

Effect of Cassava Bioethanol By-Products and Crude Palm Oil Feeding on Fatty Acid Composition of Beef Meat and Fat in Crossbred Thai Indigenous Heifers

 ,
,  and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal and Diet Background and Meat Samples
2.2. Analysis of Fatty Acid Profiles
2.3. Analysis of Healthy Indices of Lipids
2.3.1. Atherogenicity Index (AI)
2.3.2. Thrombogenicity Index (TI)
2.3.3. Hypocholesterolemic/Hypercholesterolemic Ratio (h/H)
2.3.4. Nutritive Value Index (NVI)
2.3.5. Health-Promoting Index (HPI)
2.3.6. Peroxidizability Index (PI)
2.3.7. Enzyme Activity Index
2.4. Statistical Analysis
3. Results
3.1. Fatty Acid Profiles
3.2. Healthy Indices of Lipids
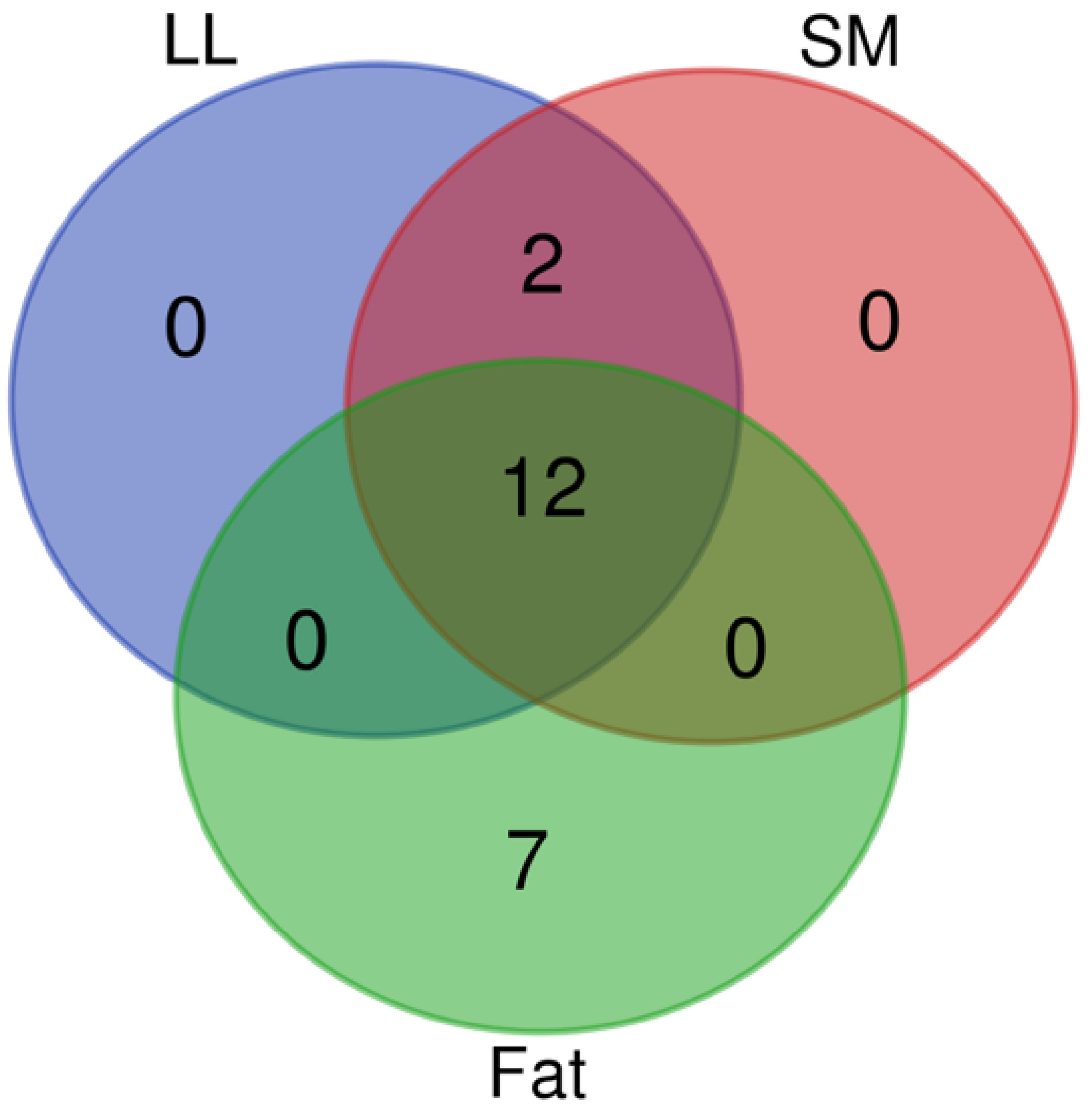
3.3. Multivariate Analysis Using Chemometric Approach
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hocquette, J.F.; Botreau, R.; Picard, B.; Jacquet, A.; Pethick, D.W.; Scollan, N.D. Opportunities for predicting and manipulating beef quality. Meat Sci. 2012, 92, 197–209. [Google Scholar] [CrossRef]
- Ponnampalam, E.; Priyashantha, H.; Vidanarachchi, J.; Kiani, A.; Holman, B. Effects of nutritional factors on fat content, fatty acid composition, and sensorial properties of meat and milk from domesticated ruminants: An overview. Animals 2024, 14, 840. [Google Scholar] [CrossRef] [PubMed]
- Aldai, N.; Dugan, M.E.R.; Rolland, D.C.; Kramer, J.K.G. Survey of the fatty acid composition of canadian beef: Backfat and longissimus lumborum muscle. Can. J. Anim. Sci. 2009, 89, 315–329. [Google Scholar] [CrossRef]
- Arshad, M.S.; Sohaib, M.; Ahmad, R.S.; Nadeem, M.T.; Imran, A.; Arshad, M.U.; Kwon, J.-H.; Amjad, Z. Ruminant meat flavor influenced by different factors with special reference to fatty acids. Lipids Health Dis. 2018, 17, 223. [Google Scholar] [CrossRef] [PubMed]
- Scollan, N.D.; Price, E.M.; Morgan, S.A.; Huws, S.A.; Shingfield, K.J. Can we improve the nutritional quality of meat? Proc. Nutr. Soc. 2017, 76, 603–618. [Google Scholar] [CrossRef]
- Scollan, N.; Hocquette, J.-F.F.; Nuernberg, K.; Dannenberger, D.; Richardson, I.; Moloney, A. Innovations in beef production systems that enhance the nutritional and health value of beef lipids and their relationship with meat quality. Meat Sci. 2006, 74, 17–33. [Google Scholar] [CrossRef]
- Scollan, N.D.; Dannenberger, D.; Nuernberg, K.; Richardson, I.; MacKintosh, S.; Hocquette, J.-F.F.; Moloney, A.P. Enhancing the nutritional and health value of beef lipids and their relationship with meat quality. Meat Sci. 2014, 97, 384–394. [Google Scholar] [CrossRef]
- Vahmani, P.; Mapiye, C.; Prieto, N.; Rolland, D.C.; McAllister, T.A.; Aalhus, J.L.; Dugan, M.E.R. The scope for manipulating the polyunsaturated fatty acid content of beef: A review. J. Anim. Sci. Biotechnol. 2015, 6, 29. [Google Scholar] [CrossRef]
- Realini, C.E.; Duckett, S.K.; Brito, G.W.; Dalla Rizza, M.; De Mattos, D. Effect of Pasture vs. concentrate feeding with or without antioxidants on carcass characteristics, fatty acid composition, and quality of Uruguayan beef. Meat Sci. 2004, 66, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Dannenberger, D.; Nuernberg, K.; Nuernberg, G.; Scollan, N.; Steinhart, H.; Ender, K. Effect of pasture vs. concentrate diet on CLA isomer distribution in different tissue lipids of beef cattle. Lipids 2005, 40, 589–598. [Google Scholar] [CrossRef]
- Hall, N.; Schönfeldt, H.; Pretorius, B. Fatty acids in beef from grain- and grass-fed cattle: The unique South African scenario. S. Afr. J. Clin. Nutr. 2016, 29, 55–62. [Google Scholar] [CrossRef]
- Fiorentini, G.; Lage, J.F.; Carvalho, I.P.C.; Messana, J.D.; Canesin, R.C.; Reis, R.A.; Berchielli, T.T. Lipid sources with different fatty acid profile alters the fatty acid profile and quality of beef from confined Nellore steers. Asian-Australas. J. Anim. Sci. 2015, 28, 976–986. [Google Scholar] [CrossRef] [PubMed]
- Mancini, A.; Imperlini, E.; Nigro, E.; Montagnese, C.; Daniele, A.; Orrù, S.; Buono, P. Biological and nutritional properties of palm oil and palmitic acid: Effects on health. Molecules 2015, 20, 17339–17361. [Google Scholar] [CrossRef] [PubMed]
- Lai, O.-M.; Phuah, E.-T.; Lee, Y.-Y.; Basiron, Y. Palm Oil. In Bailey’s Industrial Oil and Fat Products, 7th ed.; Shahidi, F., Ed.; John Wiley & Sons Inc: Hoboken, NJ, USA, 2020; pp. 385–485. [Google Scholar]
- Ogbonna, C.; Okoli, E. Evaluation of the potentials of some cassava varieties in nigeria for bio-ethanol production. Bio-Res. 2011, 8, 674–678. [Google Scholar] [CrossRef]
- Sriroth, K.; Piyachomkwan, K.; Wanlapatit, S.; Nivitchanyong, S. The promise of a technology revolution in cassava bioethanol: From thai practice to the world practice. Fuel 2010, 89, 1333–1338. [Google Scholar] [CrossRef]
- Phoemchalard, C.; Uriyapongson, S.; Berg, E.P. Effect of cassava bioethanol by-product and crude palm oil in Brahman x Thai native yearling heifer cattle diets: I. Nutrient digestibility and growth performance. Trop. Anim. Health Prod. 2014, 46, 663–668. [Google Scholar] [CrossRef]
- Phoemchalard, C.; Uriyapongson, S. Effect of cassava bioethanol by-product and crude palm oil in Brahman x Thai native yearling heifer cattle diets: II. Carcass characteristics and meat quality. Trop. Anim. Health Prod. 2015, 47, 1629–1631. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloanee Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Metcalfe, L.D.; Schmitz, A.A.; Pelka, J.R. Rapid preparation of fatty acid esters from lipids for gas chromatographic analysis. Anal. Chem. 1966, 38, 514–515. [Google Scholar] [CrossRef]
- Ostrowska, E.; Dunshea, F.R.; Muralitharan, M.; Cross, R.F. Comparison of silver-ion high-performance liquid chromatographic quantification of free and methylated conjugated linoleic acids. Lipids 2000, 35, 1147–1153. [Google Scholar] [CrossRef]
- Ulbricht, T.L.; Southgate, D.A. Coronary heart disease: Seven dietary factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Abrami, G.; Natiello, F.; Bronzi, P.; McKenzie, D.; Bolis, L.; Agradi, E. A comparison of highly unsaturated fatty acid levels in wild and farmed eels (Anguilla Anguilla). Comp. Biochem. Physiol. Part B Comp. Biochem. 1992, 101, 79–81. [Google Scholar] [CrossRef]
- Fernandes, C.E.; Vasconcelos, M.A.d.S.; de Almeida Ribeiro, M.; Sarubbo, L.A.; Andrade, S.A.C.; Filho, A.B.d.M. Nutritional and lipid profiles in marine fish species from Brazil. Food Chem. 2014, 160, 67–71. [Google Scholar] [CrossRef]
- Chen, Y.; Qiao, Y.; Xiao, Y.; Chen, H.; Zhao, L.; Huang, M.; Zhou, G. Differences in physicochemical and nutritional properties of breast and thigh meat from crossbred chickens, commercial broilers, and spent hens. Asian-Australas. J. Anim. Sci. 2015, 29, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Bobe, G.; Zimmerman, S.; Hammond, E.G.; Luhman, C.M.; Boylston, T.D.; Freeman, A.E.; Beitz, D.C. Physical and sensory properties of dairy products from cows with various milk fatty acid compositions. J. Agric. Food Chem. 2004, 52, 3422–3428. [Google Scholar] [CrossRef] [PubMed]
- Wołoszyn, J.; Haraf, G.; Okruszek, A.; Wereńska, M.; Goluch, Z.; Teleszko, M. Fatty acid profiles and health lipid indices in the breast muscles of local Polish Goose varieties. Poult. Sci. 2020, 99, 1216–1224. [Google Scholar] [CrossRef] [PubMed]
- Malau-Aduli, A.E.O.; Siebert, B.D.; Bottema, C.D.K.; Pitchford, W.S. A Comparison of the fatty acid composition of triacylglycerols in adipose tissue from Limousin and Jersey cattle. Aust. J. Agric. Res. 1997, 48, 715. [Google Scholar] [CrossRef]
- SAS. SAS/STAT® 14.1 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2015. [Google Scholar]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for comprehensive and integrative metabolomics data analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]
- Phoemchalard, C.; Tathong, T. Chemometric approach to characterizing and comparing the quality of buffalo meat from Nakhon Phanom and Khammouane Provinces. Int. J. Agric. Technol. 2023, 19, 2589–2604. [Google Scholar]
- Phoemchalard, C.; Uriyapongson, S.; Tathong, T.; Pornanek, P. 1H NMR metabolic profiling and meat quality in three beef cattle breeds from northeastern Thailand. Foods 2022, 11, 3821. [Google Scholar] [CrossRef]
- Biesalski, H.-K. Meat as a component of a healthy diet—Are There any risks or benefits if meat is avoided in the diet? Meat Sci. 2005, 70, 509–524. [Google Scholar] [CrossRef]
- Conte, G.; Serra, A.; Casarosa, L.; Ciucci, F.; Cappucci, A.; Bulleri, E.; Corrales-Retana, L.; Buccioni, A.; Mele, M. Effect of linseed supplementation on total longissimus muscle lipid composition and shelf-life of beef from young Maremmana bulls. Front. Vet. Sci. 2019, 5, 1–15. [Google Scholar] [CrossRef]
- Nuernberg, K. 14-Optimising the Nutritional Profile of Beef. In Improving the Sensory and Nutritional Quality of Fresh Meat; Kerry, J.P., Ledward, D., Eds.; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing: Cambridge, UK, 2009; pp. 321–341. ISBN 978-1-84569-343-5. [Google Scholar]
- Tomkins, T.; Drackley, J.K. Applications of palm oil in animal nutrition. J. Oil Palm Res. 2010, 22, 835–845. [Google Scholar]
- Doreau, M.; Ferlay, A. Digestion and utilisation of fatty acids by ruminants. Anim. Feed Sci. Technol. 1994, 45, 379–396. [Google Scholar] [CrossRef]
- Partida, J.A.; Olleta, J.L.; Sañudo, C.; Albertí, P.; Campo, M.M. Fatty acid composition and sensory traits of beef fed palm oil supplements. Meat Sci. 2007, 76, 444–454. [Google Scholar] [CrossRef]
- Guerrero, A.; Muela, E.; Valero, M.V.; Prado, I.N.; Campo, M.M.; Olleta, J.L.; Catalán, O.; Sañudo, C. Effect of the type of dietary fat when added as an energy source on animal performance, carcass characteristics and meat quality of intensively reared Friesian steers. Anim. Prod. Sci. 2016, 56, 1144. [Google Scholar] [CrossRef]
- Duckett, S.K.; Neel, J.P.S.; Fontenot, J.P.; Clapham, W.M. Effects of winter stocker growth rate and finishing system on: III. Tissue proximate, fatty acid, vitamin, and cholesterol content. J. Anim. Sci. 2009, 87, 2961–2970. [Google Scholar] [CrossRef]
- FAO. Fats and Fatty Acids in Human Nutrition. Rep. Expert Consult. 2008, 10, 14. [Google Scholar]
- Shramko, V.S.; Polonskaya, Y.V.; Kashtanova, E.V.; Stakhneva, E.M.; Ragino, Y.I. The short overview on the relevance of fatty acids for human cardiovascular disorders. Biomolecules 2020, 10, 1127. [Google Scholar] [CrossRef]
- dos Santos, N.J.A.; Bezerra, L.R.; Castro, D.P.V.; Marcelino, P.D.R.; Virgínio Júnior, G.F.; da Silva Júnior, J.M.; Pereira, E.S.; de Andrade, E.A.; Silva, T.M.; Barbosa, A.M.; et al. Effect of dietary palm kernel oil on the quality, fatty acid profile, and sensorial attributes of young bull meat. Foods 2022, 11, 609. [Google Scholar] [CrossRef] [PubMed]
- Noosen, P.; Lounglawan, P.; Suksombat, W. Linseed Oil supplemented concentrate fed to Brahman crossbred fattening steers on carcass quality traits and intramuscular fatty acid profiles. Songklanakarin J. Sci. Technol. 2017, 39, 1–10. [Google Scholar] [CrossRef]
- Park, S.; Yan, Z.; Choi, C.; Kim, K.; Lee, H.; Oh, Y.; Jeong, J.; Lee, J.; Smith, S.B.; Choi, S. Carcass and meat characteristics and gene expression in intramuscular adipose tissue of korean native cattle fed finishing diets supplemented with 5% palm oil. Korean J. Food Sci. Anim. Resour. 2017, 37, 168–174. [Google Scholar] [CrossRef]
- Daley, C.A.; Abbott, A.; Doyle, P.S.; Nader, G.A.; Larson, S. A review of fatty acid profiles and antioxidant content in grass-fed and grain-fed beef. Nutr. J. 2010, 9, 10. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M. Monounsaturated fatty acids and risk of cardiovascular disease. Circulation 1999, 100, 1253–1258. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Hoffmann, G. Monounsaturated fatty acids and risk of cardiovascular disease: Synopsis of the evidence available from systematic reviews and meta-analyses. Nutrients 2012, 4, 1989–2007. [Google Scholar] [CrossRef]
- Enser, M.; Hallett, K.; Hewitt, B.; Fursey, G.A.J.; Wood, J.D. Fatty acid content and composition of English beef, lamb and pork at retail. Meat Sci. 1996, 42, 443–456. [Google Scholar] [CrossRef]
- Garcia, P.T.; Pensel, N.A.; Sancho, A.M.; Latimori, N.J.; Kloster, A.M.; Amigone, M.A.; Casal, J.J. Beef lipids in relation to animal breed and nutrition in Argentina. Meat Sci. 2008, 79, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Alfaia, C.P.M.; Alves, S.P.; Martins, S.I.V.; Costa, A.S.H.; Fontes, C.M.G.A.; Lemos, J.P.C.; Bessa, R.J.B.; Prates, J.A.M. Effect of the feeding system on intramuscular fatty acids and conjugated linoleic acid isomers of beef cattle, with emphasis on their nutritional value and discriminatory ability. Food Chem. 2009, 114, 939–946. [Google Scholar] [CrossRef]
- Park, Y.; Pariza, M.W. Mechanisms of body fat modulation by conjugated linoleic acid (CLA). Food Res. Int. 2007, 40, 311–323. [Google Scholar] [CrossRef]
- Whigham, L.D.; Cook, M.E.; Atkinson, R.L. Conjugated linoleic acid: Implications for human health. Pharmacol. Res. 2000, 42, 503–510. [Google Scholar] [CrossRef]
- Nuernberg, K.; Dannenberger, D.; Nuernberg, G.; Ender, K.; Voigt, J.; Scollan, N.D.; Wood, J.D.; Nute, G.R.; Richardson, R.I. Effect of a grass-based and a concentrate feeding system on meat quality characteristics and fatty acid composition of longissimus muscle in different cattle breeds. Livest. Prod. Sci. 2005, 94, 137–147. [Google Scholar] [CrossRef]
- Kratz, M. Dietary Cholesterol, Atherosclerosis and Coronary Heart Disease. In Atherosclerosis: Diet and Drugs; von Eckardstein, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 195–213. ISBN 978-3-540-27661-6. [Google Scholar]
- Djuricic, I.; Calder, P.C. Beneficial outcomes of omega-6 and omega-3 polyunsaturated fatty acids on human health: An update for 2021. Nutrients 2021, 13, 2421. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. The omega-6/omega-3 fatty acid ratio: Health implications. OCL-Oléagineux Corps Gras Lipides 2010, 17, 267–275. [Google Scholar] [CrossRef]
- Patterson, E.; Wall, R.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. Health implications of high dietary omega-6 polyunsaturated fatty acids. J. Nutr. Metab. 2012, 2012. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Evolutionary aspects of diet, the omega-6/omega-3 ratio and genetic variation: Nutritional implications for chronic diseases. Biomed. Pharmacother. 2006, 60, 502–507. [Google Scholar] [CrossRef]
- Verbeke, W. Functional foods: Consumer willingness to compromise on taste for health? Food Qual. Prefer. 2006, 17, 126–131. [Google Scholar] [CrossRef]
- Department of Health and Social Security Diet and Cardiovascular Disease; Report on Health and Social Subjects; Her Majesty’s Stationery Office (HMSO): London, UK, 1984; Volume 28.
- Williams, C.M. Dietary Fatty Acids and Human Health. Ann. Zootech. 2000, 49, 165–180. [Google Scholar] [CrossRef]
- Chen, J.; Li, J.; Liu, X.; He, Y. Effects of dietary fat saturation level on growth performance, carcass traits, blood lipid parameters, tissue fatty acid composition and meat quality of finishing pigs. Anim. Biosci. 2021, 34, 895–903. [Google Scholar] [CrossRef]
- Smith, S.B. Oleic acid concentration in bovine adipose tissues: Impact on human health, sensory attributes, and genetic regulation. Front. Anim. Sci. 2024, 5, 1332861. [Google Scholar] [CrossRef]
- Chen, J.; Liu, H. Nutritional indices for assessing fatty acids: A mini-review. Int. J. Mol. Sci. 2020, 21, 5695. [Google Scholar] [CrossRef] [PubMed]
- Dev, K.; Begum, J.; Biswas, A.; Mir, N.A.; Singh, J.; Prakash, R.; Sonowal, J.; Bharali, K.; Tomar, S.; Kant, R.; et al. Hepatic transcriptome analysis reveals altered lipid metabolism and consequent health indices in chicken supplemented with dietary bifidobacterium bifidum and mannan-oligosaccharides. Sci. Rep. 2021, 11, 17895. [Google Scholar] [CrossRef] [PubMed]
- Pilarczyk, R.; Wójcik, J.; Sablik, P.; Czerniak, P. fatty acid profile and health lipid indices in the raw milk of Simmental and Holstein-Friesian cows from an organic farm. S. Afr. J. Anim. Sci. 2015, 45, 30. [Google Scholar] [CrossRef]
- Dal Bosco, A.; Cavallo, M.; Menchetti, L.; Angelucci, E.; Cartoni Mancinelli, A.; Vaudo, G.; Marconi, S.; Camilli, E.; Galli, F.; Castellini, C.; et al. The healthy fatty index allows for deeper insights into the lipid composition of foods of animal origin when compared with the atherogenic and thrombogenicity indexes. Foods 2024, 13, 1568. [Google Scholar] [CrossRef]
- Santos-Silva, J.; Bessa, R.J.; Santos-Silva, F. Effect of genotype, feeding system and slaughter weight on the quality of light lambs. Livest. Prod. Sci. 2002, 77, 187–194. [Google Scholar] [CrossRef]
- Mapiye, C.; Chimonyo, M.; Dzama, K.; Hugo, A.; Strydom, P.E.; Muchenje, V. Fatty acid composition of beef from Nguni steers supplemented with Acacia Karroo leaf-meal. J. Food Compos. Anal. 2011, 24, 523–528. [Google Scholar] [CrossRef]
- Frunză, G.; Murariu, O.C.; Ciobanu, M.-M.; Radu-Rusu, R.-M.; Simeanu, D.; Boișteanu, P.-C. Meat quality in rabbit (Oryctolagus Cuniculus) and hare (Lepus Europaeus Pallas)—A nutritional and technological perspective. Agriculture 2023, 13, 126. [Google Scholar] [CrossRef]
- Dal Bosco, A.; Cartoni Mancinelli, A.; Vaudo, G.; Cavallo, M.; Castellini, C.; Mattioli, S. Indexing of Fatty acids in poultry meat for its characterization in healthy human nutrition: A comprehensive application of the scientific literature and new proposals. Nutrients 2022, 14, 3110. [Google Scholar] [CrossRef] [PubMed]
- Rezamand, P.; Watts, J.S.; Yavah, K.M.; Mosley, E.E.; Ma, L.; Corl, B.A.; McGuire, M.A. Relationship between Stearoyl-CoA desaturase 1 gene expression, relative protein abundance, and its fatty acid products in bovine tissues. J. Dairy Res. 2014, 81, 333–339. [Google Scholar] [CrossRef]
- Domínguez-López, I.; Arancibia-Riveros, C.; Tresserra-Rimbau, A.; Castro-Barquero, S.; Casas, R.; Vázquez-Ruiz, Z.; Ros, E.; Fitó, M.; Estruch, R.; López-Sabater, M.C.; et al. Relationship between estimated desaturase enzyme activity and metabolic syndrome in a longitudinal study. Front. Nutr. 2022, 9, 1–9. [Google Scholar] [CrossRef]
- Murakami, K.; Sasaki, S.; Takahashi, Y.; Uenishi, K.; Watanabe, T.; Kohri, T.; Yamasaki, M.; Watanabe, R.; Baba, K.; Shibata, K.; et al. Lower estimates of δ-5 desaturase and elongase activity are related to adverse profiles for several metabolic risk factors in young Japanese women. Nutr. Res. 2008, 28, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Hyun Joo, D.; Chung, H.K.; Moon, J.; Shin, M.-J. Relationship between the estimates of desaturase activities and cardiometabolic phenotypes in koreans. J. Clin. Biochem. Nutr. 2011, 49, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Bonafini, S.; Giontella, A.; Tagetti, A.; Bresadola, I.; Gaudino, R.; Cavarzere, P.; Ramaroli, D.A.; Branz, L.; Marcon, D.; Pietrobelli, A.; et al. Fatty acid profile and desaturase activities in 7–10-year-old children attending primary school in Verona South District: Association between palmitoleic acid, SCD-16, indices of adiposity, and blood pressure. Int. J. Mol. Sci. 2020, 21, 3899. [Google Scholar] [CrossRef] [PubMed]
- van Cleef, E.H.C.B.; D’Áurea, A.P.; Fávaro, V.R.; van Cleef, F.O.S.; Barducci, R.S.; Almeida, M.T.C.; Machado Neto, O.R.; Ezequiel, J.M.B. Effects of dietary inclusion of high concentrations of crude glycerin on meat quality and fatty acid profile of feedlot fed Nellore bulls. PLoS ONE 2017, 12, e0179830. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yu, H.; Gao, R.; Liu, M.; Xie, W. A Comprehensive review of the family of very-long-chain fatty acid elongases: Structure, function, and implications in physiology and pathology. Eur. J. Med. Res. 2023, 28, 532. [Google Scholar] [CrossRef]
- Tillander, V.; Alexson, S.E.H.; Cohen, D.E. Deactivating Fatty Acids: Acyl-CoA Thioesterase-mediated control of lipid metabolism. Trends Endocrinol. Metab. 2017, 28, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Caswell, B.T.; de Carvalho, C.C.; Nguyen, H.; Roy, M.; Nguyen, T.; Cantu, D.C. Thioesterase enzyme families: Functions, structures, and mechanisms. Protein Sci. 2022, 31, 652–676. [Google Scholar] [CrossRef]
- Xia, X.; Chen, C.; Yang, L.; Wang, Y.; Duan, A.; Wang, D. Analysis of metabolites in young and mature Docynia Delavayi (Franch.) Schneid leaves using UPLC-ESI-MS/MS. PeerJ 2022, 10, e12844. [Google Scholar] [CrossRef]

{kind=link}
| Fatty Acids | LCEP | HCEP | CPO | RS |
|---|---|---|---|---|
| C8:0 | 2.05 | 1.91 | ND | ND |
| C10:0 | 1.83 | 1.94 | ND | ND |
| C12:0 | 18.40 | 21.65 | ND | ND |
| C14:0 | 8.21 | 9.39 | 1.05 | 1.28 |
| C14:1 | 0.00 | 0.07 | ND | ND |
| C16:0 | 22.00 | 24.96 | 45.31 | 47.49 |
| C16:1 | 8.52 | 6.37 | ND | ND |
| C18:0 | 3.20 | 3.63 | 3.95 | 8.57 |
| C18:1n9t | 0.00 | 0.30 | 37.72 | ND |
| C18:1n9c | 23.73 | 22.54 | ND | 16.76 |
| C18:2n6c | 11.17 | 6.55 | 10.75 | 19.88 |
| C18:3n3 | 0.73 | 0.58 | ND | 6.03 |
| C20:3n6 | 0.15 | 0.12 | ND | ND |
| Total | 100.00 | 100.00 | 100.00 | 100.00 |
| Fatty Acid | Cassava Bioethanol By-Product (CEP) | p-Value | Crude Palm Oil (CPO) | p-Value | |||
|---|---|---|---|---|---|---|---|
| LCEP (15%) | HCEP (30%) | CPO-0 (0%) | CPO-2 (2%) | CPO-4 (4%) | |||
| C12:0 | 0.37 ± 0.11 | 0.37 ± 0.11 | NS | 0.48 ± 0.01 a | 0.39 ± 0.00 b | 0.23 ± 0.01 c | *** |
| C13:0 | 0.25 ± 0.17 | 0.25 ± 0.17 | NS | 0.04 ± 0.03 c | 0.26 ± 0.01 b | 0.44 ± 0.02 a | *** |
| C14:0 | 4.99 ± 1.14 | 4.99 ± 1.12 | NS | 6.11 ± 0.12 a | 5.31 ± 0.04 b | 3.56 ± 0.07 c | *** |
| C14:1 | 0.45 ± 0.11 | 0.46 ± 0.10 | NS | 0.55 ± 0.06 a | 0.49 ± 0.03 a | 0.33 ± 0.00 b | *** |
| C16:0 | 28.07 ± 2.92 | 28.09 ± 2.86 | NS | 30.69 ± 0.27 a | 29.22 ± 0.11 b | 24.34 ± 0.42 c | *** |
| C16:1 | 14.41 ± 5.29 | 14.34 ± 4.88 | NS | 10.49 ± 0.92 b | 11.71 ± 0.20 b | 20.91 ± 2.05 a | *** |
| C18:0 | 16.47 ± 1.99 | 16.40 ± 1.88 | NS | 18.29 ± 0.71 a | 16.99 ± 0.38 b | 14.03 ± 0.09 c | *** |
| C18:1n9t | 0.96 ± 0.54 | 0.95 ± 0.47 | NS | 1.39 ± 0.11 a | 1.10 ± 0.40 a | 0.37 ± 0.02 b | *** |
| C18:1n9c | 29.03 ± 1.66 | 29.16 ± 1.08 | NS | 28.67 ± 0.10 b | 30.28 ± 0.80 a | 28.32 ± 1.74 b | * |
| C18:2n6c | 2.32 ± 0.82 | 2.32 ± 0.80 | NS | 1.60 ± 0.03 c | 1.98 ± 0.04 b | 3.38 ± 0.15 a | *** |
| C18:3n3 | 0.17 ± 0.09 | 0.18 ± 0.05 | NS | 0.15 ± 0.04 | 0.23 ± 0.00 | 0.16 ± 0.10 | NS |
| C20:3n6 | 0.30 ± 0.16 | 0.31 ± 0.16 | NS | 0.18 ± 0.00 c | 0.22 ± 0.02 b | 0.52 ± 0.01 a | *** |
| C20:3n3 | 1.49 ± 0.65 | 1.50 ± 0.64 | NS | 0.92 ± 0.04 c | 1.21 ± 0.03 b | 2.34 ± 0.04 a | *** |
| C22:6n3 | 0.65 ± 0.28 | 0.64 ± 0.27 | NS | 0.38 ± 0.03 c | 0.55 ± 0.00 b | 1.01 ± 0.06 a | *** |
| SFAs | 50.16 ± 5.95 | 50.11 ± 5.80 | NS | 55.62 ± 1.13 a | 52.18 ± 0.24 b | 42.61 ± 0.40 c | *** |
| MUFAs | 44.87 ± 4.06 | 44.91 ± 3.91 | NS | 41.12 ± 0.98 c | 43.60 ± 0.15 b | 49.95 ± 0.35 a | *** |
| PUFAs | 4.95 ± 1.90 | 4.96 ± 1.88 | NS | 3.25 ± 0.14 c | 4.21 ± 0.08 b | 7.42 ± 0.05 a | *** |
| Omega 3 | 2.32 ± 0.93 | 2.33 ± 0.92 | NS | 1.46 ± 0.11 c | 2.00 ± 0.01 b | 3.52 ± 0.08 a | *** |
| Omega 6 | 2.63 ± 0.98 | 2.63 ± 0.96 | NS | 1.78 ± 0.02 c | 2.20 ± 0.06 b | 3.90 ± 0.14 a | *** |
| Omega 6:3 | 1.15 ± 0.10 | 1.14 ± 0.06 | NS | 1.23 ± 0.08 a | 1.10 ± 0.02 b | 1.11 ± 0.06 b | * |
| PUFA:SFA | 0.10 ± 0.03 | 0.10 ± 0.03 | NS | 0.07 ± 0.00 c | 0.09 ± 0.00 b | 0.14 ± 0.00 a | *** |
| MUFA:SFA | 0.91 ± 0.2 | 0.91 ± 0.19 | NS | 0.74 ± 0.03 c | 0.83 ± 0.00 b | 1.17 ± 0.01 a | *** |
| UFA:SFA | 1.02 ± 0.25 | 1.02 ± 0.24 | NS | 0.79 ± 0.03 c | 0.91 ± 0.00 b | 1.34 ± 0.02 a | *** |
| HFA:SFA | 0.66 ± 0.00 | 0.66 ± 0.00 | NS | 0.67 ± 0.00 a | 0.66 ± 0.00 a | 0.65 ± 0.00 b | * |
| AI | 0.99 ± 0.26 | 0.99 ± 0.25 | NS | 1.25 ± 0.04 a | 1.06 ± 0.00 b | 0.67 ± 0.01 c | *** |
| TI | 1.63 ± 0.44 | 1.62 ± 0.42 | NS | 2.07 ± 0.11 a | 1.72 ± 0.01 b | 1.09 ± 0.00 c | *** |
| h:H | 1.07 ± 0.17 | 1.07 ± 0.17 | NS | 0.90 ± 0.01 c | 1.03 ± 0.00 b | 1.29 ± 0.03 a | *** |
| NVI | 1.66 ± 0.08 | 1.66 ± 0.08 | NS | 1.57 ± 0.00 c | 1.65 ± 0.00 b | 1.75 ± 0.03 a | *** |
| HPI | 1.07 ± 0.31 | 1.07 ± 0.30 | NS | 0.79 ± 0.03 c | 0.94 ± 0.00 b | 1.47 ± 0.03 a | *** |
| PI | 12.62 ± 4.81 | 12.63 ± 4.74 | NS | 8.24 ± 0.48 c | 10.82 ± 0.10 b | 18.8 ± 0.33 a | *** |
| SCD-16 | 33.38 ± 10.05 | 33.30 ± 9.50 | NS | 25.42 ± 1.84 b | 28.61 ± 0.44 b | 46.00 ± 2.87 a | *** |
| SCD-18 | 63.86 ± 2.85 | 64.04 ± 2.56 | NS | 61.08 ± 0.83 c | 64.04 ± 1.13 b | 66.74 ± 1.52 a | *** |
| EI | 51.70 ± 3.02 | 51.77 ± 2.54 | NS | 53.27 ± 0.82 a | 53.59 ± 0.27 a | 48.34 ± 1.87 b | *** |
| TE | 5.78 ± 0.81 | 5.78 ± 0.81 | NS | 5.02 ± 0.05 c | 5.50 ± 0.02 b | 6.83 ± 0.01 a | *** |
| Fatty Acid | Cassava Bioethanol By-Product (CEP) | p-Value | Crude Palm Oil (CPO) | p-Value | |||
|---|---|---|---|---|---|---|---|
| LCEP (15%) | HCEP (30%) | CPO-0 (0%) | CPO-2 (2%) | CPO-4 (4%) | |||
| C12:0 | 0.32 ± 0.08 | 0.32 ± 0.05 | NS | 0.33 ± 0.04 a | 0.25 ± 0.04 b | 0.38 ± 0.06 a | ** |
| C13:0 | 0.43 ± 0.07 | 0.43 ± 0.07 | NS | 0.46 ± 0.00 b | 0.50 ± 0.00 a | 0.33 ± 0.03 c | *** |
| C14:0 | 3.83 ± 0.60 | 3.82 ± 0.37 | NS | 4.11 ± 0.22 a | 3.37 ± 0.50 b | 3.99 ± 0.35 a | * |
| C14:1 | 0.41 ± 0.03 | 0.41 ± 0.02 | NS | 0.44 ± 0.01 a | 0.38 ± 0.01 b | 0.43 ± 0.03 a | ** |
| C16:0 | 24.55 ± 1.53 | 24.6 ± 0.95 | NS | 25.19 ± 0.05 a | 23.43 ± 1.61 b | 25.1 ± 0.49 a | * |
| C16:1 | 20.44 ± 3.11 | 20.25 ± 2.11 | NS | 18.23 ± 0.78 b | 22.66 ± 3.18 a | 20.13 ± 0.34 a,b | * |
| C18:0 | 13.55 ± 1.22 | 13.62 ± 0.77 | NS | 13.53 ± 0.00 a,b | 12.82 ± 1.36 b | 14.4 ± 0.01 a | * |
| C18:1n9t | 0.47 ± 0.08 | 0.47 ± 0.06 | NS | 0.44 ± 0.06 b | 0.42 ± 0.01 b | 0.55 ± 0.03 a | *** |
| C18:1n9c | 27.86 ± 1.26 | 27.94 ± 0.98 | NS | 28.73 ± 1.22 a | 28.20 ± 0.04 a | 26.77 ± 0.38 b | ** |
| C18:2n6c | 3.60 ± 0.31 | 3.57 ± 0.20 | NS | 3.73 ± 0.32 a | 3.36 ± 0.16 b | 3.66 ± 0.01 a | * |
| C18:3n3 | 0.23 ± 0.10 | 0.23 ± 0.08 | NS | 0.27 ± 0.01 a | 0.12 ± 0.08 b | 0.30 ± 0.00 a | *** |
| C20:3n6 | 0.54 ± 0.03 | 0.54 ± 0.01 | NS | 0.55 ± 0.00 | 0.53 ± 0.00 | 0.54 ± 0.04 | NS |
| C20:3n3 | 2.58 ± 0.35 | 2.59 ± 0.23 | NS | 2.70 ± 0.25 a | 2.73 ± 0.21 a | 2.31 ± 0.21 b | * |
| C22:6n3 | 1.14 ± 0.12 | 1.13 ± 0.09 | NS | 1.21 ± 0.01 a | 1.17 ± 0.11 a | 1.03 ± 0.03 b | ** |
| SFAs | 42.69 ± 3.29 | 42.81 ± 2.00 | NS | 43.65 ± 0.32 a | 40.39 ± 3.54 b | 44.23 ± 0.93 a | * |
| MUFAs | 49.19 ± 3.07 | 49.08 ± 2.02 | NS | 47.85 ± 0.40 b | 51.66 ± 3.10 a | 47.90 ± 0.65 b | * |
| PUFAs | 8.10 ± 0.47 | 8.09 ± 0.33 | NS | 8.49 ± 0.07 a | 7.93 ± 0.43 b | 7.86 ± 0.28 b | * |
| Omega 3 | 3.95 ± 0.40 | 3.97 ± 0.27 | NS | 4.19 ± 0.24 a | 4.04 ± 0.25 a | 3.65 ± 0.25 b | * |
| Omega 6 | 4.14 ± 0.32 | 4.12 ± 0.21 | NS | 4.29 ± 0.32 a | 3.89 ± 0.17 b | 4.21 ± 0.02 a | * |
| Omega 6:3 | 1.06 ± 0.14 | 1.05 ± 0.09 | NS | 1.03 ± 0.14 a,b | 0.96 ± 0.01 b | 1.16 ± 0.07 a | * |
| PUFA:SFA | 0.16 ± 0.01 | 0.16 ± 0.01 | NS | 0.17 ± 0.00 a | 0.15 ± 0.00 c | 0.16 ± 0.00 b | *** |
| MUFA:SFA | 1.16 ± 0.18 | 1.15 ± 0.10 | NS | 1.09 ± 0.01 b | 1.30 ± 0.19 a | 1.08 ± 0.03 b | * |
| UFA:SFA | 1.36 ± 0.21 | 1.34 ± 0.11 | NS | 1.29 ± 0.01 b | 1.50 ± 0.22 a | 1.26 ± 0.04 b | * |
| HFA:SFA | 0.67 ± 0.00 | 0.67 ± 0.00 | NS | 0.67 ± 0.00 a | 0.67 ± 0.00 a | 0.66 ± 0.00 b | ** |
| AI | 0.70 ± 0.10 | 0.70 ± 0.06 | NS | 0.74 ± 0.02 a | 0.63 ± 0.09 b | 0.74 ± 0.04 a | * |
| TI | 1.07 ± 0.13 | 1.06 ± 0.08 | NS | 1.08 ± 0.03 a,b | 0.97 ± 0.14 b | 1.15 ± 0.05 a | * |
| h:H | 1.29 ± 0.13 | 1.28 ± 0.07 | NS | 1.28 ± 0.04 a,b | 1.37 ± 0.12 b | 1.21 ± 0.05 a | * |
| NVI | 1.71 ± 0.08 | 1.71 ± 0.05 | NS | 1.69 ± 0.04 b | 1.77 ± 0.06 a | 1.66 ± 0.04 b | * |
| HPI | 1.44 ± 0.25 | 1.42 ± 0.14 | NS | 1.34 ± 0.03 b | 1.62 ± 0.26 a | 1.34 ± 0.08 b | * |
| PI | 20.69 ± 1.60 | 20.66 ± 1.11 | NS | 21.72 ± 0.27 | 20.85 ± 1.48 | 19.45 ± 0.79 | * |
| SCD-16 | 45.22 ± 5.00 | 44.94 ± 3.31 | NS | 41.94 ± 0.99 b | 48.79 ± 5.21 a | 44.52 ± 0.91 a,b | * |
| SCD-18 | 67.30 ± 2.55 | 67.25 ± 1.87 | NS | 67.91 ± 0.93 a | 68.90 ± 2.28 a | 65.00 ± 0.30 b | ** |
| EI | 47.93 ± 1.82 | 48.10 ± 1.20 | NS | 49.31 ± 1.20 a | 47.09 ± 1.70 b | 47.64 ± 0.32 a,b | * |
| TE | 6.51 ± 0.74 | 6.46 ± 0.42 | NS | 6.13 ± 0.31 b | 7.01 ± 0.61 a | 6.31 ± 0.43 a,b | * |
| Treatment | LCEP | HCEP | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CPO-0 | CPO-2 | CPO-4 | CPO-0 | CPO-2 | CPO-4 | CEP | CPO | Int. | |
| C10:0 | 0.10 ± 0.02 A,a | 0.06 ± 0.01 A,b | 0.06 ± 0.01 A,b | 0.07 ± 0.01 B,a | 0.05 ± 0.03 B,b | 0.05 ± 0.01 B,b | * | *** | NS |
| C12:0 | 1.00 ± 0.09 | 0.98 ± 0.29 | 1.05 ± 0.03 | 1.05 ± 0.04 | 1.07 ± 0.27 | 1.27 ± 0.35 | NS | NS | NS |
| C13:0 | 0.16 ± 0.03 a | 0.11 ± 0.04 b | 0.14 ± 0.03 b | 0.20 ± 0.03 a | 0.14 ± 0.02 b | 0.13 ± 0.01 b | NS | ** | NS |
| C14:0 | 10.54 ± 0.59 | 9.89 ± 0.65 | 9.95 ± 0.34 | 10.4 ± 0.21 | 10.18 ± 0.64 | 9.39 ± 0.53 | NS | NS | NS |
| C16:0 | 36.35 ± 0.16 a,b | 35.85 ± 1.57 b | 37.37 ± 0.42 a | 36.29 ± 0.13 a,b | 35.64 ± 0.55 b | 36.78 ± 0.47 a | NS | * | NS |
| C16:1 | 0.80 ± 0.77 y | 1.89 ± 0.04 x | 1.70 ± 0.15 x | 2.13 ± 0.03 x | 2.07 ± 0.08 x | 2.23 ± 0.18 x | *** | * | * |
| C18:0 | 22.02 ± 3.58 | 20.29 ± 0.90 | 21.84 ± 0.94 | 19.98 ± 6.33 | 19.44 ± 1.62 | 22.45 ± 5.19 | NS | NS | NS |
| C18:1n9c | 28.30 ± 1.68 | 30.15 ± 3.52 | 27.19 ± 0.04 | 28.84 ± 5.99 | 30.38 ± 3.08 | 26.82 ± 6.19 | NS | NS | NS |
| C18:2n6t | 0.16 ± 0.03 a | 0.10 ± 0.03 b | 0.07 ± 0.01 b | 0.16 ± 0.05 a | 0.10 ± 0.02 b | 0.10 ± 0.07 b | NS | * | NS |
| C18:2n6c | 0.24 ± 0.20 y | 0.43 ± 0.05 x | 0.41 ± 0.01 x | 0.52 ± 0.03 x | 0.50 ± 0.01 x | 0.42 ± 0.01 x | ** | NS | * |
| C9T11 | 0.11 ± 0.02 B | 0.09 ± 0.02 B | 0.11 ± 0.01 B | 0.14 ± 0.02 A | 0.13 ± 0.02 A | 0.11 ± 0.02 A | ** | NS | NS |
| C18:3n3 | 0.10 ± 0.07 B | 0.08 ± 0.07 B | 0.02 ± 0.01 B | 0.08 ± 0.06 A | 0.13 ± 0.03 A | 0.12 ± 0.04 A | * | NS | NS |
| C9C11 | 0.01 ± 0.00 B | 0.01 ± 0.00 B | 0.01 ± 0.00 B | 0.01 ± 0.01 A | 0.03 ± 0.02 A | 0.01 ± 0.01 A | * | NS | NS |
| T10C12 | 0.03 ± 0.01 | 0.02 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | NS | NS | NS |
| C21:0 | 0.04 ± 0.01 B | 0.05 ± 0.02 B | 0.05 ± 0.01 B | 0.06 ± 0.02 A | 0.08 ± 0.01 A | 0.07 ± 0.02 A | * | NS | NS |
| C20:2 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | NS | NS | NS |
| C20:3n6 | 0.02 ± 0.01 y,z | 0.04 ± 0.02 x | 0.01 ± 0.01 z | 0.02 ± 0.01 y,z | 0.03 ± 0.01 y | 0.02 ± 0.01 y,z | NS | ** | * |
| C20:3n3 | 0.04 ± 0.02 a | 0.03 ± 0.01 b | 0.02 ± 0.01 b | 0.04 ± 0.01 a | 0.03 ± 0.01 b | 0.03 ± 0.01 b | NS | * | NS |
| C22:6n3 | 0.02 ± 0.01 B,a | 0.00 ± 0.00 B,b | 0.03 ± 0.01 B,a | 0.03 ± 0.01 A,a | 0.02 ± 0.02 A,b | 0.04 ± 0.01 A,a | * | ** | NS |
| SFAs | 70.19 ± 2.77 | 67.2 ± 3.45 | 70.43 ± 0.14 | 68.03 ± 5.99 | 66.59 ± 3.06 | 70.12 ± 6.50 | NS | NS | NS |
| MUFAs | 29.10 ± 2.45 | 32.03 ± 3.56 | 28.89 ± 0.11 | 30.97 ± 6.02 | 32.44 ± 3.00 | 29.04 ± 6.37 | NS | NS | NS |
| PUFAs | 0.73 ± 0.32 B | 0.78 ± 0.12 B | 0.69 ± 0.03 B | 1.01 ± 0.04 A | 0.98 ± 0.06 A | 0.86 ± 0.14 A | *** | NS | NS |
| Omega 3 | 0.15 ± 0.09 B | 0.10 ± 0.07 B | 0.06 ± 0.01 B | 0.14 ± 0.06 A | 0.18 ± 0.05 A | 0.18 ± 0.03 A | * | NS | NS |
| Omega 6 | 0.59 ± 0.24 B | 0.69 ± 0.06 B | 0.64 ± 0.02 B | 0.88 ± 0.09 A | 0.81 ± 0.02 A | 0.68 ± 0.11 A | * | NS | NS |
| Omega 6:3 | 4.64 ± 1.01 y | 11.29 ± 6.60 x | 11.83 ± 0.44 x | 8.22 ± 4.22 x,y | 5.02 ± 1.21 y | 3.90 ± 0.04 y | * | NS | * |
| PUFA:SFA | 0.01 ± 0.01 B | 0.01 ± 0.01 B | 0.01 ± 0.01 B | 0.02 ± 0.01 A | 0.02 ± 0.01 A | 0.02 ± 0.01 A | * | NS | NS |
| MUFA:SFA | 0.42 ± 0.06 | 0.48 ± 0.08 | 0.41 ± 0.01 | 0.47 ± 0.13 | 0.49 ± 0.07 | 0.43 ± 0.14 | NS | NS | NS |
| UFA:SFA | 0.43 ± 0.06 | 0.49 ± 0.08 | 0.42 ± 0.01 | 0.48 ± 0.14 | 0.51 ± 0.07 | 0.44 ± 0.14 | NS | NS | NS |
| HFA:SFA | 0.68 ± 0.04 | 0.70 ± 0.01 | 0.69 ± 0.02 | 0.71 ± 0.07 | 0.70 ± 0.02 | 0.68 ± 0.05 | NS | NS | NS |
| AI | 2.68 ± 0.17 | 2.37 ± 0.39 | 2.64 ± 0.05 | 2.55 ± 0.45 | 2.34 ± 0.32 | 2.68 ± 0.68 | NS | NS | NS |
| TI | 4.52 ± 0.67 | 3.98 ± 0.54 | 4.61 ± 0.04 | 4.28 ± 1.21 | 3.81 ± 0.54 | 4.68 ± 1.39 | NS | NS | NS |
| h:H | 0.62 ± 0.04 | 0.58 ± 0.11 | 0.59 ± 0.01 | 0.64 ± 0.13 | 0.69 ± 0.09 | 0.60 ± 0.15 | NS | NS | NS |
| NVI | 1.38 ± 0.06 | 1.41 ± 0.14 | 1.31 ± 0.05 | 1.35 ± 0.02 | 1.40 ± 0.07 | 1.34 ± 0.05 | NS | NS | NS |
| HPI | 0.37 ± 0.03 | 0.43 ± 0.08 | 0.38 ± 0.01 | 0.40 ± 0.08 | 0.43 ± 0.06 | 0.40 ± 0.11 | NS | NS | NS |
| PI | 1.71 ± 0.49 B | 1.71 ± 0.10 B | 1.61 ± 0.06 B | 2.09 ± 0.18 A | 2.09 ± 0.28 A | 1.96 ± 0.30 A | * | NS | NS |
| SCD-16 | 2.10 ± 2.02 y | 5.00 ± 0.31 x | 4.34 ± 0.31 x | 5.54 ± 0.06 x | 5.48 ± 0.12 x | 5.71 ± 0.50 x | *** | * | * |
| SCD-18 | 56.45 ± 5.47 | 59.58 ± 3.89 | 55.49 ± 1.04 | 59.17 ± 12.68 | 60.86 ± 4.40 | 54.21 ± 11.46 | NS | NS | NS |
| EI | 57.51 ± 1.53 | 57.18 ± 2.27 | 55.65 ± 0.85 | 55.97 ± 0.27 | 56.91 ± 1.13 | 55.81 ± 0.68 | NS | NS | NS |
| TE | 0.008 ± 0.00 B | 0.014 ± 0.00 B | 0.015 ± 0.01 B | 0.018 ± 0.00 A | 0.016 ± 0.00 A | 0.16 ± 0.00 A | * | NS | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phoemchalard, C.; Prommachat, R.; Tathong, T.; Uriyapongson, S. Effect of Cassava Bioethanol By-Products and Crude Palm Oil Feeding on Fatty Acid Composition of Beef Meat and Fat in Crossbred Thai Indigenous Heifers. Animals 2024, 14, 3478. https://doi.org/10.3390/ani14233478
Phoemchalard C, Prommachat R, Tathong T, Uriyapongson S. Effect of Cassava Bioethanol By-Products and Crude Palm Oil Feeding on Fatty Acid Composition of Beef Meat and Fat in Crossbred Thai Indigenous Heifers. Animals. 2024; 14(23):3478. https://doi.org/10.3390/ani14233478
Chicago/Turabian StylePhoemchalard, Chirasak, Ronnachai Prommachat, Tanom Tathong, and Suthipong Uriyapongson. 2024. "Effect of Cassava Bioethanol By-Products and Crude Palm Oil Feeding on Fatty Acid Composition of Beef Meat and Fat in Crossbred Thai Indigenous Heifers" Animals 14, no. 23: 3478. https://doi.org/10.3390/ani14233478
APA StylePhoemchalard, C., Prommachat, R., Tathong, T., & Uriyapongson, S. (2024). Effect of Cassava Bioethanol By-Products and Crude Palm Oil Feeding on Fatty Acid Composition of Beef Meat and Fat in Crossbred Thai Indigenous Heifers. Animals, 14(23), 3478. https://doi.org/10.3390/ani14233478