 1.6 | 2.0 |
| Roman Period | 261 | 8 | 17 | 20 | 18,933 | 0.7 | 1.6 | 2.0 |
| Early medieval period | 252 | 8 | 9 | 12 | 9655 | 3.4 | 0.8 | 1.5 |
| Late medieval period | 205 | 9 | 21 | 17 | 28,291 | 0.4 | 1.0 | 0.8 |
| 1511 | 160,645 | |||||||
| Sites (n) | Others | Common Buzzard | Golden Eagle | Gyrfalcon | Peregrine Falcon | Common Kestrel | Sparrow-Hawk | Goshawk | White-Tailed Eagle | |
|---|---|---|---|---|---|---|---|---|---|---|
| Mesolithic age | 10 | 10.0 | 10.0 | 20.0 | 20.0 | |||||
| Neolithic age | 118 | 3.4 | 1.7 | 4.2 | ||||||
| Bronze Age | 87 | 1.1 | 1.1 | 1.1 | 2.3 | 1.1 | 3.4 | |||
| Iron Age | 64 | 3.1 | 1.6 | 3.1 | ||||||
| Roman Period | 41 | 2.4 | 2.4 | 2.4 | 2.4 | |||||
| Early medieval period | 89 | 1.1 | 1.1 | 1.1 | 1.1 | 6.7 | 3.4 | |||
| Late medieval period | 150 | 3.3 | 3.3 | 1.3 | 0.7 | 0.7 | 2.7 | 3.3 | 14.0 | 8.7 |
| 559 |
| Sites (n) | Others | Common Buzzard | Golden Eagle | Red Kite | Peregrine Falcon | Common Kestrel | Sparrow-Hawk | Goshawk | White-Tailed Eagle | |
|---|---|---|---|---|---|---|---|---|---|---|
| Mesolithic age | 159 | 7.5 | 4.4 | 1.9 | 0.6 | 0.6 | 0.6 | 23.3 | ||
| Neolithic age | 409 | 1.2 | 1.0 | 0.8 | 0.3 | 0.3 | 0.5 | 1.2 | 3.4 | |
| Bronze Age | 90 | 2.2 | 1.1 | 1.1 | 2.2 | 1.1 | ||||
| Iron Age | 135 | 1.5 | 1.5 | 3.0 | 2.2 | 1.5 | 4.4 | 3.0 | ||
| Roman Period | 261 | 2.3 | 1.5 | 0.4 | 0.8 | 0.4 | 0.8 | 3.5 | 5.4 | |
| Early medieval period | 252 | 3.6 | 2.0 | 1.2 | 1.2 | 1.2 | 0.8 | 2.4 | 4.4 | 5.6 |
| Late medieval period | 205 | 4.9 | 3.4 | 1.0 | 2.4 | 1.5 | 1.5 | 4.9 | 10.7 | 8.8 |
| 1511 |
| Sites (n) | Others | Common Buzzard | Golden Eagle | Gyrfalcon | Peregrine Falcon | Common Kestrel | Sparrow-Hawk | Goshawk | White-Tailed Eagle | |
|---|---|---|---|---|---|---|---|---|---|---|
| Mesolithic age | 108 | 0.9 | 1.8 | 2.7 | 0.9 | 09 | 0.9 | 2.7 | 4.6 | |
| Neolithic age | 103 | 1.0 | 1.0 | 1.0 | 2.9 | |||||
| Bronze Age | 25 | 4.0 | 4.0 | |||||||
| Iron Age | 49 | 2.0 | ||||||||
| Roman Period | 71 | 7.0 | 1.4 | 2.8 | 2.8 | 4.2 | 4.2 | |||
| Early medieval period | 50 | 12.0 | 2.0 | 6.0 | 14.0 | 2.0 | 10.0 | 72.0 | 6.0 | |
| Late medieval period | 59 | 16.9 | 3.4 | 3.4 | 5.1 | 3.4 | 6.8 | 25.4 | 27.1 | |
| 465 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmölcke, U.; Grimm, O. A Special Relationship—Aspects of Human–Animal Interaction in Birds of Prey, Brown Bears, Beavers, and Elk in Prehistoric Europe. Animals 2024, 14, 417. https://doi.org/10.3390/ani14030417
Schmölcke U, Grimm O. A Special Relationship—Aspects of Human–Animal Interaction in Birds of Prey, Brown Bears, Beavers, and Elk in Prehistoric Europe. Animals. 2024; 14(3):417. https://doi.org/10.3390/ani14030417
Chicago/Turabian StyleSchmölcke, Ulrich, and Oliver Grimm. 2024. "A Special Relationship—Aspects of Human–Animal Interaction in Birds of Prey, Brown Bears, Beavers, and Elk in Prehistoric Europe" Animals 14, no. 3: 417. https://doi.org/10.3390/ani14030417
APA StyleSchmölcke, U., & Grimm, O. (2024). A Special Relationship—Aspects of Human–Animal Interaction in Birds of Prey, Brown Bears, Beavers, and Elk in Prehistoric Europe. Animals, 14(3), 417. https://doi.org/10.3390/ani14030417