Methodology Advances in Vertebrate Age Estimation


Abstract
Simple Summary
Abstract
1. Introduction
2. Destructive Samples
2.1. Skeletons and the Most Calcified Structures of the Body
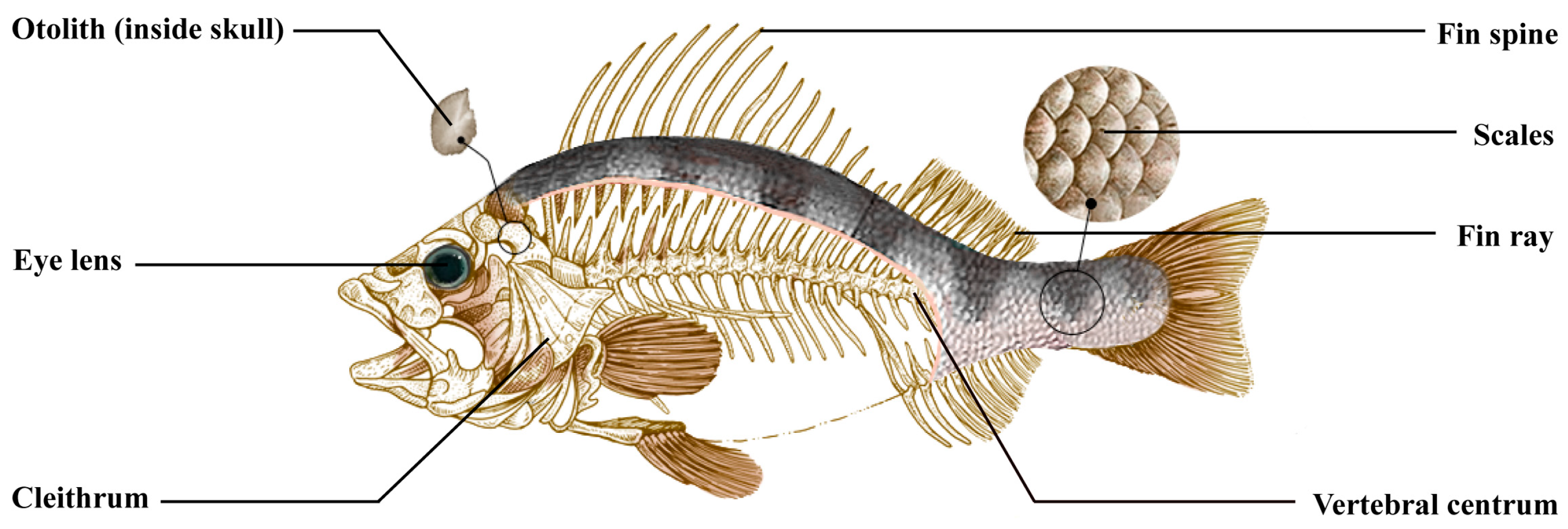
2.2. Eyeball Lens
2.3. Otoliths
3. Non-Destructive Samples
3.1. Blood
3.2. Scales
3.3. Fin Rays
3.4. Fin Spines
3.5. Teeth
4. Non-Invasive Samples
4.1. Feces
4.2. Physical Characteristics
4.3. Scute
4.4. Voice
4.5. Hair
5. Accuracy Assessment
5.1. Fish
5.2. Amphibia and Reptilia
5.3. Aves
5.4. Mammalia
6. The Future of Age Estimation
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Paoli-Iseppi, R.; Deagle, B.E.; Polanowski, A.M.; McMahon, C.R.; Dickinson, J.L.; Hindell, M.A.; Jarman, S.N. Age estimation in a long-lived seabird (Ardenna tenuirostris) using DNA methylation-based biomarkers. Mol. Ecol. Resour. 2019, 19, 411–425. [Google Scholar] [CrossRef]
- Froy, H.; Phillips, R.A.; Wood, A.G.; Nussey, D.H.; Lewis, S. Age-related variation in reproductive traits in the wandering albatross: Evidence for terminal improvement following senescence. Ecol. Lett. 2013, 16, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Gianuca, D.; Phillips, R.A.; Townley, S.; Votier, S.C. Global patterns of sex-and age-specific variation in seabird bycatch. Biol. Conserv. 2017, 205, 60–76. [Google Scholar] [CrossRef]
- Scott, M.E. The impact of infection and disease on animal populations: Implications for conservation biology. Conserv. Biol. 1988, 2, 40–56. [Google Scholar] [CrossRef]
- Tomaszewicz, C.N.T.; Seminoff, J.A.; Avens, L.; Goshe, L.R.; Peckham, S.H.; Rguez-Baron, J.M.; Bickerman, K.; Kurle, C.M. Age and residency duration of loggerhead turtles at a North Pacific bycatch hotspot using skeletochronology. Biol. Conserv. 2015, 186, 134–142. [Google Scholar] [CrossRef]
- Sajeevan, M.; Kurup, B.M. Age and growth of cobia, Rachycentron canadum (Linnaeus, 1766) occurring in North West coast of India. Indian J. Geo-Mar. Sci. 2017, 46, 1390–1395. [Google Scholar]
- Wiedower, E.E.; Kouba, A.J.; Vance, C.K.; Hansen, R.L.; Stuth, J.W.; Tolleson, D.R. Fecal near infrared spectroscopy to discriminate physiological status in giant pandas. PLoS ONE 2012, 7, e38908. [Google Scholar] [CrossRef] [PubMed]
- Horvath, S.; Lu, A.T.; Haghani, A.; Zoller, J.A.; Li, C.Z.; Lim, A.R.; Brooke, R.T.; Raj, K.; Serres-Armero, A.; Dreger, D.L. DNA methylation clocks for dogs and humans. Proc. Natl. Acad. Sci. USA 2022, 119, e2120887119. [Google Scholar] [CrossRef]
- Taberlet, P.; Waits, L.P.; Luikart, G. Noninvasive genetic sampling: Look before you leap. Trends Ecol. Evol. 1999, 14, 323–327. [Google Scholar] [CrossRef]
- Dessauer, H.; Hafner, M. Collections of Frozen Tissues: Value, Management, Field and Laboratory Procedures, and Directory of Existing Collections; Association of Systematics Collections: Lawrence, KS, USA, 1984. [Google Scholar]
- Morin, P. Noninvasive genotyping for vertebrate conservation. In Molecular Genetics Approaches to Conservation; Oxford University Press: Oxford, UK, 1992. [Google Scholar]
- Ming, L.; Fuwen, W.; Gang, R.; Shengguo, F.; Zuojian, F. Application of noninvasive sampling in conservation genetics. Acta Zool. Sin. 2001, 47, 242–338. [Google Scholar]
- Campana, S. Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J. Fish Biol. 2001, 59, 197–242. [Google Scholar] [CrossRef]
- Devine, J.A.; Baker, K.D.; Haedrich, R.L. Deep-sea fishes qualify as endangered. Nature 2006, 439, 29. [Google Scholar] [CrossRef] [PubMed]
- Beamish, R.; McFarlane, G.A. A discussion of the importance of aging errors, and an application to walleye pollock: The world’s largest fishery. In Recent Developments in Fish Otolith Research; University of South Carolina Press: Columbia, SC, USA, 1995; pp. 545–565. [Google Scholar]
- Castanet, J. Age estimation and longevity in reptiles. Gerontology 1994, 40, 174–192. [Google Scholar] [CrossRef] [PubMed]
- Zug, G.R.; Chaloupka, M.; Balazs, G.H. Age and growth in olive ridley seaturtles (Lepidochelys olivacea) from the North-central Pacific: A skeletochronological analysis. Mar. Ecol. 2006, 27, 263–270. [Google Scholar] [CrossRef]
- Kurita, T.; Toda, M. Validation and application of skeletochronology for age determination of the Ryukyu ground gecko, Goniurosaurus kuroiwae (Squamata: Eublepharidae). Asian Herpetol. Res. 2013, 4, 223–241. [Google Scholar] [CrossRef]
- Meunier, F.J. Skeletochronological studies of cyclical growth of freshwater fishes in French Guiana. A review. Cybium 2012, 36, 55–62. [Google Scholar]
- Khonsue, W.; Matsui, M.; Misawa, Y. Age determination by skeletochronology of Rana nigrovittata, a frog from tropical forest of Thailand. Zool. Sci. 2000, 17, 253–257. [Google Scholar] [CrossRef]
- Matsuki, T.; Matsui, M. The validity of skeletochronology in estimating ages of Japanese clouded salamander, Hynobius nebulosus (Amphibia, Caudata). Curr. Herpetol. 2009, 28, 41–48. [Google Scholar]
- Coles, W.C.; Musick, J.A.; Williamson, L.A. Skeletochronology validation from an adult loggerhead (Caretta caretta). Copeia 2001, 2001, 240–242. [Google Scholar] [CrossRef]
- Castanet, J.; Croci, S.; Aujard, F.; Perret, M.; Cubo, J.; de Margerie, E. Lines of arrested growth in bone and age estimation in a small primate: Microcebus murinus. J. Zool. 2004, 263, 31–39. [Google Scholar] [CrossRef]
- Khan, M.A.; Khan, S. Comparison of age estimates from scale, opercular bone, otolith, vertebrae and dorsal fin ray in Labeo rohita (Hamilton), Catla catla (Hamilton) and Channa marulius (Hamilton). Fish. Res. 2009, 100, 255–259. [Google Scholar] [CrossRef]
- Ma, B.; Xie, C.; Huo, B.; Yang, X.; Li, P. Age validation, and comparison of otolith, vertebra and opercular bone for estimating age of Schizothorax o’connori in the Yarlung Tsangpo River, Tibet. Environ. Biol. Fishes 2011, 90, 159–169. [Google Scholar] [CrossRef]
- Sharma, C.; Borgstrøm, R. Age determination and backcalculation of pike length through use of the metapterygoid bone. J. Fish Biol. 2007, 70, 1636–1641. [Google Scholar] [CrossRef]
- Blackwell, B.G.; Kaufman, T.M.; Moos, T.S. An assessment of calcified structures for estimating northern pike ages. N. Am. J. Fish. Manag. 2016, 36, 964–974. [Google Scholar] [CrossRef]
- Korostelev, N.; Frey, P.; Orlov, A. Using different hard structures to estimate the age of deep-sea fishes: A case study of the Pacific flatnose, Antimora microlepis (Moridae, Gadiformes, Teleostei). Fish. Res. 2020, 232, 105731. [Google Scholar] [CrossRef]
- Scharer, R.M.; Patterson III, W.F.; Carlson, J.K.; Poulakis, G.R. Age and growth of endangered smalltooth sawfish (Pristis pectinata) verified with LA-ICP-MS analysis of vertebrae. PLoS ONE 2012, 7, e47850. [Google Scholar] [CrossRef] [PubMed]
- Phelps, Q.E.; Edwards, K.R.; Willis, D.W. Precision of five structures for estimating age of common carp. N. Am. J. Fish. Manag. 2007, 27, 103–105. [Google Scholar] [CrossRef]
- Luo, W.-W.; Liu, C.-S.; Cao, X.-J.; Huang, L.-F.; Huang, S.-Q. Precision of age estimations from scales, otoliths, vertebrae, opercular bones and cleithra of two loaches, Misgurnus anguillicaudatus and Paramisgurnus dabryanus. Folia Zool. 2016, 65, 183–188. [Google Scholar] [CrossRef]
- Sanfelice, D.; Molina-Schiller, D.; De Freitas, T.R. Development and growth in skulls of three Otariidae species: A comparative morphometric study. J. Mar. Biol. Assoc. UK 2018, 98, 1801–1815. [Google Scholar] [CrossRef]
- Kryštufek, B.; Pistotnik, M.; Sedmak Časar, K. Age determination and age structure in the edible dormouse Glis glis based on incremental bone lines. Mammal Rev. 2005, 35, 210–214. [Google Scholar] [CrossRef]
- Augusteyn, R.C. Growth of the lens: In vitro observations. Clin. Exp. Optom. 2008, 91, 226–239. [Google Scholar] [CrossRef] [PubMed]
- Augusteyn, R.C. Growth of the eye lens: I. Weight accumulation in multiple species. Mol. Vis. 2014, 20, 410. [Google Scholar]
- Augusteyn, R.C.; Cake, M.A. Post-mortem water uptake by sheep lenses left in situ. Mol. Vis. 2005, 11, 749–751. [Google Scholar]
- Forsyth, D.M.; Garel, M.; McLeod, S.R. Estimating age and age class of harvested hog deer from eye lens mass using frequentist and Bayesian methods. Wildl. Biol. 2016, 22, 137–143. [Google Scholar] [CrossRef]
- McLeod, S.; Druhan, J.; Hacker, R. Estimating the age of kangaroos using eye lens weight. Wildl. Res. 2006, 33, 25–28. [Google Scholar] [CrossRef]
- Lord, R.D. The lens as an indicator of age in the gray fox. J. Mammal. 1961, 42, 109–111. [Google Scholar] [CrossRef]
- Lord, R.D. Growth of the lens of the pampas gray fox (Dusicyon gymnocercus antiquus) and the Patagonian gray fox (Dusicyon griseus griseus). J. Mammal. 1966, 47, 536–538. [Google Scholar] [CrossRef]
- Lombaard, L.J. Age determination and growth curves in the black-backed jackal, Canis mesomelas Schreber, 1775 (Carnivora: Canidae). Ann. Transvaal Mus. 1971, 27, 135–169. [Google Scholar]
- George, J.C.; Bada, J.; Zeh, J.; Scott, L.; Brown, S.E.; O’Hara, T.; Suydam, R. Age and growth estimates of bowhead whales (Balaena mysticetus) via aspartic acid racemization. Can. J. Zool. 1999, 77, 571–580. [Google Scholar] [CrossRef]
- Olsen, E.; Sunde, J. Age determination of minke whales (Balaenoptera acutorostrata) using the aspartic acid racemization technique. Sarsia 2002, 87, 1–8. [Google Scholar] [CrossRef]
- Bada, J.L. Amino acid racemization reactions and their geochemical implications. Naturwissenschaften 1975, 62, 71–79. [Google Scholar] [CrossRef]
- Garde, E.; Heide-Jørgensen, M.P.; Hansen, S.H.; Nachman, G.; Forchhammer, M.C. Age-specific growth and remarkable longevity in narwhals (Monodon monoceros) from West Greenland as estimated by aspartic acid racemization. J. Mammal. 2007, 88, 49–58. [Google Scholar] [CrossRef]
- Rosa, C.; Zeh, J.; Craig George, J.; Botta, O.; Zauscher, M.; Bada, J.; O’Hara, T.M. Age estimates based on aspartic acid racemization for bowhead whales (Balaena mysticetus) harvested in 1998–2000 and the relationship between racemization rate and body temperature. Mar. Mammal Sci. 2013, 29, 424–445. [Google Scholar] [CrossRef]
- Boye, T.K.; Garde, E.; Nielsen, J.; Hedeholm, R.; Olsen, J.; Simon, M. Estimating the age of West Greenland humpback whales through aspartic acid racemization and eye lens bomb radiocarbon methods. Front. Mar. Sci. 2020, 6, 811. [Google Scholar] [CrossRef]
- Garde, E.; Hansen, S.H.; Ditlevsen, S.; Tvermosegaard, K.B.; Hansen, J.; Harding, K.C.; Heide-Jørgensen, M.P. Life history parameters of narwhals (Monodon monoceros) from Greenland. J. Mammal. 2015, 96, 866–879. [Google Scholar] [CrossRef]
- Nielsen, J.; Hedeholm, R.B.; Heinemeier, J.; Bushnell, P.G.; Christiansen, J.S.; Olsen, J.; Ramsey, C.B.; Brill, R.W.; Simon, M.; Steffensen, K.F. Eye lens radiocarbon reveals centuries of longevity in the Greenland shark (Somniosus microcephalus). Science 2016, 353, 702–704. [Google Scholar] [CrossRef]
- Lynnerup, N.; Kjeldsen, H.; Heegaard, S.; Jacobsen, C.; Heinemeier, J. Radiocarbon dating of the human eye lens crystallines reveal proteins without carbon turnover throughout life. PLoS ONE 2008, 3, e1529. [Google Scholar] [CrossRef] [PubMed]
- Hedeholm, R.; Qvist, T.; Frausing, M.; Olsen, J.; Nielsen, J.; Grønkjær, P. Age of black dogfish (Centroscyllium fabricii) estimated from fin spines growth bands and eye lens bomb radiocarbon dating. Polar Biol. 2021, 44, 751–759. [Google Scholar] [CrossRef]
- Hamady, L.L.; Natanson, L.J.; Skomal, G.B.; Thorrold, S.R. Vertebral bomb radiocarbon suggests extreme longevity in white sharks. PLoS ONE 2014, 9, e84006. [Google Scholar] [CrossRef][Green Version]
- Campana, S.E. Chemistry and composition of fish otoliths: Pathways, mechanisms and applications. Mar. Ecol. Prog. Ser. 1999, 188, 263–297. [Google Scholar] [CrossRef]
- Popper, A.N.; Ramcharitar, J.; Campana, S.E. Why otoliths? Insights from inner ear physiology and fisheries biology. Mar. Freshw. Res. 2005, 56, 497–504. [Google Scholar] [CrossRef]
- Thomas, O.R.; Swearer, S.E. Otolith biochemistry—A review. Rev. Fish. Sci. Aquac. 2019, 27, 458–489. [Google Scholar] [CrossRef]
- Campana, S.E.; Secor, D.H.; Dean, J.M. Recent Developments in Fish Otolith Research; University of South Carolina Press: Columbia, SC, USA, 1995. [Google Scholar]
- Ross, J.R.; Crosby, J.D.; Kosa, J.T. Accuracy and precision of age estimation of crappies. N. Am. J. Fish. Manag. 2005, 25, 423–428. [Google Scholar] [CrossRef]
- Snow, R.A.; Porta, M.J.; Long, J.M. Precision of four otolith techniques for estimating age of white perch from a thermally altered reservoir. N. Am. J. Fish. Manag. 2018, 38, 725–733. [Google Scholar] [CrossRef]
- Buslov, A.; Ovsyannikov, E. The Validity of Otolith-Based Age Estimates for the Walleye Pollock Theragra chalcogramma (Pallas, 1814) from the Eastern Sea of Okhotsk. Russ. J. Mar. Biol. 2019, 45, 370–376. [Google Scholar] [CrossRef]
- Pacheco, C.; Bustamante, C.; Araya, M. Mass-effect: Understanding the relationship between age and otolith weight in fishes. Fish Fish. 2021, 22, 623–633. [Google Scholar] [CrossRef]
- Khan, M.A.; Nazir, A.; Banday, U.Z. Utility of otolith weight to estimate age of Labeo bata (Actinopterygii: Cypriniformes: Cyprinidae) inhabiting the Ganga River. Acta Ichthyol. Piscat. 2018, 48, 257–260. [Google Scholar] [CrossRef]
- Andrews, A.H.; Kerr, L.A.; Cailliet, G.M.; Brown, T.A.; Lundstrom, C.C.; Stanley, R.D. Age validation of canary rockfish (Sebastes pinniger) using two independent otolith techniques: Lead-radium and bomb radiocarbon dating. Mar. Freshw. Res. 2007, 58, 531–541. [Google Scholar] [CrossRef]
- Andrews, A.H.; Cailliet, G.M.; Coale, K.H. Age and growth of the Pacific grenadier (Coryphaenoides acrolepis) with age estimate validation using an improved radiometric ageing technique. Can. J. Fish. Aquat. Sci. 1999, 56, 1339–1350. [Google Scholar] [CrossRef]
- Kimura, D.K.; Kastelle, C.R. Perspectives on the relationship between otolith growth and the conversion of isotope activity ratios to fish ages. Can. J. Fish. Aquat. Sci. 1995, 52, 2296–2303. [Google Scholar] [CrossRef]
- Francis, R.C. The precision of otolith radiometric ageing of fish and the effect of within-sample heterogeneity. Can. J. Fish. Aquat. Sci. 2003, 60, 441–447. [Google Scholar] [CrossRef]
- Andrews, A.; Ashford, J.; Brooks, C.; Krusic-Golub, K.; Duhamel, G.; Belchier, M.; Lundstrom, C.; Cailliet, G. Lead–radium dating provides a framework for coordinating age estimation of Patagonian toothfish (Dissostichus eleginoides) between fishing areas. Mar. Freshw. Res. 2011, 62, 781–789. [Google Scholar] [CrossRef]
- Shervette, V.R.; Rivera Hernández, J.M. Queen triggerfish Balistes vetula: Validation of otolith-based age, growth, and longevity estimates via application of bomb radiocarbon. PLoS ONE 2022, 17, e0262281. [Google Scholar] [CrossRef] [PubMed]
- Healy, J.; Helser, T.E.; Benson, I.M.; Tornabene, L. Aging Pacific cod (Gadus macrocephalus) from otoliths using Fourier-transformed near-infrared spectroscopy. Ecosphere 2021, 12, e03697. [Google Scholar] [CrossRef]
- Horvath, S. DNA methylation age of human tissues and cell types. Genome Biol. 2013, 14, 1–20. [Google Scholar] [CrossRef]
- Hannum, G.; Guinney, J.; Zhao, L.; Zhang, L.; Hughes, G.; Sadda, S.; Klotzle, B.; Bibikova, M.; Fan, J.-B.; Gao, Y. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol. Cell 2013, 49, 359–367. [Google Scholar] [CrossRef]
- Vidal-Bralo, L.; Lopez-Golan, Y.; Gonzalez, A. Simplified assay for epigenetic age estimation in whole blood of adults. Front. Genet. 2016, 7, 126. [Google Scholar] [CrossRef]
- Thompson, M.J.; Horvath, S.; Pellegrini, M. An epigenetic aging clock for dogs and wolves. Aging 2017, 9, 1055. [Google Scholar] [CrossRef]
- Ito, G.; Yoshimura, K.; Momoi, Y. Analysis of DNA methylation of potential age-related methylation sites in canine peripheral blood leukocytes. J. Vet. Med. Sci. 2017, 79, 745–750. [Google Scholar] [CrossRef][Green Version]
- Stacy, B.A.; Whitaker, N. Hematology and blood biochemistry of captive mugger crocodiles (Crocodylus palustris). J. Zoo Wildl. Med. 2000, 31, 339–347. [Google Scholar]
- Rørtveit, R.; Sævik, B.K.; Eggertsdóttir, A.V.; Skancke, E.; Lingaas, F.; Thoresen, S.I.; Jansen, J.H. Age-related changes in hematologic and serum biochemical variables in dogs aged 16–60 days. Vet. Clin. Pathol. 2015, 44, 47–57. [Google Scholar] [CrossRef]
- Tóthová, C.; Nagy, O.; Kováč, G. Changes in the concentrations of serum protein fractions in calves with age and nutrition. Ital. J. Anim. Sci. 2014, 13, 2993. [Google Scholar] [CrossRef][Green Version]
- Khalil, A.M.; Murakami, N.; Kaseda, Y. Relashionship between plasma testosterone concentrations and age, breeding season and harem size in misaki feral horses. J. Vet. Med. Sci. 1998, 60, 643–645. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Husakova, T.; Pavlata, L.; Pechova, A.; Hauptmanova, K.; Pitropovska, E.; Tichy, L. Reference values for biochemical parameters in blood serum of young and adult alpacas (Vicugna pacos). Animal 2014, 8, 1448–1455. [Google Scholar] [CrossRef] [PubMed]
- Jakubas, D.; Wojczulanis-Jakubas, K.; Glac, W. Variation of the reed bunting (Emberiza schoeniclus) body condition and haematological parameters in relation to sex, age and season. In Annales Zoologici Fennici; Finnish Zoological and Botanical Publishing Board: Helsinki, Finland, 2011; Volume 48, pp. 243–250. [Google Scholar]
- Fisher, J. Spacing of scale circuli versus growth rate in young coho salmon. Fish. Bull. 1990, 88, 637–643. [Google Scholar]
- Sire, J.-Y.; Akimenko, M.-A. Scale development in fish: A review, with description of sonic hedgehog (shh) expression in the zebrafish (Danio rerio). Int. J. Dev. Biol. 2003, 48, 233–247. [Google Scholar] [CrossRef] [PubMed]
- Tzadik, O.E.; Curtis, J.S.; Granneman, J.E.; Kurth, B.N.; Pusack, T.J.; Wallace, A.A.; Hollander, D.J.; Peebles, E.B.; Stallings, C.D. Chemical archives in fishes beyond otoliths: A review on the use of other body parts as chronological recorders of microchemical constituents for expanding interpretations of environmental, ecological, and life-history changes. Limnol. Oceanogr. Methods 2017, 15, 238–263. [Google Scholar] [CrossRef]
- Lee, R.T.H.; Thiery, J.P.; Carney, T.J. Dermal fin rays and scales derive from mesoderm, not neural crest. Curr. Biol. 2013, 23, R336–R337. [Google Scholar] [CrossRef]
- Singh, S.P.; Holdway, J.E.; Poss, K.D. Regeneration of amputated zebrafish fin rays from de novo osteoblasts. Dev. Cell 2012, 22, 879–886. [Google Scholar] [CrossRef]
- Sweet, D.E.; Compton, R.I.; Hubert, W.A. Age and growth of bluehead suckers and flannelmouth suckers in headwater tributaries, Wyoming. West. North Am. Nat. 2009, 69, 35–41. [Google Scholar] [CrossRef]
- Sylvester, R.M.; Berry, C.R., Jr. Comparison of white sucker age estimates from scales, pectoral fin rays, and otoliths. N. Am. J. Fish. Manag. 2006, 26, 24–31. [Google Scholar] [CrossRef]
- Kerr, L.A.; Campana, S.E. Chemical composition of fish hard parts as a natural marker of fish stocks. In Stock Identification Methods; Elsevier: Amsterdam, The Netherlands, 2014; pp. 205–234. [Google Scholar]
- Bruch, R.M.; Campana, S.E.; Davis-Foust, S.L.; Hansen, M.J.; Janssen, J. Lake Sturgeon Age Validation using Bomb Radiocarbon and Known-Age Fish. Trans. Am. Fish. Soc. 2009, 138, 361–372. [Google Scholar] [CrossRef]
- Klein, Z.B.; Bonvechio, T.F.; Bowen, B.R.; Quist, M.C. Precision and Accuracy of Age Estimates Obtained from Anal Fin Spines, Dorsal Fin Spines, and Sagittal Otoliths for Known-Age Largemouth Bass. Southeast. Nat. 2017, 16, 225–234. [Google Scholar] [CrossRef]
- Arai, T.; Levin, A.; Boltunov, A.; Miyazaki, N. Migratory history of the Russian sturgeon Acipenser guldenstadti in the Caspian Sea, as revealed by pectoral fin spine Sr: Ca ratios. Mar. Biol. 2002, 141, 315–319. [Google Scholar]
- Baremore, I.E.; Rosati, J.D. A validated, minimally deleterious method for aging sturgeon. Fish. Bull. 2014, 112, 274–282. [Google Scholar] [CrossRef]
- James, K.C.; Natanson, L.J.; Flight, C.; Tribuzio, C.; Hoey, J.; Candless, C.M. Validation of the use of vertebrae and dorsal-fin spines for age determination of spiny dogfish (Squalus acanthias) in the western North Atlantic Ocean. Fish. Bull. 2021, 119, 41–49. [Google Scholar] [CrossRef]
- Kopf, R.K.; Drew, K.; Humphreys, R.L. Age estimation of billfishes (Kajikia spp.) using fin spine cross-sections: The need for an international code of practice. Aquat. Living Resour. 2010, 23, 13–23. [Google Scholar] [CrossRef]
- Weber, M.J.; Brown, M.L. Comparison of common carp (Cyprinus carpio) age estimates derived from dorsal fin spines and pectoral fin rays. J. Freshw. Ecol. 2011, 26, 195–202. [Google Scholar] [CrossRef]
- Morehouse, R.L.; Donabauer, S.B.; Grier, A.C. Estimating largemouth bass age: Precision and comparisons among scales, pectoral fin rays, and dorsal fin spines as nonlethal methods. Fish. Aquac. J. 2013, 4, 1–9. [Google Scholar]
- Natanson, L.J.; Skomal, G.B. Age and growth of the white shark, Carcharodon carcharias, in the western North Atlantic Ocean. Mar. Freshw. Res. 2015, 66, 387–398. [Google Scholar] [CrossRef]
- Natanson, L.J.; Skomal, G.B.; Hoffmann, S.L.; Porter, M.E.; Goldman, K.J.; Serra, D. Age and growth of sharks: Do vertebral band pairs record age? Mar. Freshw. Res. 2018, 69, 1440–1452. [Google Scholar] [CrossRef]
- Rigby, C.L.; Wedding, B.B.; Grauf, S.; Simpfendorfer, C.A. The utility of near infrared spectroscopy for age estimation of deepwater sharks. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2014, 94, 184–194. [Google Scholar] [CrossRef]
- Asmus, J.; Weckerly, F.W. Evaluating precision of cementum annuli analysis for aging mule deer from southern California. J. Wildl. Manag. 2011, 75, 1194–1199. [Google Scholar] [CrossRef]
- Foley, A.M.; Lewis, J.S.; Cortez, O.; Hellickson, M.W.; Hewitt, D.G.; DeYoung, R.W.; DeYoung, C.A.; Schnupp, M.J. Accuracies and biases of ageing white-tailed deer in semiarid environments. Wildl. Res. 2021, 49, 237–249. [Google Scholar] [CrossRef]
- Rust, L.B.; Danil, K.; Melin, S.R.; Wilkerson, B. Accuracy and precision of age determination using growth layer groups for California sea lions (Zalophus californianus) with known ages. Mar. Mammal Sci. 2019, 35, 1355–1368. [Google Scholar] [CrossRef] [PubMed]
- Habermehl, K.-H. Die Altersbestimmung bei Haus-und Labortieren; P. Parey: Berlin, Germany, 1975. [Google Scholar]
- Rosatte, R.; Silver, A.; Gibson, M.; Chisholm, B.; Cool, N. Correlation between age estimates for elk, Cervus elaphus, using tooth wear/eruption patterns and counts of annuli in tooth cementum. Can. Field Nat. 2007, 121, 214–215. [Google Scholar] [CrossRef]
- Olifiers, N.; de Cassia Bianchi, R.; D’Andrea, P.S.; Mourao, G.; Gompper, M.E. Estimating age of carnivores from the Pantanal region of Brazil. Wildl. Biol. 2010, 16, 389–399. [Google Scholar] [CrossRef]
- Gipson, P.S.; Ballard, W.B.; Nowak, R.M.; Mech, L.D. Accuracy and precision of estimating age of gray wolves by tooth wear. J. Wildl. Manag. 2000, 64, 752–758. [Google Scholar] [CrossRef]
- Gol’din, P.; Godlevska, L.; Ghazali, M. Age-related changes in the teeth of two bat species: Dental wear, pulp cavity and dentine growth layers. Acta Chiropterol. 2018, 20, 519–530. [Google Scholar] [CrossRef]
- Storz, J.F.; Bhat, H.R.; Kunz, T.H. Social structure of a polygynous tent-making bat, Cynopterus sphinx (Megachiroptera). J. Zool. 2000, 251, 151–165. [Google Scholar] [CrossRef]
- Laws, R. A new method of age determination for mammals. Nature 1952, 169, 972–973. [Google Scholar] [CrossRef] [PubMed]
- Garde, E.; Frie, A.K.; Dunshea, G.; Hansen, S.H.; Kovacs, K.M.; Lydersen, C. Harp seal ageing techniques—Teeth, aspartic acid racemization, and telomere sequence analysis. J. Mammal. 2010, 91, 1365–1374. [Google Scholar] [CrossRef]
- Rolandsen, C.M.; Solberg, E.J.; Heim, M.; Holmstrøm, F.; Solem, M.I.; Sæther, B.-E. Accuracy and repeatability of moose (Alces alces) age as estimated from dental cement layers. Eur. J. Wildl. Res. 2008, 54, 6–14. [Google Scholar] [CrossRef]
- Sharma, V.; Gupta, S.K.; Sharma, C.P.; Sankar, K.; Goyal, S.P. Development of a fast and low-cost age determination method in spotted deer, Axis axis. Folia Zool. 2018, 67, 186–197. [Google Scholar]
- Cool, S.; Bennet, M.; Romaniuk, K. Age estimation of pteropodid bats (Megachiroptera) from hard tissue parameters. Wildl. Res. 1994, 21, 353–363. [Google Scholar] [CrossRef]
- Roulichova, J.; Andera, M. Simple method of age determination in red fox, Vulpes vulpes. Folia Zool. 2007, 56, 440. [Google Scholar]
- Fàbregas, M.C.; Garcés-Narro, C. Validation of gum-line recession as a reliable technique to age tigers. Eur. J. Wildl. Res. 2014, 60, 947–950. [Google Scholar] [CrossRef]
- Currier, M.J.P. An Age Estimation Technique and Some Normal Blood Values for Mountain Lions (Felis concolor); Colorado State University: Fort Collins, CO, USA, 1979. [Google Scholar]
- Laundré, J.W.; Hernández, L.; Streubel, D.; Altendorf, K.; González, C.L. Aging mountain lions using gum-line recession. Wildl. Soc. Bull. 2000, 28, 963–966. [Google Scholar]
- Peers, M.J.; Majchrzak, Y.N.; Studd, E.K.; Menzies, A.K.; Kukka, P.M.; Konkolics, S.M.; Boonstra, R.; Boutin, S.; Jung, T.S. Evaluation of Gum-line Recession for Aging Lynx (Lynx canadensis). Wildl. Soc. Bull. 2021, 45, 706–710. [Google Scholar] [CrossRef]
- WILkINSON, G.S.; Brunet-Rossinni, A. Methods for age estimation and the study of senescence in bats. In Ecological and Behavioral Methods for the Study of Bats; Johns Hopkins University Press: Baltimore, MD, USA, 2009; pp. 315–325. [Google Scholar]
- Zhixiao, L.; Helin, S.; Yuanguang, L.; Zhaoding, W.; Denghai, Z.; Heping, D. The current status and conservation of the isolated population of alpine musk deer (Moschus chrysogaster) at Helan Mountain. Acta Ecol. Sin. 2000, 20, 463–467. [Google Scholar]
- Zhixiao, L.; Qiang, L.; Fagong, K. Some ecological characteristics of the isolated population of alphine musk deer (Moschus chrysogaster) in the Xinglong forest, Gangsu Province. Acta Ecol. Sin. 2001, 21, 964–968. [Google Scholar]
- Yan, X.; Du, Z.; Bai, S.; Zuo, Y.; Zhou, X.; Kou, J.; Yan, J.; Zhang, J.; Li, P.; You, M. Potential Applicability of Fecal NIRs: A Review. Spectrosc. Spectral Anal. 2015, 35, 3382–3387. [Google Scholar]
- Rueda, M.; Rebollo, S.; Bravo, L.G. Age and season determine European rabbit habitat use in Mediterranean ecosystems. Acta Oecol. 2008, 34, 266–273. [Google Scholar] [CrossRef]
- Kongrit, C.; Siripunkaw, C. Determination of age and construction of population age structure of wild Asian elephants based on dung bolus circumference. Thai J. Vet. Med. 2017, 47, 145. [Google Scholar] [CrossRef]
- Austin, S.H.; Robinson, T.R.; Robinson, W.D.; Ricklefs, R.E. Potential biases in estimating the rate parameter of sigmoid growth functions. Methods Ecol. Evol. 2011, 2, 43–51. [Google Scholar] [CrossRef]
- Einum, S.; Forseth, T.; Finstad, A.G. Individual variation in response to intraspecific competition: Problems with inference from growth variation measures. Methods Ecol. Evol. 2012, 3, 438–444. [Google Scholar] [CrossRef]
- Paine, C.T.; Marthews, T.R.; Vogt, D.R.; Purves, D.; Rees, M.; Hector, A.; Turnbull, L.A. How to fit nonlinear plant growth models and calculate growth rates: An update for ecologists. Methods Ecol. Evol. 2012, 3, 245–256. [Google Scholar] [CrossRef]
- Pardo, S.A.; Cooper, A.B.; Dulvy, N.K. Avoiding fishy growth curves. Methods Ecol. Evol. 2013, 4, 353–360. [Google Scholar] [CrossRef]
- Gislason, H.; Daan, N.; Rice, J.C.; Pope, J.G. Size, growth, temperature and the natural mortality of marine fish. Fish Fish. 2010, 11, 149–158. [Google Scholar] [CrossRef]
- Pauly, D. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. ICES J. Mar. Sci. 1980, 39, 175–192. [Google Scholar] [CrossRef]
- Lorenzen, K. Allometry of natural mortality as a basis for assessing optimal release size in fish-stocking programmes. Can. J. Fish. Aquat. Sci. 2000, 57, 2374–2381. [Google Scholar] [CrossRef]
- Hoenig, J. Empirical Use of Longevity Data to Estimate Mortality Rates. Fish. Bull. 2013, 82, 898–903. [Google Scholar]
- Andersen, K.H.; Beyer, J.E. Asymptotic size determines species abundance in the marine size spectrum. Am. Nat. 2006, 168, 54–61. [Google Scholar] [CrossRef]
- Botero-Delgadillo, E. Criterios morfométricos y cualitativos para la determinación de la edad y el sexo en Mionectes olivaceus (Tyrannidae). El Hornero 2010, 25, 9–16. [Google Scholar] [CrossRef]
- Green, D.J.; Whitehorne, I.B.; Taylor, A.L.; Drake, E.L. Wing morphology varies with age but not migratory habit in American Dippers. Wilson J. Ornithol. 2009, 121, 141–147. [Google Scholar] [CrossRef]
- Brown, W.P.; Alexander, A.L.; Alexander, D.A.; Zuefle, M.E.; Underwood, T.J. Estimating ages of House Wren nestlings based on body mass, wing chord length, and feather tract development patterns. N. Am. Bird Bander 2011, 36, 101–110. [Google Scholar]
- Mayberry, C.; Bencini, R.; Mawson, P.R.; Maloney, S.K. Estimating the age of western grey kangaroos (Macropus fuliginosus ocydromus). Aust. Mammal. 2017, 40, 254–261. [Google Scholar] [CrossRef]
- Sukumar, R. The Asian Elephant: Ecology and Management; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Arivazhagan, C.; Sukumar, R. Constructing age structures of Asian elephant populations: A comparison of two field methods of age estimation. Gajah 2008, 29, 11–16. [Google Scholar]
- Sharifi, M.; Akmali, V. Postnatal growth in the lesser mouse-eared bat, Myotis blythii, in captivity. Zool. Middle East 2006, 37, 13–20. [Google Scholar] [CrossRef]
- Elangovan, V.; Raghuram, H.; Yuvana Satya Priya, E.; Marimuthu, G. Postnatal growth, age estimation and development of foraging behaviour in the fulvous fruit bat Rousettus leschenaulti. J. Biosci. 2002, 27, 695–702. [Google Scholar] [CrossRef]
- Whitman, K.; Starfield, A.M.; Quadling, H.S.; Packer, C. Sustainable trophy hunting of African lions. Nature 2004, 428, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Van Horn, R.C.; Zug, B.; Appleton, R.D.; Velez-Liendo, X.; Paisley, S.; LaCombe, C. Photos provide information on age, but not kinship, of Andean bear. PeerJ 2015, 3, e1042. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Moustakas-Verho, J.E.; Cebra-Thomas, J.; Gilbert, S.F. Patterning of the turtle shell. Curr. Opin. Genet. Dev. 2017, 45, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.S.; Tracy, C.R.; Tracy, C.R. Estimating age of turtles from growth rings: A critical evaluation of the technique. Herpetologica 2003, 59, 178–194. [Google Scholar] [CrossRef]
- Howell, H.J.; Seigel, R.A. An examination of the accuracy of using plastral scute rings to age Spotted Turtles (Clemmys guttata). Chelonian Conserv. Biol. 2018, 17, 104–108. [Google Scholar] [CrossRef]
- Brooks, R.J.; Krawchuk, M.A.; Stevens, C.; Koper, N. Testing the precision and accuracy of age estimation using lines in scutes of Chelydra serpentina and Chrysemys picta. J. Herpetol. 1997, 31, 521–529. [Google Scholar] [CrossRef]
- Stoeger, A.S.; Zeppelzauer, M.; Baotic, A. Age-group estimation in free-ranging African elephants based on acoustic cues of low-frequency rumbles. Bioacoustics 2014, 23, 231–246. [Google Scholar] [CrossRef]
- Vaytina, T.; Shitikov, D. Age-related changes in song repertoire size and song type sharing in the Whinchat Saxicola rubetra. Bioacoustics 2019, 28, 140–154. [Google Scholar] [CrossRef]
- Hao, T.; Guo, J.; Liu, J.; Wang, J.; Liu, Z.; Cheng, X.; Li, J.; Ren, J.; Li, Z.; Yan, J. Predicting human age by detecting DNA methylation status in hair. Electrophoresis 2021, 42, 1255–1261. [Google Scholar] [CrossRef]
- Cattet, M.; Stenhouse, G.B.; Boulanger, J.; Janz, D.M.; Kapronczai, L.; Swenson, J.E.; Zedrosser, A. Can concentrations of steroid hormones in brown bear hair reveal age class? Conserv. Physiol. 2018, 6, coy001. [Google Scholar] [CrossRef]
- Schell, C.J.; Young, J.K.; Lonsdorf, E.V.; Mateo, J.M.; Santymire, R.M. Investigation of Techniques to Measure Cortisol and Testosterone Concentrations in Coyote Hair. Zoo. Biol. 2017, 36, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Carlitz, E.H.; Runge, J.-N.; König, B.; Winkler, L.; Kirschbaum, C.; Gao, W.; Lindholm, A.K. Steroid hormones in hair reveal sexual maturity and competition in wild house mice (Mus musculus domesticus). Sci. Rep. 2019, 9, 16925. [Google Scholar] [CrossRef] [PubMed]
- Ento, K.; Matsui, M. Estimation of age structure by skeletochronology of a population of Hynobius nebulosus in a breeding season (Amphibia, Urodela). Zool. Sci. 2002, 19, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Comas, M.; Reguera, S.; Zamora-Camacho, F.J.; Salvadó i Cabré, H.; Moreno-Rueda, G. Comparison of the effectiveness of phalanges vs. humeri and femurs to estimate lizard age with skeletochronology. In Animal Biodiversity and Conservation; Museu de Ciències Naturals de Barcelona: Barcelona, Spain, 2016; Volume 39, pp. 237–240. [Google Scholar]
- Amiot, C.; Ji, W.; Hill, S. Using plumage and behavioural development to age New Zealand Fantail nestlings. N. Z. J. Zool. 2015, 42, 35–43. [Google Scholar] [CrossRef]
- Costa, J.S.; Rocha, A.D.; Correia, R.A.; Alves, J.A. Developing and validating a nestling photographic aging guide for cavity-nesting birds: An example with the European Bee-eater (Merops apiaster). Avian Res. 2020, 11, 2. [Google Scholar] [CrossRef]
- Izzo, L.K.; Parrish, D.L.; Zydlewski, G.B.; Koenigs, R. Second fin ray shows promise for estimating ages of juvenile but not adult lake Sturgeon. North Am. J. Fish. Manag. 2021, 41, 217–228. [Google Scholar] [CrossRef]
- Helms, D. Shovelnose sturgeon, Scaphirhynchus platorynchus (Rafinesque), in the navigational impoundments of the upper Mississippi River. Iowa Conservation Commission. Fish. Res. Tech. Ser. 1974, 74, 68. [Google Scholar]
- Koch, J.; Steffensen, K.; Pegg, M. Validation of age estimates obtained from juvenile pallid sturgeon Scaphirhynchus albus pectoral fin spines. J. Appl. Ichthyol. 2011, 27, 209–212. [Google Scholar] [CrossRef]
- Kapfer, J.M.; Coggins, J.R.; Hay, R. Estimates of Population Size, Measurements of Sex Ratios, and Reported Mortality Rates for Bullsnakes (Pituophis catenifer sayi) at a Site in the Upper Midwestern United States. J. Herpetol. 2008, 42, 265–269, 265. [Google Scholar] [CrossRef]
- Wails, C.N.; Oswald, S.A.; Arnold, J.M. Are morphometrics sufficient for estimating age of pre-fledging birds in the field? A test using Common Terns (Sterna hirundo). PLoS ONE 2014, 9, e111987. [Google Scholar] [CrossRef]
- Nielsen, N.H.; Garde, E.; Heide-Jørgensen, M.P.; Lockyer, C.H.; Ditlevsen, S.; Òlafsdóttir, D.; Hansen, S.H. Application of a novel method for age estimation of a baleen whale and a porpoise. Mar. Mammal Sci. 2013, 29, E1–E23. [Google Scholar] [CrossRef]
- Stewart, R.; Campana, S.; Jones, C.; Stewart, B. Bomb radiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates. Can. J. Zool. 2006, 84, 1840–1852. [Google Scholar] [CrossRef]
- Evans, K.; Hindell, M.A.; Robertson, K.; Lockyer, C.; Rice, D. Factors affecting the precision of age determination of sperm whales (Physeter macrocephalus). J. Cetacean Res. Manag. 2002, 4, 193–201. [Google Scholar] [CrossRef]
- Garde, E.; Peter Heide-Jørgensen, M.; Ditlevsen, S.; Hansen, S.H. Aspartic acid racemization rate in narwhal (Monodon monoceros) eye lens nuclei estimated by counting of growth layers in tusks. Polar Res. 2012, 31, 15865. [Google Scholar] [CrossRef]
- Brunet-Rossinni, A.K.; Austad, S.N. Ageing studies on bats: A review. Biogerontology 2004, 5, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Horvath, S.; Zhang, Y.; Langfelder, P.; Kahn, R.S.; Boks, M.P.; van Eijk, K.; van den Berg, L.H.; Ophoff, R.A. Aging effects on DNA methylation modules in human brain and blood tissue. Genome Biol. 2012, 13, R97. [Google Scholar] [CrossRef]
- Day, K.; Waite, L.L.; Thalacker-Mercer, A.; West, A.; Bamman, M.M.; Brooks, J.D.; Myers, R.M.; Absher, D. Differential DNA methylation with age displays both common and dynamic features across human tissues that are influenced by CpG landscape. Genome Biol. 2013, 14, R102. [Google Scholar] [CrossRef]
- Freire-Aradas, A.; Phillips, C.; Girón-Santamaría, L.; Mosquera-Miguel, A.; Gómez-Tato, A.; de Cal, M.Á.C.; Álvarez-Dios, J.; Lareu, M.V. Tracking age-correlated DNA methylation markers in the young. Forensic Sci. Int. Genet. 2018, 36, 50–59. [Google Scholar] [CrossRef]
- Hong, S.R.; Shin, K.-J.; Jung, S.-E.; Lee, E.H.; Lee, H.Y. Platform-independent models for age prediction using DNA methylation data. Forensic Sci. Int. Genet. 2019, 38, 39–47. [Google Scholar] [CrossRef]
- Dettenborn, L.; Tietze, A.; Kirschbaum, C.; Stalder, T. The assessment of cortisol in human hair: Associations with sociodemographic variables and potential confounders. Stress 2012, 15, 578–588. [Google Scholar] [CrossRef]
- Fourie, N.H.; Jolly, C.J.; Phillips-Conroy, J.E.; Brown, J.L.; Bernstein, R.M. Variation of hair cortisol concentrations among wild populations of two baboon species (Papio anubis, P. hamadryas) and a population of their natural hybrids. Primates 2015, 56, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Phillip, J.M.; Wu, P.-H.; Gilkes, D.M.; Williams, W.; McGovern, S.; Daya, J.; Chen, J.; Aifuwa, I.; Lee, J.S.; Fan, R. Biophysical and biomolecular determination of cellular age in humans. Nat. Biomed. Eng. 2017, 1, 0093. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, B.K.; Berger, S.L.; Brunet, A.; Campisi, J.; Cuervo, A.M.; Epel, E.S.; Franceschi, C.; Lithgow, G.J.; Morimoto, R.I.; Pessin, J.E. Geroscience: Linking aging to chronic disease. Cell 2014, 159, 709–713. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.J.; Wijshake, T.; Tchkonia, T.; LeBrasseur, N.K.; Childs, B.G.; Van De Sluis, B.; Kirkland, J.L.; Van Deursen, J.M. Clearance of p16Ink4a-positive senescent cells delays ageing-associated disorders. Nature 2011, 479, 232–236. [Google Scholar] [CrossRef]
- Belsky, D.W.; Caspi, A.; Houts, R.; Cohen, H.J.; Corcoran, D.L.; Danese, A.; Harrington, H.; Israel, S.; Levine, M.E.; Schaefer, J.D. Quantification of biological aging in young adults. Proc. Natl. Acad. Sci. USA 2015, 112, E4104–E4110. [Google Scholar] [CrossRef]
- Rodier, F.; Campisi, J. Four faces of cellular senescence. J. Cell Biol. 2011, 192, 547–556. [Google Scholar] [CrossRef]


{kind=link}
| Class of Animal | Structure | Lethality | Method | Study Examples | Tradeoffs |
|---|---|---|---|---|---|
| Fish | Skeleton or majority of calcified body parts | Lethal | Band counts | Blackwell et al. (2016) and Korostelev et al. (2020) [27,28] | There may be errors when counting the bands and may underestimate the age. |
| Otolith | Lethal | Annuli counts | Snow et al. (2008) [58] | There may be errors when counting the annuli and may underestimate the age. | |
| Weight and length | Khan et al. (2018) [61] | Easy to operate, but not accurate enough. | |||
| Near-infrared spectroscopy (NIRS) | Healy et al. (2021) [68] | Spectral analysis has a higher cost, but it has good repeatability and higher efficiency. | |||
| Lead-radium dating | Andrews et al. (2011) [66] | Sample preprocessing needs time and effects, but it can relatively accurately obtain age information. | |||
| Lens | Lethal | Radiocarbon dating | Nielsen et al. (2016) and Boye et al. (2020) [47,49] | Regional variability can affect the accuracy of age estimates. | |
| Fin rays | Non-lethal | Annuli counts | Morehouse et al. (2013) [95] | There may be errors when counting the bands and may overestimate the age. | |
| Fin spines | Non-lethal | Annuli counts | Hedeholm et al. (2021) [51] | There may be errors when counting the bands and may underestimate the age. | |
| Near-infrared spectroscopy (NIRS) | Rigby et al. (2014) [98] | Spectral analysis has a higher cost, but it has good repeatability and higher efficiency. And mostly used to evaluate deep-sea cartilaginous fishes. | |||
| Scales | Non-lethal | Annuli counts | Ross et al. (2005) [57] | There may be errors when counting the annuli and may underestimate the age. | |
| Body length | Non-lethal | Growth model | Sajeevan and Kurup (2017) [6] | The standards need to be established, which requires the body length data from individuals of known ages and may underestimate the age. | |
| Amphibia | Skeleton | Non-lethal/Lethal | Skeletochronology | Ento et al. (2002) [153] | LAGs may overlap during counting, resulting in underestimation of age. |
| Reptilia | Skeleton | Non-lethal/Lethal | Skeletochronology | Comas et al. (2016) [154] | LAGs may overlap during counting, resulting in underestimation of age. |
| Scute | Non-lethal | Counting annual rings | Howell et al. (2018) [145] | Easy to operate, but not accurate enough. | |
| Avia | Blood | Non-lethal | DNA methylation (DNAm) | De Paoli-Iseppi et al. (2019) [1] | It has a high cost, and not universal among different species. |
| Voice | Non-lethal | - | Vaytina and Shitikov (2019) [148] | Only age classes can be judged, and not accurate enough. | |
| Physical characteristics | Non-lethal | - | Amiot et al. (2015) and Costa et al. (2020) [155,156] | Often used to determine the age of nestlings, and not accurate enough. | |
| Mammalia | Skeleton | Non-lethal/Lethal | - | Kryštufek et al. (2005) [33] | Mostly used in land mammals, and it may have a large error. |
| Lens | Lethal | Dry weight | McLeod et al. (2006) [38] | The standards need to be established, which requires the dry weight data from individuals of known ages. | |
| Aspartic acid racemization (AAR) | McLeod et al. (2006) and Boye et al. (2020) [38,51] | The samples need to be kept fresh, and the freshness of the sample may have an impact on the results. | |||
| Blood | Non-lethal | Hematologic and serum biochemical analyses | Rørtveit et al. (2015) [75] | Easy operation, but only age classes can be judged. | |
| DNAm | Horvath et al. (2022) [8] | It has a high cost, and not universal among different species. | |||
| Teeth | Non-lethal | Tooth replacement and wear (TRW) | Rosatte et al. (2007) [103] | The results may have some substantial errors. | |
| Growth layer groups (GLGs) | Rust et al. (2019) [101] | As age increases, the difficulty of calculating GLG often increases. | |||
| Cementum annuli analysis (CAA) | Asmus et al. (2011) [99] | There may be errors when counting the annuli and not accurate enough. | |||
| Gum-line recession | Fàbregas and Garcés-Narro (2014) [114] | Not accurate enough. | |||
| Size of pulp cavity | Gol’din et al. (2018) [106] | There may be errors in elderly individuals | |||
| Feces | Non-lethal | Size | Kongrit and Siripunkaw (2017) [123] | Easy to operate, but only age classes can be judged, and not accurate enough. | |
| Near-infrared spectroscopy (NIRS) | Wiedower et al. (2012) [7] | Spectral analysis has a higher cost, and only age classes can be judged. | |||
| Hair | Non-lethal | Hormone | Cattet et al. (2018) [150] | Hormone extraction costs are relatively high, and only specific hormones have a significant correlation with age. | |
| DNA methylation (DNAm) | Hao et al. (2021) [149] | It has a high cost, and not universal among different species. | |||
| Voice | Non-lethal | - | Stoeger et al. (2014) [147] | Only age classes can be judged, and not accurate enough. | |
| Physical characteristics | Non-lethal | - | Van Horn et al. (2015) [142] | Only age classes can be judged, and not accurate enough. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Bi, J.; Ning, Y.; Feng, J. Methodology Advances in Vertebrate Age Estimation. Animals 2024, 14, 343. https://doi.org/10.3390/ani14020343
Zhang Y, Bi J, Ning Y, Feng J. Methodology Advances in Vertebrate Age Estimation. Animals. 2024; 14(2):343. https://doi.org/10.3390/ani14020343
Chicago/Turabian StyleZhang, Yifei, Jinping Bi, Yao Ning, and Jiang Feng. 2024. "Methodology Advances in Vertebrate Age Estimation" Animals 14, no. 2: 343. https://doi.org/10.3390/ani14020343
APA StyleZhang, Y., Bi, J., Ning, Y., & Feng, J. (2024). Methodology Advances in Vertebrate Age Estimation. Animals, 14(2), 343. https://doi.org/10.3390/ani14020343