
Effects of Thyroid Powder on Tadpole (Lithobates catesbeiana) Metamorphosis and Growth: The Role of Lipid Metabolism and Gut Microbiota


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Diet Preparation
2.2. Trial Animals and Feeding Management
2.3. Sample Collection and Analyses
2.4. Statistical Analysis
3. Results
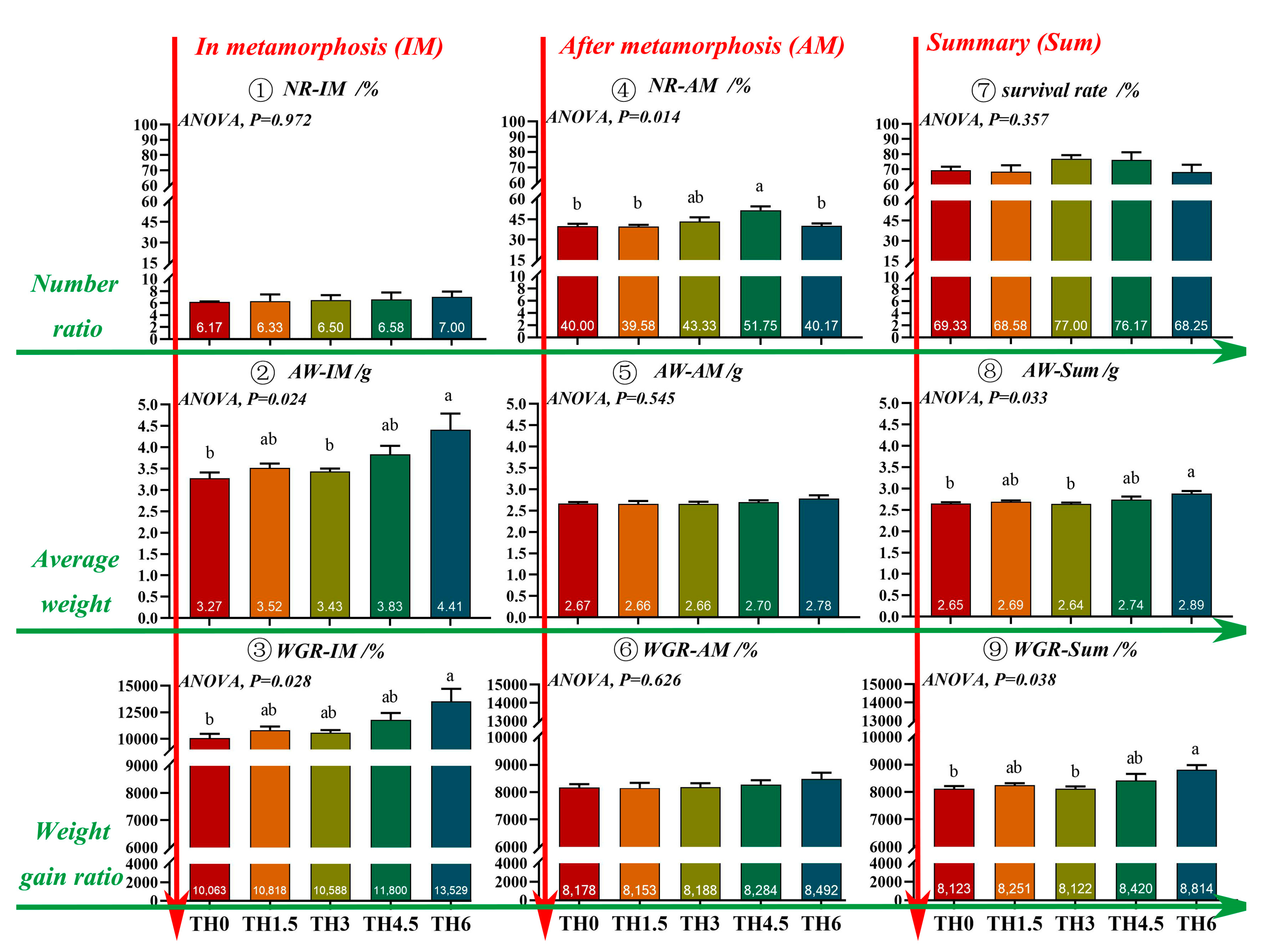
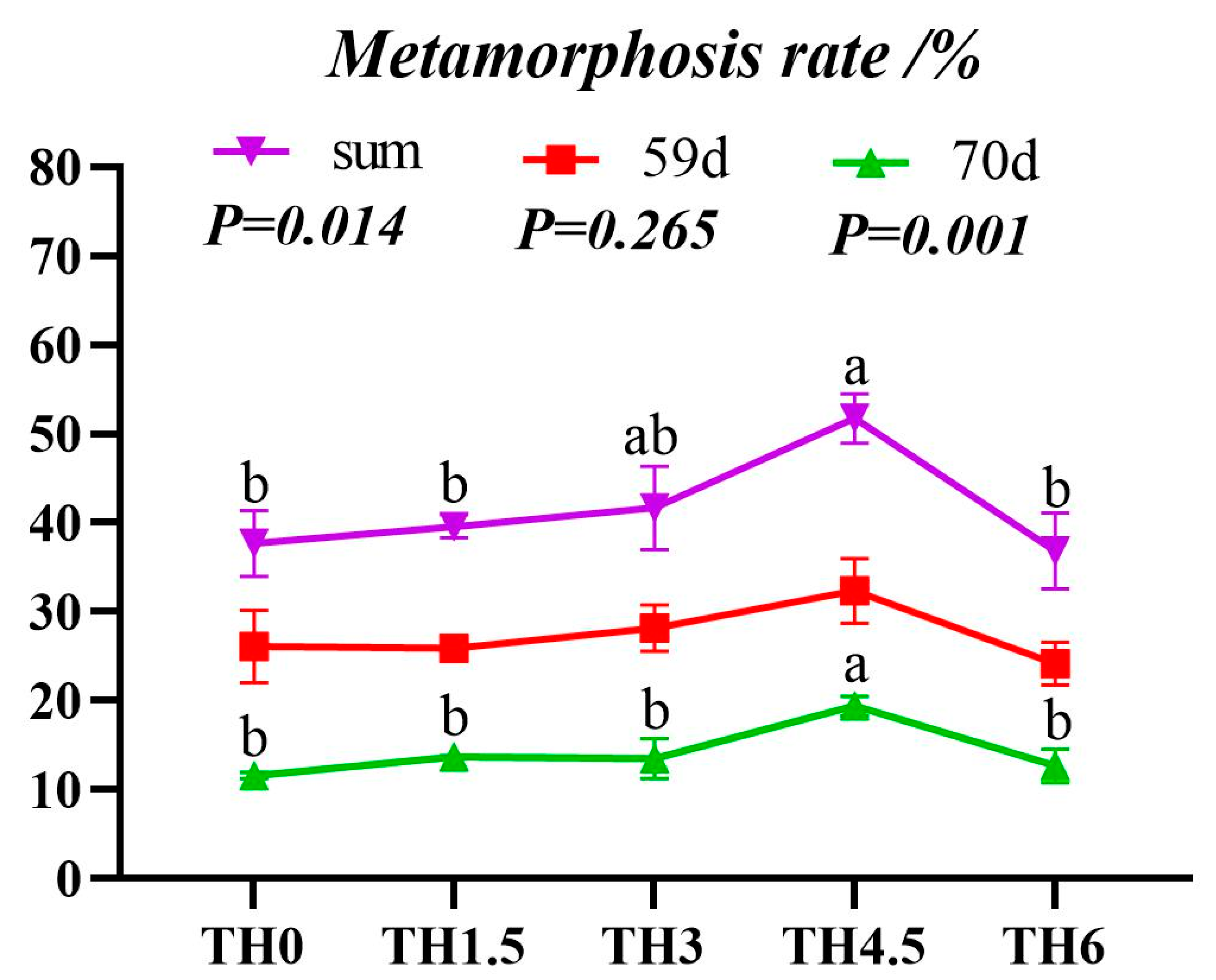
3.1. Growth and Metamorphosis
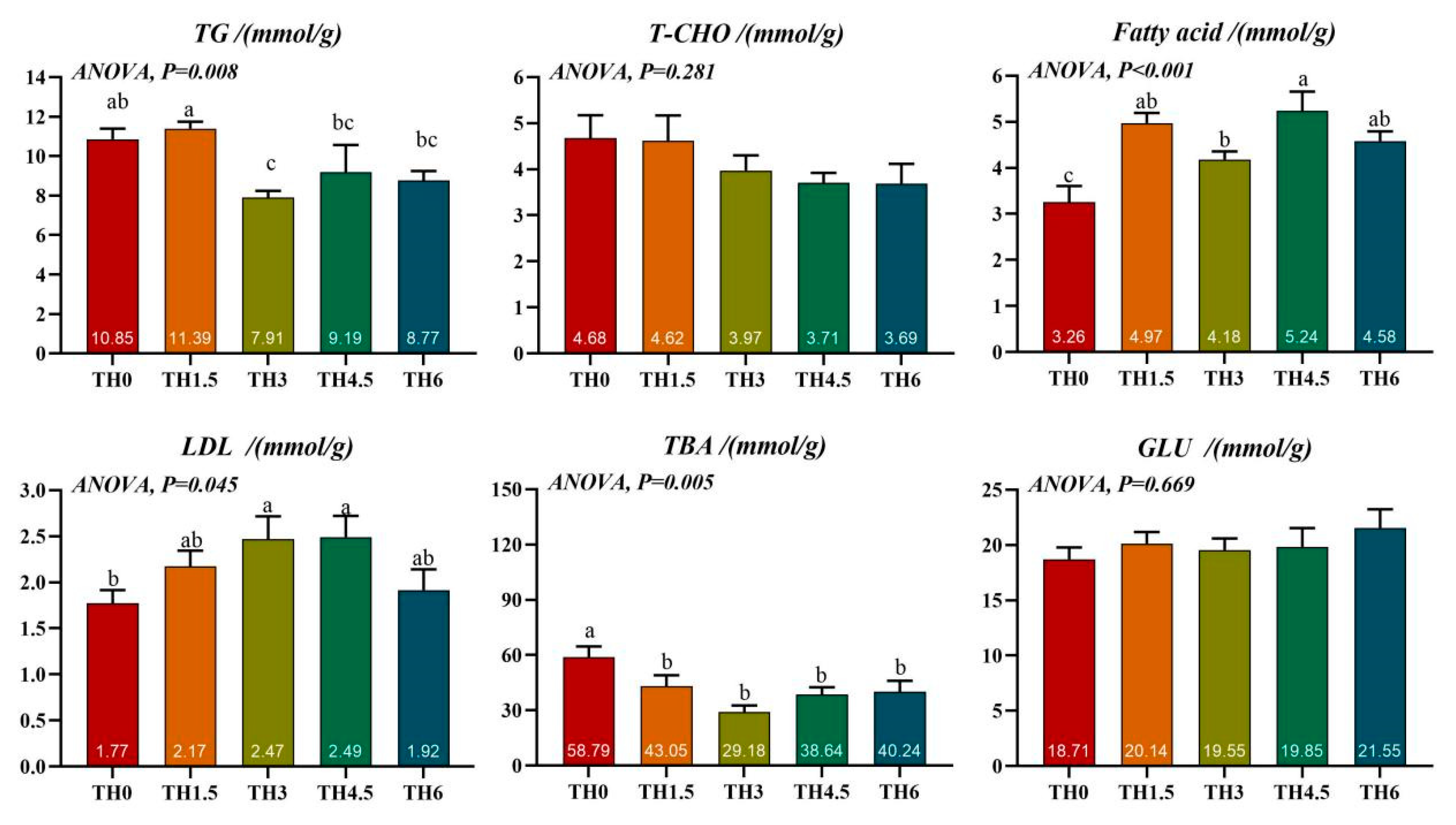
3.2. Liver Biochemical Tests
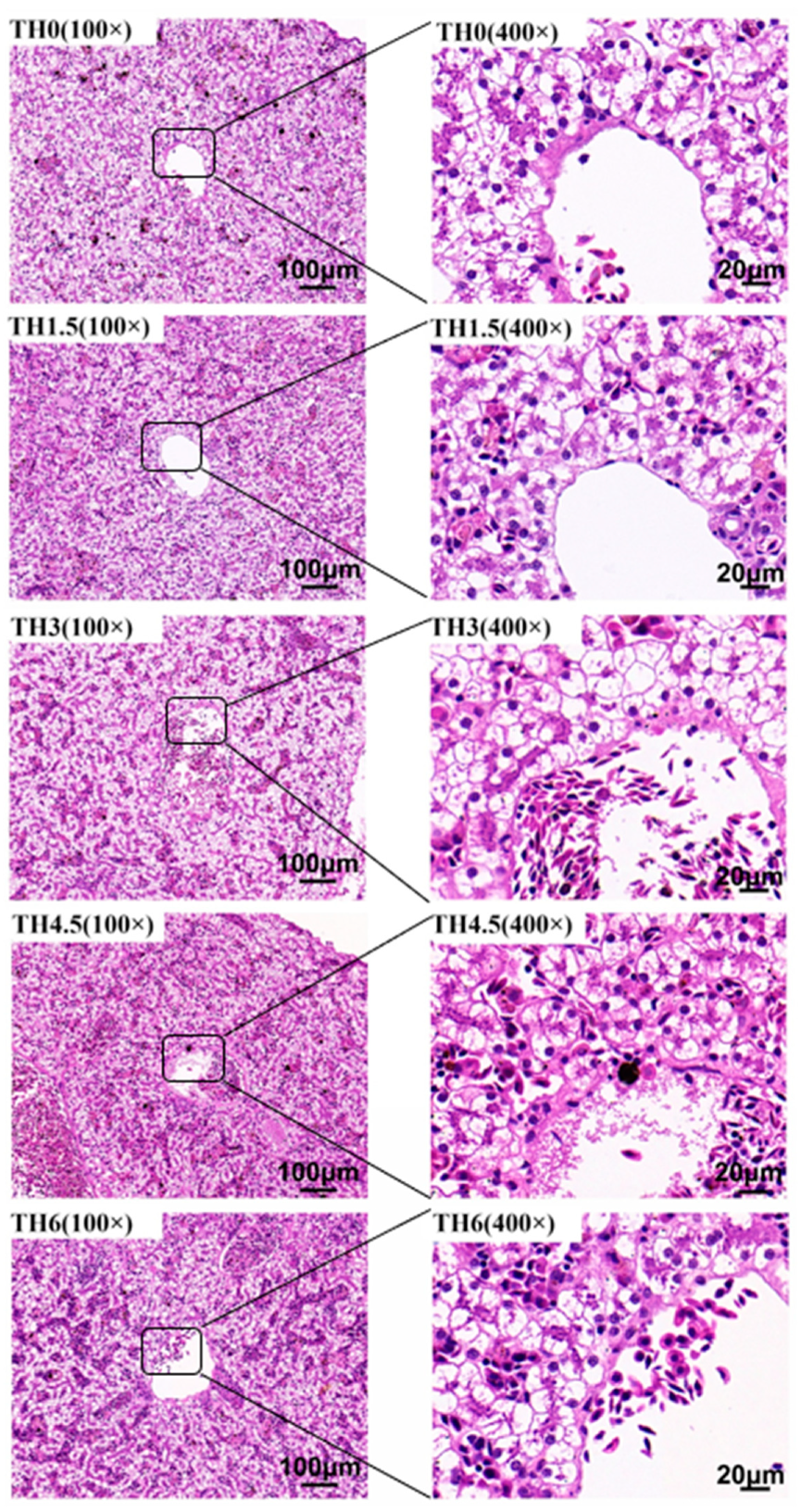
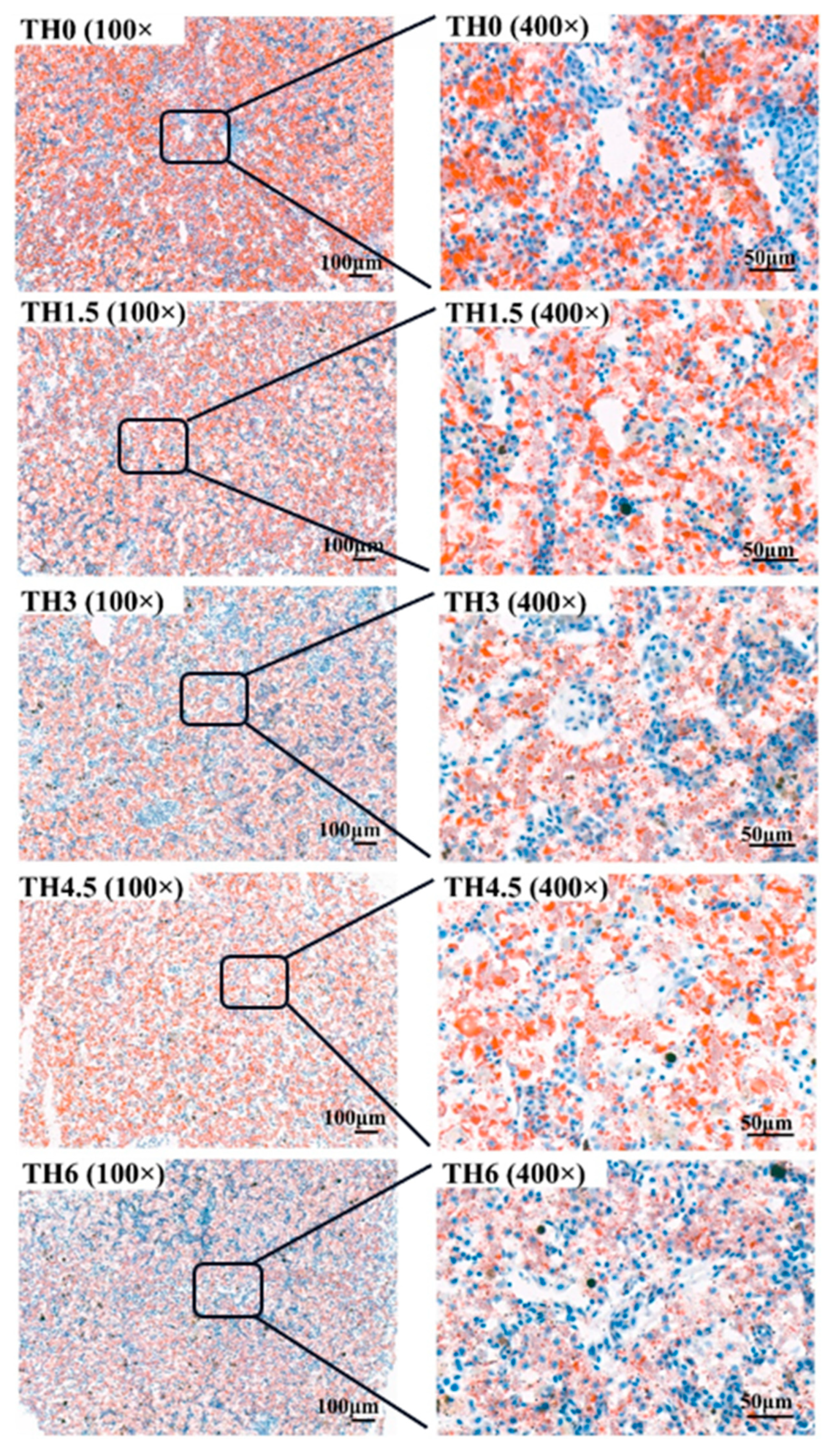
3.3. Liver HE Staining and Oil Red O Staining
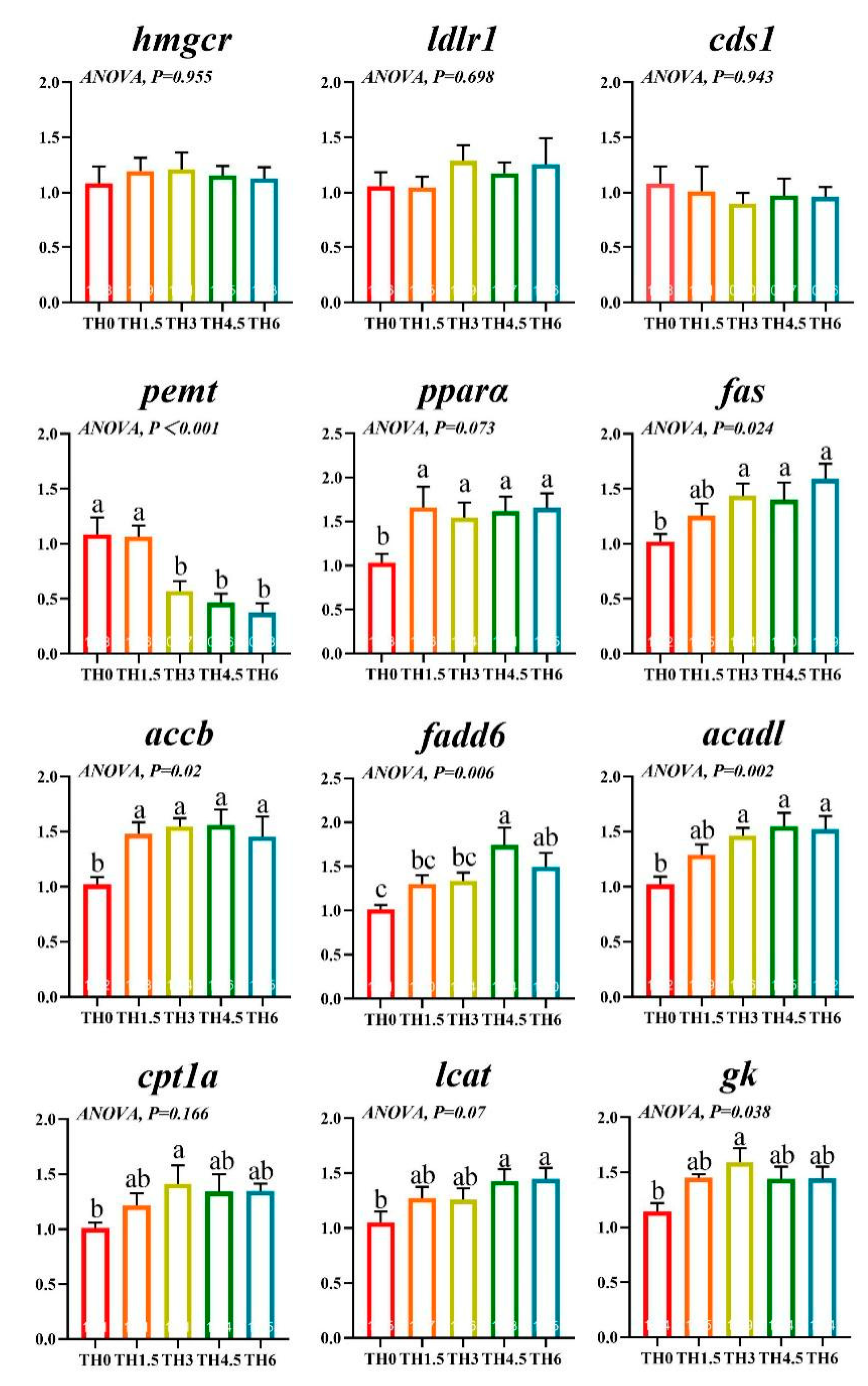
3.4. Lipid Metabolism-Related Gene Transcription Level
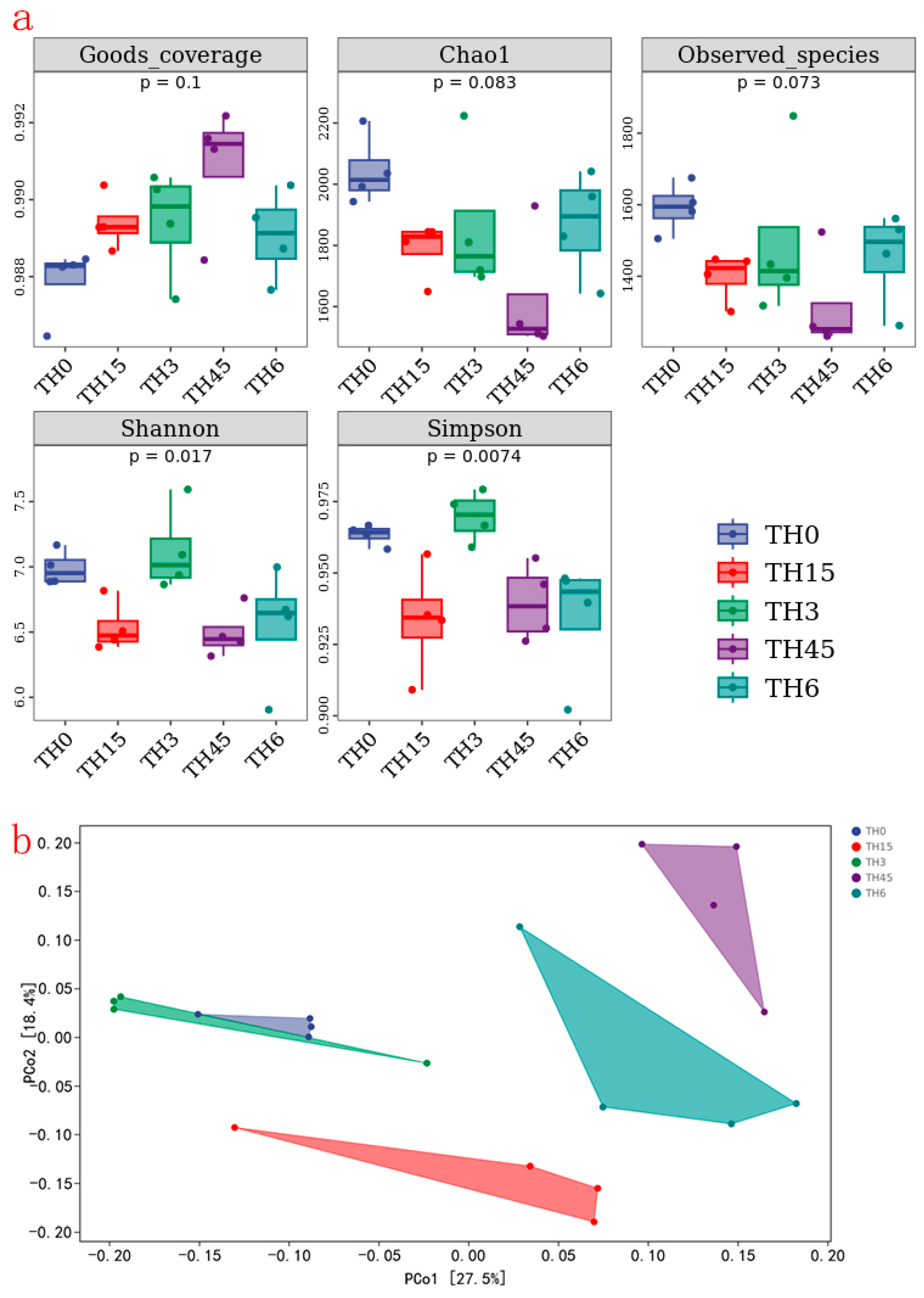
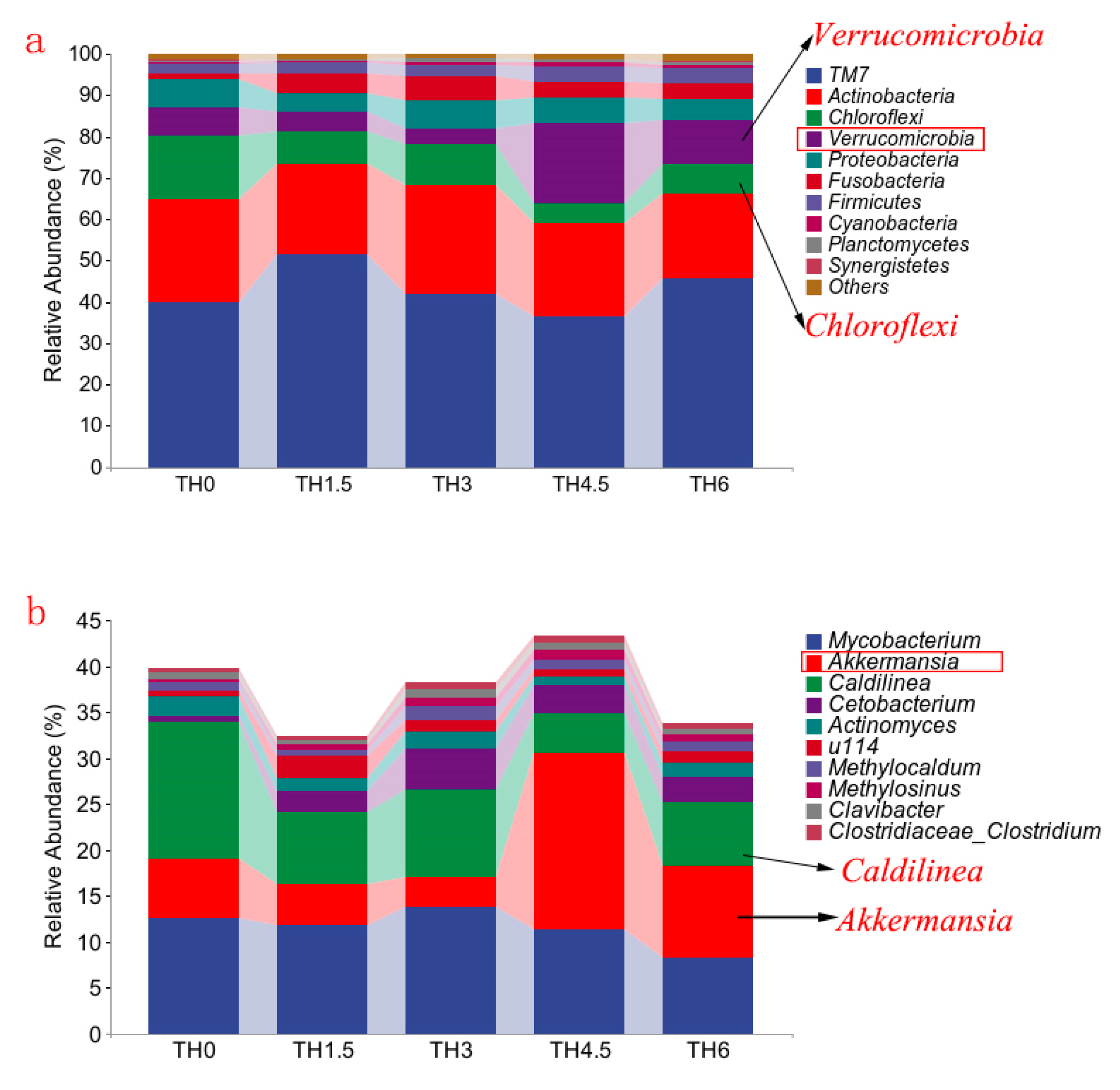
3.5. Gut Microbiota
4. Discussion
5. Conclusions
6. Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Funk, W.C.; Garcia, T.S.; Cortina, G.A.; Hill, R.H. Population genetics of introduced bullfrogs, Rana (Lithobates) catesbeianus, in the Willamette Valley, Oregon, USA. Biol. Invasions 2011, 13, 651–658. [Google Scholar] [CrossRef]
- Zhu, Y.L.; Bao, M.; Chen, C.; Yang, X.L.; Yan, W.L.; Ren, F.Z.; Wang, P.J.; Wen, P.C. Comparison of the Nutritional Composition of Bullfrog Meat from Different Parts of the Animal. Food Sci. Anim. Resour. 2021, 41, 1049–1059. [Google Scholar] [CrossRef] [PubMed]
- Niu, Z.; Xue, H.; Jiang, Z.; Chai, L.; Wang, H. Effects of temperature on metamorphosis and endochondral ossification in Rana chensinensis tadpoles, Comparative biochemistry and physiology. Part D Genom. Proteom. 2023, 45, 101057. [Google Scholar] [CrossRef]
- Wright, M.L.; Blanchard, L.S.; Jorey, S.T.; Basso, C.A.; Myers, Y.M.; Paquette, C.M. Metamorphic rate as a function of the light/dark cycle in Rana pipiens larvae. Comp. Biochem. Physiol. A Comp. Physiol. 1990, 96, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Bellakhal, M.; Neveu, A.; Fartouna-Bellakhal, M.; Missaoui, L. Aleya, Effects of temperature, density and food quality on larval growth and metamorphosis in the north African green frog Pelophylax saharicus. J. Therm. Biol. 2014, 45, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Ishizuya-Oka, A.; Shi, Y.-B. Molecular mechanisms for thyroid hormone-induced remodeling in the amphibian digestive tract: A model for studying organ regeneration. Dev. Growth Regen. 2005, 47, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Furlow, J.D.; Neff, E.S. A developmental switch induced by thyroid hormone: Xenopus laevis metamorphosis. Trends Endocrinol. Metab. TEM 2006, 17, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Tanizaki, Y.; Shibata, Y.; Zhang, H.; Shi, Y.B. Analysis of Thyroid Hormone Receptor α-Knockout Tadpoles Reveals That the Activation of Cell Cycle Program Is Involved in Thyroid Hormone-Induced Larval Epithelial Cell Death and Adult Intestinal Stem Cell Development During Xenopus tropicalis Metamorphosis. Thyroid 2021, 31, 128–142. [Google Scholar] [CrossRef]
- Nakajima, K.; Tazawa, I.; Yaoita, Y. Thyroid Hormone Receptor α- and β-Knockout Xenopus tropicalis Tadpoles Reveal Subtype-Specific Roles During Development. Endocrinology 2018, 159, 733–743. [Google Scholar] [CrossRef]
- Denver, R.J. The molecular basis of thyroid hormone-dependent central nervous system remodeling during amphibian metamorphosis, Comparative biochemistry and physiology. Part C Pharmacol. Toxicol. Endocrinol. 1998, 119, 219–228. [Google Scholar] [CrossRef]
- Koide, E.M.; Abbott, E.A.; Helbing, C.C. Uncovering early thyroid hormone signalling events through temperature-mediated activation of molecular memory in the cultured bullfrog tadpole tail fin. Gen. Comp. Endocrinol. 2022, 323, 114047. [Google Scholar] [CrossRef] [PubMed]
- Ashley, H.; Katti, P.; Frieden, E. Urea excretion in the bullrog tadpole: Effect of temperature, metamorphosis, and thyroid hormones. Dev. Biol. 1968, 17, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.A.; You, S.H.; Zhou, J.; Siddique, M.M.; Bay, B.H.; Zhu, X.; Privalsky, M.L.; Cheng, S.Y.; Stevens, R.D.; Summers, S.A.; et al. Thyroid hormone stimulates hepatic lipid catabolism via activation of autophagy. J. Clin. Investig. 2012, 122, 2428–2438. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.-P.; Zhou, J.; Zhang, J.; Xu, S.; Fu, G.; Dai, J.; Cai, M.; Hu, Y. Dietary enzymatic rice protein and enzymatic fish paste affect the growth, muscle development and quality traits of juvenile channel catfish (Ictalurus punctatus). Aquaculture 2022, 559, 738425. [Google Scholar] [CrossRef]
- Chakravarty, P.; Bordoloi, S.; Grosjean, S.; Ohler, A.; Borkotoki, A. Tadpole morphology and table of developmental stages of Polypedates teraiensis (Dubois, 1987). Alytes 2011, 27, 85–115. [Google Scholar]
- Feldman, A.T.; Wolfe, D. Tissue processing and hematoxylin and eosin staining. Methods Mol. Biol. 2014, 1180, 31–43. [Google Scholar] [CrossRef]
- Marquez, V.; Beccaria, A. Application of Oil Red O staining in oleaginous microalgae: Practical uses in screening and lipid quantification of different species. J. Appl. Phycol. 2020, 32, 1755–1761. [Google Scholar] [CrossRef]
- Peng, M.; Xue, J.; Hu, Y.; Wen, C.; Hu, B.; Jian, S.; Liang, L.; Yang, G. Disturbance in the homeostasis of intestinal microbiota by a high-fat diet in the rice field eel (Monopterus albus). Aquaculture 2019, 502, 347–355. [Google Scholar] [CrossRef]
- Ghasemi, A.; Zahediasl, S. Normality tests for statistical analysis: A guide for non-statisticians. Int. J. Endocrinol. Metab. 2012, 10, 486–489. [Google Scholar] [CrossRef]
- Shi, Y.B.; Wong, J.; Puzianowska-Kuznicka, M.; Stolow, M.A. Tadpole competence and tissue-specific temporal regulation of amphibian metamorphosis: Roles of thyroid hormone and its receptors. Bioessays 1996, 18, 391–399. [Google Scholar] [CrossRef]
- Sterner, Z.R.; Jabrah, A.; Shaidani, N.I.; Horb, M.E.; Dockery, R.; Paul, B.; Buchholz, D.R. Development and metamorphosis in frogs deficient in the thyroid hormone transporter MCT8. Gen. Comp. Endocrinol. 2023, 331, 114179. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Brown, D.D. Expression of type II iodothyronine deiodinase marks the time that a tissue responds to thyroid hormone-induced metamorphosis in Xenopus laevis. Dev. Biol. 2004, 266, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, A.; Baker, B.S.; Tata, J.R. Developmental and regional expression of thyroid hormone receptor genes during Xenopus metamorphosis. Development 1991, 112, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Thambirajah, A.A.; Koide, E.M.; Imbery, J.J.; Helbing, C.C. Contaminant and Environmental Influences on Thyroid Hormone Action in Amphibian Metamorphosis. Front. Endocrinol. 2019, 10, 276. [Google Scholar] [CrossRef] [PubMed]
- Raj, S.; Kyono, Y.; Sifuentes, C.J.; Arellanes-Licea, E.D.C.; Subramani, A.; Denver, R.J. Thyroid Hormone Induces DNA Demethylation in Xenopus Tadpole Brain. Endocrinology 2020, 161, bqaa155. [Google Scholar] [CrossRef] [PubMed]
- Cabello, G.; Wrutniak, C. Thyroid hormone and growth: Relationships with growth hormone effects and regulation. Reprod. Nutr. Dev. 1989, 29, 387–402. [Google Scholar] [CrossRef] [PubMed]
- Ross, I.; Omengan, D.B.; Huang, G.N.; Payumo, A.Y. Thyroid hormone-dependent regulation of metabolism and heart regeneration. J. Endocrinol. 2022, 252, R71–R82. [Google Scholar] [CrossRef] [PubMed]
- Wilsterman, K.; Buck, C.L.; Barnes, B.M.; Williams, C.T. Energy regulation in context: Free-living female arctic ground squirrels modulate the relationship between thyroid hormones and activity among life history stages. Horm. Behav. 2015, 75, 111–119. [Google Scholar] [CrossRef]
- Dearlove, D.J.; Hodson, L. Intrahepatic triglyceride content: Influence of metabolic and genetics drivers. Curr. Opin. Clin. Nutr. Metab. Care 2022, 25, 241–247. [Google Scholar] [CrossRef]
- Sifuentes, M.M.; Lechleiter, J.D. Thyroid Hormone Stimulation of Adult Brain Fatty Acid Oxidation. Vitam. Horm. 2018, 106, 163–193. [Google Scholar] [CrossRef]
- Giammanco, M.; Di Liegro, C.M.; Schiera, G.; Di Liegro, I. Genomic and Non-Genomic Mechanisms of Action of Thyroid Hormones and Their Catabolite 3,5-Diiodo-L-Thyronine in Mammals. Int. J. Mol. Sci. 2020, 21, 4140. [Google Scholar] [CrossRef] [PubMed]
- Gnocchi, D.; Ellis, E.C.S.; Johansson, H.; Eriksson, M.; Bruscalupi, G.; Steffensen, K.R.; Parini, P. Diiodothyronines regulate metabolic homeostasis in primary human hepatocytes by modulating mTORC1 and mTORC2 activity. Mol. Cell. Endocrinol. 2020, 499, 110604. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.A.; Singh, B.K.; Yen, P.M. Direct effects of thyroid hormones on hepatic lipid metabolism. Nat. Rev. Endocrinol. 2018, 14, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Klieverik, L.P.; Coomans, C.P.; Endert, E.; Sauerwein, H.P.; Havekes, L.M.; Voshol, P.J.; Rensen, P.C.; Romijn, J.A.; Kalsbeek, A.; Fliers, E. Thyroid hormone effects on whole-body energy homeostasis and tissue-specific fatty acid uptake in vivo. Endocrinology 2009, 150, 5639–5648. [Google Scholar] [CrossRef] [PubMed]
- Cicatiello, A.G.; Di Girolamo, D.; Dentice, M. Metabolic Effects of the Intracellular Regulation of Thyroid Hormone: Old Players, New Concepts. Front. Endocrinol. 2018, 9, 474. [Google Scholar] [CrossRef] [PubMed]
- Duntas, L.H.; Brenta, G. A Renewed Focus on the Association Between Thyroid Hormones and Lipid Metabolism. Front. Endocrinol. 2018, 9, 511. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, M.; Zhao, M.; Zhi, S.; Zhang, W.; Qu, L.; Xiong, J.; Yan, X.; Qin, C.; Nie, G.; et al. Dietary Bile Acid Supplementation Could Regulate the Glucose, Lipid Metabolism, and Microbiota of Common Carp (Cyprinus carpio L.) Fed with a High-Lipid Diet. Aquac. Nutr. 2023, 2023, 9953927. [Google Scholar] [CrossRef]
- Jia, M.; Zhang, Y.; Gao, Y.; Ma, X. Effects of Medium Chain Fatty Acids on Intestinal Health of Monogastric Animals. Curr. Protein Pept. Sci. 2020, 21, 777–784. [Google Scholar] [CrossRef]
- Wang, X.; Zeng, X.; Zhang, X.; Wei, J.; Zhang, Y.; Long, F.; Yue, T.; Yuan, Y. Aspergillus cristatus attenuates DSS-induced intestinal barrier damage through reducing the oxidative stress, regulating short-chain fatty acid and inhibiting MAPK signaling pathways. J. Sci. Food Agric. 2023, 103, 1736–1748. [Google Scholar] [CrossRef]
- Choudhuri, S.; Klaassen, C.D. Molecular Regulation of Bile Acid Homeostasis. Drug Metab. Dispos. 2022, 50, 425–455. [Google Scholar] [CrossRef]
- Zhu, B.P.; Zhou, J.C.; Wang, Z.Q.; Hu, Y.J.; Cai, M.L.; Yang, L.L.; Dai, J.H.; Hu, Y. Interactions between intestinal morphology, digestion, inflammatory responses, and gut microbiota of juvenile channel catfish elicited by dietary enzymatic rice protein. Fish Shellfish Immunol. 2022, 127, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Depommier, C.; Van Hul, M.; Everard, A.; Delzenne, N.M.; De Vos, W.M.; Cani, P.D. Pasteurized Akkermansia muciniphila increases whole-body energy expenditure and fecal energy excretion in diet-induced obese mice. Gut Microbes 2020, 11, 1231–1245. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, N.; Tan, H.Y.; Li, S.; Zhang, C.; Feng, Y. Function of Akkermansia muciniphila in Obesity: Interactions With Lipid Metabolism, Immune Response and Gut Systems. Front. Microbiol. 2020, 11, 219. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Lu, J.; Sun, L.; Lyu, X.; Chang, X.Y.; Mi, X.; Hu, M.G.; Wu, C.; Chen, X. Akkermansia muciniphila: A potential novel mechanism of nuciferine to improve hyperlipidemia. Biomed. Pharmacother. 2021, 133, 111014. [Google Scholar] [CrossRef] [PubMed]
- Platnich, J.M.; Muruve, D.A. NOD-like receptors and inflammasomes: A review of their canonical and non-canonical signaling pathways. Arch. Biochem. Biophys. 2019, 670, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, D. NOD1 and NOD2 Are Potential Therapeutic Targets for Cancer Immunotherapy. Comput. Intell. Neurosci. 2022, 2022, 2271788. [Google Scholar] [CrossRef]
- Hänninen, A.; Toivonen, R.; Pöysti, S.; Belzer, C.; Plovier, H.; Ouwerkerk, J.P.; Emani, R.; Cani, P.D.; De Vos, W.M. Akkermansia muciniphila induces gut microbiota remodelling and controls islet autoimmunity in NOD mice. Gut 2018, 67, 1445–1453. [Google Scholar] [CrossRef]
- Shi, Z.; Lei, H.; Chen, G.; Yuan, P.; Cao, Z.; Ser, H.L.; Zhu, X.; Wu, F.; Liu, C.; Dong, M.; et al. Impaired Intestinal Akkermansia muciniphila and Aryl Hydrocarbon Receptor Ligands Contribute to Nonalcoholic Fatty Liver Disease in Mice. mSystems 2021, 6, 10–1128. [Google Scholar] [CrossRef]
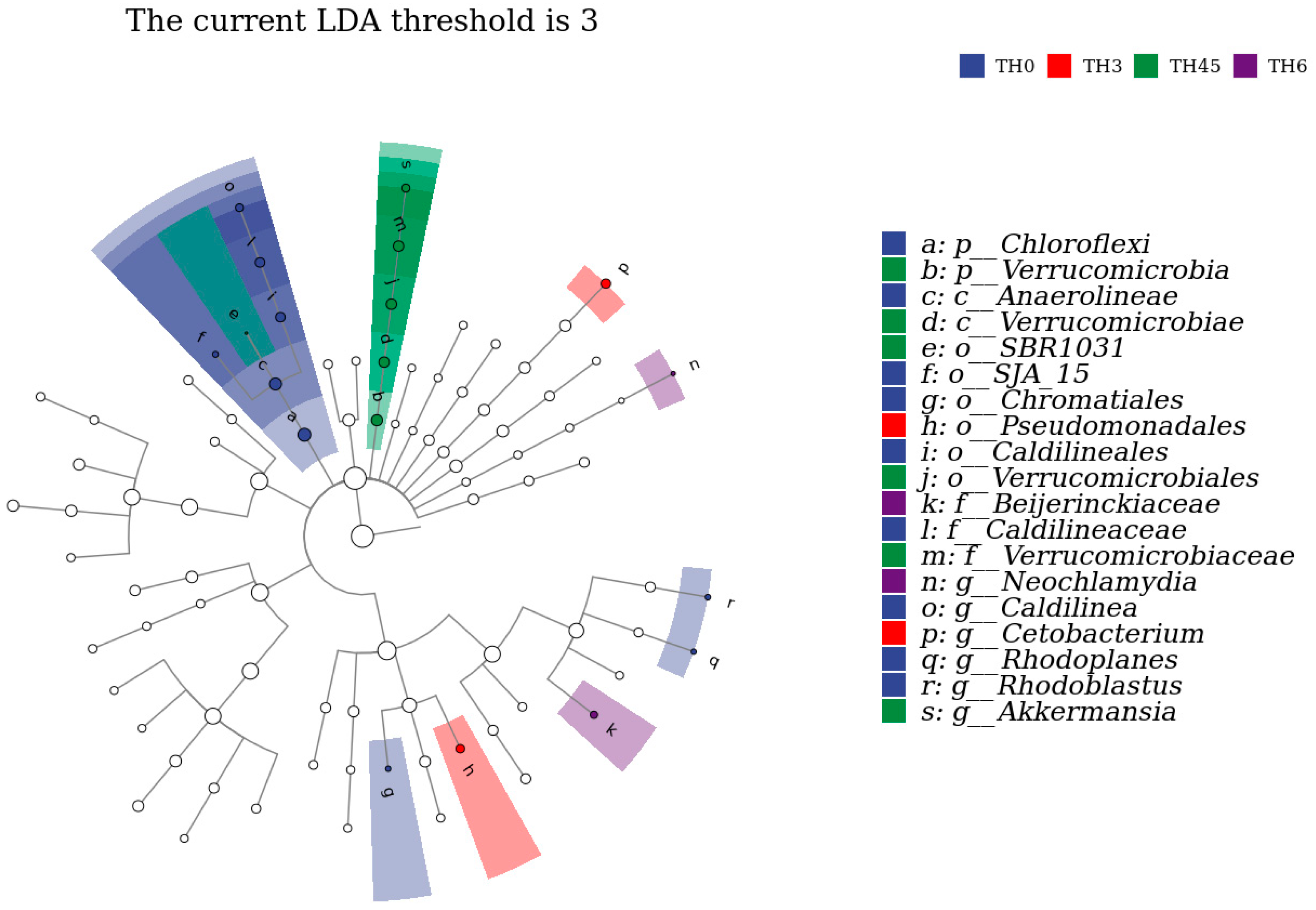
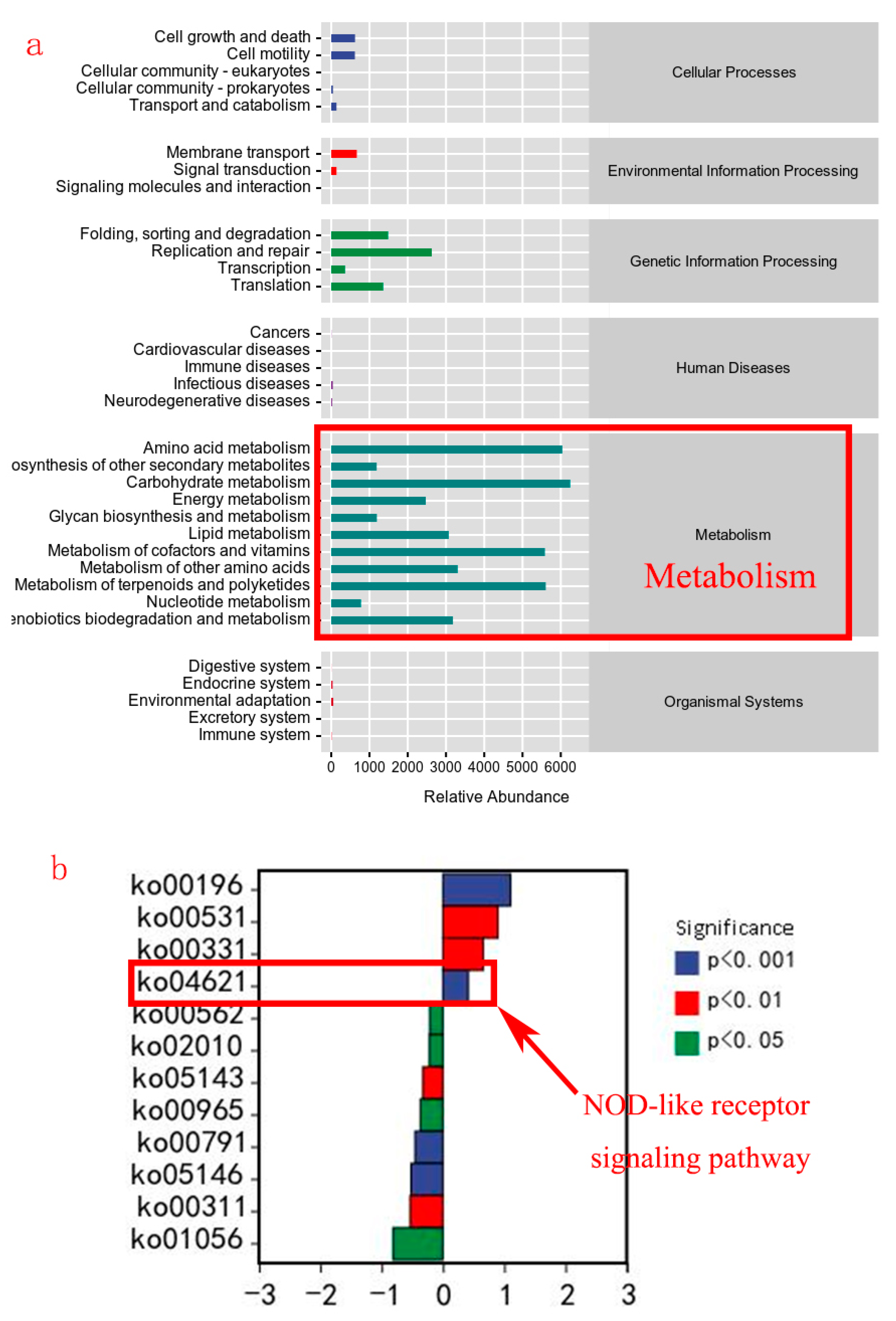

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | TH0 | TH1.5 | TH3 | TH4.5 | TH6 |
|---|---|---|---|---|---|
| Fish meal | 15.00 | 15.00 | 15.00 | 15.00 | 15.00 |
| Chicken powder | 20.00 | 20.00 | 20.00 | 20.00 | 20.00 |
| Soybean meal | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| Soy protein concentrate | 18.00 | 18.00 | 18.00 | 18.00 | 18.00 |
| Rice bran | 8.00 | 8.00 | 8.00 | 8.00 | 8.00 |
| Wheat flour | 23.95 | 23.80 | 23.65 | 23.50 | 23.35 |
| Thyroid powder (TH) | 0 | 0.15 | 0.30 | 0.45 | 0.60 |
| Soybean oil | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Vitamin and mineral premixes ※ | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 |
| Ca(H2PO4)2 | 1.50 | 1.50 | 1.50 | 1.50 | 1.50 |
| Choline chloride | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 |
| Ethoxyquin | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 |
| Mold inhibitor | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 |
| Proximate composition | |||||
| Dry matter | 92.31 | 92.31 | 92.31 | 92.31 | 92.31 |
| Crude protein | 43.42 | 43.42 | 43.42 | 43.42 | 43.42 |
| Crude lipid | 6.68 | 6.68 | 6.68 | 6.68 | 6.68 |
| Ash | 7.58 | 7.58 | 7.58 | 7.58 | 7.58 |
| Abbreviations | Explanation |
|---|---|
| acadl | acetyl-CoA carboxylase beta |
| accb | acetyl-CoA carboxylase beta |
| actb_g1 | actin beta/gamma 1 |
| cpt1a | carnitine palmitoyltransferase 1a |
| cds1 | CDP-diacylglycerol synthase |
| fas | fatty acid synthase |
| gk | glycerol kinase |
| hmgcr | 3-hydroxy-3-methylglutaryl-CoA reductase |
| lcat | lecithin-cholesterol acyltransferase |
| ldlr1 | low-density lipoprotein receptor 1 |
| pemt | phosphatidylethanolamine N-methyltransferase |
| fadd6 | fatty acid desaturase delta-6 |
| pparα | peroxisome proliferator-activated receptor alpha |
| TG | triglyceride |
| T-CHO | total cholesterol |
| LDL | low-density lipoprotein |
| TBA | total bile acid |
| GLU | glucose |
| TH | thyroid powder |
| Gene | Sequence, 5′→3′ | GenBank Number |
|---|---|---|
| acadl | Fwd: TGAGGAAACCCGGAACTATGTC Rev: TGTGCTGCACGGTCTGTAAGT | LH364687.1 |
| accb | Fwd: GTTAAAGCTGCCATCCTCACTGT Rev: TGTCCGTCTGGCTAAGATGGT | LH212450.1 |
| actb_g1 | Fwd: ATGATGCTCCTCGTGCTGTGT Rev: CCCCATTCCAACCATGACA | LH355272.1 |
| cpt1a | Fwd: TGATTGGCAAAATCAAAGAACATC Rev: AATGCTCTGACCCTGGTGAGA | LH022414.1 |
| cds1 | Fwd: GGTTTCTGCATGTTTGTGTTGAG Rev: TCCATCCAAACATGTAAAACTGAAG | LH114672.1 |
| fas | Fwd: CCTCCACGCCAGAACAAGAT Rev: GATATTTTTATGAGTGGACATTGTATCGA | LH228595.1 |
| gk | Fwd: AACGCTTTGAGCCACAGATTAAT Rev: CTGCTTTTTTCCATCGAGCAT | LH193866.1 |
| hmgcr | Fwd: TGCATCCTCAAAAACCCAGAT Rev: GGGATGTGTTTAGCATTCACCAA | LH363056.1 |
| lcat | Fwd: GCTGTAGGGTGACCTGTTCCAT Rev: AGATACGAAGGGCCTTCTGGAT | LH171224.1 |
| ldlr1 | Fwd: AAGGCTACCAACTAGATCCAGTAACTG Rev: CGGTTGGTGAAGAACAGGTATG | LH243159.1 |
| pemt | Fwd: CCGATATACGGTGACCCAAAA Rev: ACCCGCTCTTCTGGAATGTG | LH373164.1 |
| fadd6 | Fwd: TGGATCCTTGCTGAATATGTTAGG | LH144230.1 |
| Rev: AAGGGAGCTTCAGCCAACTG | ||
| pparα | Fwd: CCCGACATTCGATGTTTAGAGATT Rev: CCAGCCCATCTTCTATCACCTT | LH193621.1 |
| Initial Average Weight/g | Initial Total Weight/g | Final Total Weight/g | Total Weight Gain Ratio/% | Feed Conversion Ratio | |
|---|---|---|---|---|---|
| TH0 | 0.03 ± 0.00 | 12.88 ± 0.03 | 733.92 ± 15.65 | 5597.26 ± 128.36 | 1.18 ± 0.03 |
| TH1.5 | 0.03 ± 0.00 | 12.87 ± 0.06 | 738.13 ± 51.23 | 5630.70 ± 372.67 | 1.18 ± 0.08 |
| TH3 | 0.03 ± 0.00 | 12.83 ± 0.03 | 811.91 ± 15.87 | 6227.31 ± 137.21 | 1.06 ± 0.02 |
| TH4.5 | 0.03 ± 0.00 | 12.87 ± 0.03 | 834.37 ± 54.48 | 6384.41 ± 437.83 | 1.04 ± 0.07 |
| TH6 | 0.03 ± 0.00 | 12.95 ± 0.05 | 785.69 ± 37.73 | 5967.86 ± 301.60 | 1.10 ± 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, B.; Shao, C.; Xu, W.; Dai, J.; Fu, G.; Hu, Y. Effects of Thyroid Powder on Tadpole (Lithobates catesbeiana) Metamorphosis and Growth: The Role of Lipid Metabolism and Gut Microbiota. Animals 2024, 14, 208. https://doi.org/10.3390/ani14020208
Zhu B, Shao C, Xu W, Dai J, Fu G, Hu Y. Effects of Thyroid Powder on Tadpole (Lithobates catesbeiana) Metamorphosis and Growth: The Role of Lipid Metabolism and Gut Microbiota. Animals. 2024; 14(2):208. https://doi.org/10.3390/ani14020208
Chicago/Turabian StyleZhu, Bo, Chuang Shao, Wenjie Xu, Jihong Dai, Guihong Fu, and Yi Hu. 2024. "Effects of Thyroid Powder on Tadpole (Lithobates catesbeiana) Metamorphosis and Growth: The Role of Lipid Metabolism and Gut Microbiota" Animals 14, no. 2: 208. https://doi.org/10.3390/ani14020208
APA StyleZhu, B., Shao, C., Xu, W., Dai, J., Fu, G., & Hu, Y. (2024). Effects of Thyroid Powder on Tadpole (Lithobates catesbeiana) Metamorphosis and Growth: The Role of Lipid Metabolism and Gut Microbiota. Animals, 14(2), 208. https://doi.org/10.3390/ani14020208