
Chronic Inflammatory Enteropathy and Low-Grade Intestinal T-Cell Lymphoma Are Associated with Altered Microbial Tryptophan Catabolism in Cats
 ,
,  and
and
Abstract
:Simple Summary
Abstract
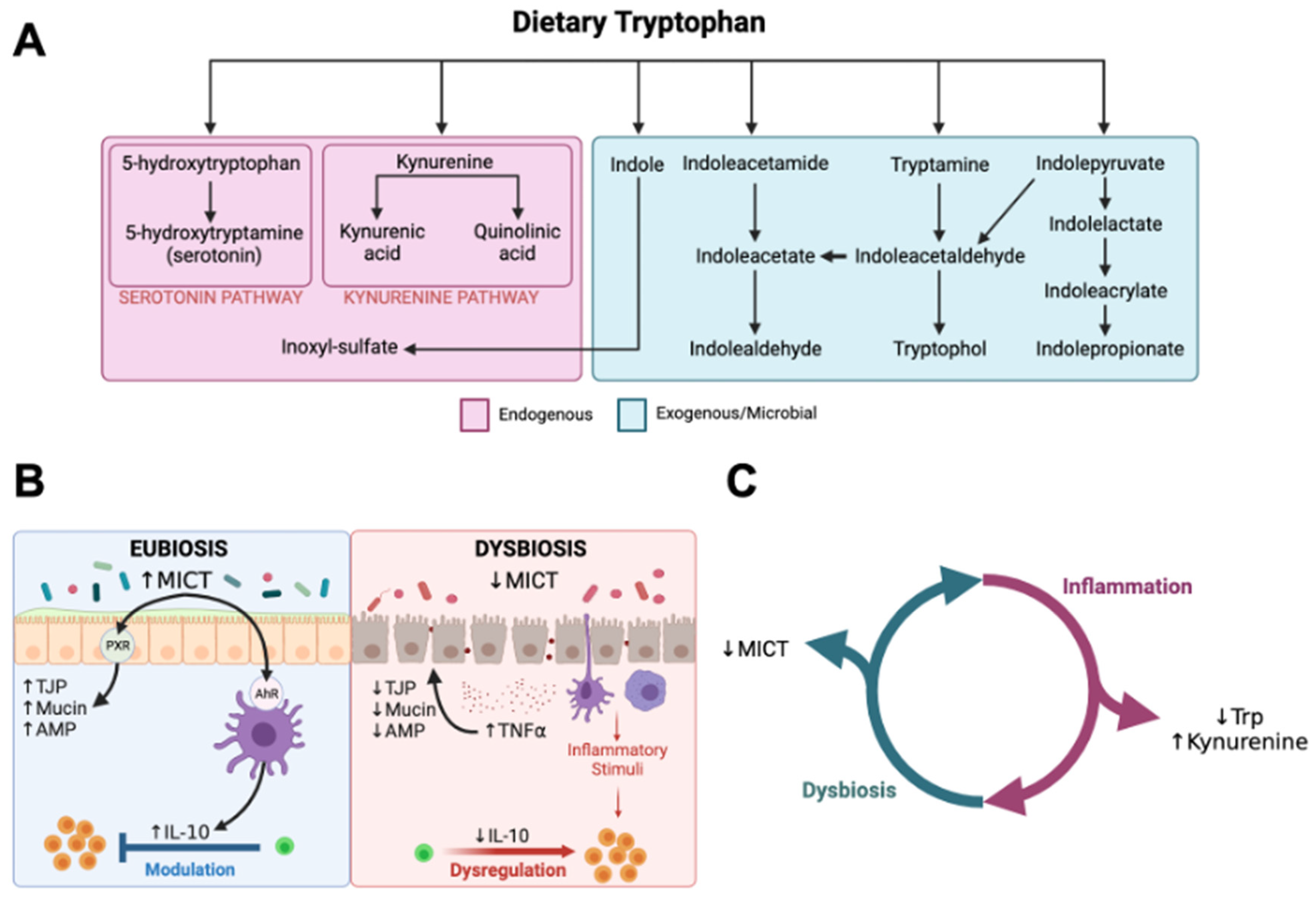
1. Introduction
2. Materials and Methods
2.1. Patient Population and Sample Acquisition
2.2. Quantification of Tryptophan Catabolites
2.3. Statistical Analysis
3. Results
3.1. Patient Population
3.2. Serum Concentrations of Tryptophan Catabolites
3.3. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moore, P.F.; Rodriguez-Bertos, A.; Kass, P.H. Feline Gastrointestinal Lymphoma: Mucosal Architecture, Immunophenotype, and Molecular Clonality. Vet. Pathol. 2012, 49, 658–668. [Google Scholar] [CrossRef]
- Moore, P.F.; Woo, J.C.; Vernau, W.; Kosten, S.; Graham, P.S. Characterization of feline T cell receptor gamma (TCRG) variable region genes for the molecular diagnosis of feline intestinal T cell lymphoma. Vet. Immunol. Immunopathol. 2005, 106, 167–178. [Google Scholar] [CrossRef]
- Yu, L.C.H. Microbiota dysbiosis and barrier dysfunction in inflammatory bowel disease and colorectal cancers: Exploring a common ground hypothesis. J. Biomed. Sci. 2018, 25, 79. [Google Scholar] [CrossRef]
- Hertli, S.; Zimmermann, P. Molecular interactions between the intestinal microbiota and the host. Mol. Microbiol. 2022, 117, 1297. [Google Scholar] [CrossRef]
- Quaglio, A.E.V.; Grillo, T.G.; De Oliveira, E.C.S.; Di Stasi, L.C.; Sassaki, L.Y. Gut microbiota, inflammatory bowel disease and colorectal cancer. World J. Gastroenterol. 2022, 28, 4053–4060. [Google Scholar] [CrossRef]
- Marsilio, S.; Pilla, R.; Sarawichitr, B.; Chow, B.; Hill, S.L.; Ackermann, M.R.; Estep, J.S.; Lidbury, J.A.; Steiner, J.M.; Suchodolski, J.S. Characterization of the fecal microbiome in cats with inflammatory bowel disease or alimentary small cell lymphoma. Sci. Rep. 2019, 9, 19208. [Google Scholar] [CrossRef]
- Sung, C.-H.; Marsilio, S.; Chow, B.; Zornow, K.A.; Slovak, J.E.; Pilla, R.; Lidbury, J.A.; Steiner, J.M.; Park, S.Y.; Hong, M.-P.; et al. Dysbiosis index to evaluate the fecal microbiota in healthy cats and cats with chronic enteropathies. J. Feline Med. Surg. 2022, 24, e1–e12. [Google Scholar] [CrossRef]
- Badawy, A.A.B. Kynurenine Pathway of Tryptophan Metabolism: Regulatory and Functional Aspects. Int. J. Tryptophan Res. 2017, 10, 1178646917691938. [Google Scholar] [CrossRef]
- Dehhaghi, M.; Kazemi Shariat Panahi, H.; Guillemin, G.J. Microorganisms, Tryptophan Metabolism, and Kynurenine Pathway: A Complex Interconnected Loop Influencing Human Health Status. Int. J. Tryptophan Res. 2019, 12, 1178646919852996. [Google Scholar] [CrossRef] [PubMed]
- Murakami, Y.; Saito, K. Species and cell types difference in tryptophan metabolism. Int. J. Tryptophan Res. 2013, 6, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Haq, S.; Grondin, J.A.; Khan, W.I. Tryptophan-derived serotonin-kynurenine balance in immune activation and intestinal inflammation. FASEB J. 2021, 35, e21888. [Google Scholar] [CrossRef]
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 2018, 9, 3294. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Z.H.; Zabed, H.M.; Yun, J.; Zhang, G.; Qi, X. An Insight into the Roles of Dietary Tryptophan and Its Metabolites in Intestinal Inflammation and Inflammatory Bowel Disease. Mol. Nutr. Food Res. 2021, 65, 2000461. [Google Scholar] [CrossRef]
- Fordtran, J.S.; Scroggie, W.B.; Polter, D.E. Colonic absorption of tryptophan metabolites in man. J. Lab. Clin. Med. 1964, 64, 125–132. [Google Scholar]
- Agus, A.; Planchais, J.; Sokol, H. Cell Host & Microbe Review Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef]
- Jin, U.-H.; Lee, S.-O.; Sridharan, G.; Lee, K.; Davidson, L.A.; Jayaraman, A.; Chapkin, R.S.; Alaniz, R.; Safe, S. Microbiome-derived tryptophan metabolites and their aryl hydrocarbon receptor-dependent agonist and antagonist activities. Mol. Pharmacol. 2014, 85, 777–788. [Google Scholar] [CrossRef]
- Wikoff, W.R.; Anfora, A.T.; Liu, J.; Schultz, P.G.; Lesley, S.A.; Peters, E.C.; Siuzdak, G. Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites. Proc. Natl. Acad. Sci. USA 2009, 106, 3698. [Google Scholar] [CrossRef]
- Bhattarai, Y.; Williams, B.B.; Battaglioli, E.J.; Whitaker, W.R.; Till, L.; Grover, M.; Linden, D.R.; Akiba, Y.; Kandimalla, K.K.; Zachos, N.C.; et al. Gut Microbiota-Produced Tryptamine Activates an Epithelial G-Protein-Coupled Receptor to Increase Colonic Secretion. Cell Host Microbe 2018, 23, 775–785.e5. [Google Scholar] [CrossRef]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef]
- Scott, S.A.; Fu, J.; Chang, P.V. Microbial tryptophan metabolites regulate gut barrier function via the aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. USA 2020, 117, 19376–19387. [Google Scholar] [CrossRef]
- Gupta, N.K.; Thaker, A.I.; Kanuri, N.; Riehl, T.E.; Rowley, C.W.; Stenson, W.F.; Ciorba, M.A. Serum analysis of tryptophan catabolism pathway: Correlation with Crohn’s disease activity. Inflamm. Bowel Dis. 2012, 18, 1214–1220. [Google Scholar] [CrossRef]
- Nikolaus, S.; Schulte, B.; Al-Massad, N.; Thieme, F.; Schulte, D.M.; Bethge, J.; Rehman, A.; Tran, F.; Aden, K.; Häsler, R.; et al. Increased Tryptophan Metabolism Is Associated With Activity of Inflammatory Bowel Diseases. Gastroenterology 2017, 153, 1504–1516.e2. [Google Scholar] [CrossRef]
- Sofia, M.A.; Ciorba, M.A.; Meckel, K.; Lim, C.K.; Guillemin, G.J.; Weber, C.R.; Bissonnette, M.; Pekow, J.R. Tryptophan Metabolism through the Kynurenine Pathway is Associated with Endoscopic Inflammation in Ulcerative Colitis. Inflamm. Bowel Dis. 2018, 24, 1471–1480. [Google Scholar] [CrossRef]
- Alexeev, E.E.; Lanis, J.M.; Kao, D.J.; Campbell, E.L.; Kelly, C.J.; Battista, K.D.; Gerich, M.E.; Jenkins, B.R.; Walk, S.T.; Kominsky, D.J.; et al. Microbiota-Derived Indole Metabolites Promote Human and Murine Intestinal Homeostasis through Regulation of Interleukin-10 Receptor. Am. J. Pathol. 2018, 188, 1183–1194. [Google Scholar] [CrossRef]
- Lamas, B.; Richard, M.L.; Leducq, V.; Pham, H.-P.; Michel, M.-L.; Da Costa, G.; Bridonneau, C.; Jegou, S.; Hoffmann, T.W.; Natividad, J.M.; et al. CARD9 impacts colitis by altering gut microbiota metabolism of tryptophan into aryl hydrocarbon receptor ligands. Nat. Med. 2016, 22, 598–605. [Google Scholar] [CrossRef]
- Vyhlídalová, B.; Krasulová, K.; Pečinková, P.; Marcalíková, A.; Vrzal, R.; Zemánková, L.; Vančo, J.; Trávníček, Z.; Vondráček, J.; Karasová, M.; et al. Gut microbial catabolites of tryptophan are ligands and agonists of the aryl hydrocarbon receptor: A detailed characterization. Int. J. Mol. Sci. 2020, 21, 2614. [Google Scholar] [CrossRef]
- Bansal, T.; Alaniz, R.C.; Wood, T.K.; Jayaraman, A. The bacterial signal indole increases epithelial-cell tight-junction resistance and attenuates indicators of inflammation. Proc. Natl. Acad. Sci. USA 2010, 107, 228–233. [Google Scholar] [CrossRef]
- Wlodarska, M.; Luo, C.; Kolde, R.; D’hennezel, E.; Annand, J.W.; Heim, C.E.; Krastel, P.; Schmitt, E.K.; Omar, A.S.; Creasey, E.A.; et al. Indoleacrylic Acid Produced by Commensal Peptostreptococcus Species Suppresses Inflammation. Cell Host Microbe 2017, 22, 25–37.e6. [Google Scholar] [CrossRef]
- Ehrlich, A.M.; Pacheco, A.R.; Henrick, B.M.; Taft, D.; Xu, G.; Huda, M.N.; Mishchuk, D.; Goodson, M.L.; Slupsky, C.; Barile, D.; et al. Indole-3-lactic acid associated with Bifidobacterium-dominated microbiota significantly decreases inflammation in intestinal epithelial cells. BMC Microbiol. 2020, 20, 357. [Google Scholar] [CrossRef]
- Vyhlídalová, B.; Bartoňková, I.; Jiskrová, E.; Li, H.; Mani, S.; Dvořák, Z. Differential activation of human pregnane X receptor PXR by isomeric mono-methylated indoles in intestinal and hepatic in vitro models. Toxicol. Lett. 2020, 324, 104–110. [Google Scholar] [CrossRef]
- Descamps, H.C.; Herrmann, B.; Wiredu, D.; Thaiss, C.A. The path toward using microbial metabolites as therapies. EBioMedicine 2019, 44, 747–754. [Google Scholar] [CrossRef]
- Pernomian, L.; Duarte-Silva, M.; de Barros Cardoso, C.R. The Aryl Hydrocarbon Receptor (AHR) as a Potential Target for the Control of Intestinal Inflammation: Insights from an Immune and Bacteria Sensor Receptor. Clin. Rev. Allergy Immunol. 2020, 59, 382–390. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, S.; Ma, N.; Johnston, L.J.; Wu, C.; Ma, X. Metabolites of microbiota response to tryptophan and intestinal mucosal immunity: A therapeutic target to control intestinal inflammation. Med. Res. Rev. 2021, 41, 1061–1088. [Google Scholar] [CrossRef]
- Barko, P.; Wu, Y.-A.; Steiner, J.; Norsworthy, G.; Gal, A.; Williams, D.; Estep, S. Altered Tryptophan Metabolism in Cats with Chronic Inflammatory Enteropathy or Alimentary Small Cell Lymphoma. Research Communications of the 32nd ECVIM-CA Online Congress. J. Vet. Intern. Med. 2022, 36, 2455–2551. [Google Scholar] [CrossRef]
- Briscoe, K.; Krockenberger, M.; Beatty, J.; Crowley, A.; Dennis, M.; Canfield, P.; Dhand, N.; Lingard, A.; Barrs, V. Histopathological and Immunohistochemical Evaluation of 53 Cases of Feline Lymphoplasmacytic Enteritis and Low-Grade Alimentary Lymphoma. J. Comp. Pathol. 2011, 145, 187–198. [Google Scholar] [CrossRef]
- Langerak, A.W.; Groenen, P.J.T.A.; Brüggemann, M.; Beldjord, K.; Bellan, C.; Bonello, L.; Boone, E.; I Carter, G.; Catherwood, M.; Davi, F.; et al. EuroClonality/BIOMED-2 guidelines for interpretation and reporting of Ig/TCR clonality testing in suspected lymphoproliferations. Leukemia 2012, 26, 2159–2171. [Google Scholar] [CrossRef]
- Paulin, M.V.; Couronné, L.; Beguin, J.; Le Poder, S.; Delverdier, M.; Semin, M.-O.; Bruneau, J.; Cerf-Bensussan, N.; Malamut, G.; Cellier, C.; et al. Feline low-grade alimentary lymphoma: An emerging entity and a potential animal model for human disease. BMC Vet. Res. 2018, 14, 306. [Google Scholar] [CrossRef]
- Marsilio, S. Differentiating Inflammatory Bowel Disease from Alimentary Lymphoma in Cats: Does It Matter? Vet. Clin. North. Am. Small Anim. Pract. 2021, 51, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Marsilio, S.; Newman, S.J.; Estep, J.S.; Giaretta, P.R.; Lidbury, J.A.; Warry, E.; Flory, A.; Morley, P.S.; Smoot, K.; Seeley, E.H.; et al. Differentiation of lymphocytic-plasmacytic enteropathy and small cell lymphoma in cats using histology-guided mass spectrometry. J. Vet. Intern. Med. 2020, 34, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Freiche, V.; Paulin, M.V.; Cordonnier, N.; Huet, H.; Turba, M.; Macintyre, E.; Molina, T.; Hermine, O.; Couronné, L.; Bruneau, J. Histopathologic, phenotypic, and molecular criteria to discriminate low-grade intestinal T-cell lymphoma in cats from lymphoplasmacytic enteritis. J. Vet. Intern. Med. 2021, 35, 2673–2684. [Google Scholar] [CrossRef] [PubMed]
- Marsilio, S.; Freiche, V.; Johnson, E.; Leo, C.; Langerak, A.W.; Peters, I.; Ackermann, M.R. ACVIM consensus statement guidelines on diagnosing and distinguishing low-grade neoplastic from inflammatory lymphocytic chronic enteropathies in cats. J. Vet. Intern. Med. 2023, 37, 794–816. [Google Scholar] [CrossRef] [PubMed]
- Laflamme, D. Development and validation of a body condition score system for cats: A clinical tool. Feline Pract. 1997, 5, 13–18. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Sakai, K.; Maeda, S.; Yonezawa, T.; Matsuki, N. Decreased plasma amino acid concentrations in cats with chronic gastrointestinal diseases and their possible contribution in the inflammatory response. Vet. Immunol. Immunopathol. 2018, 195, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, I.; Rizzo, A.; Sarra, M.; Sica, G.; Sileri, P.; Biancone, L.; Macdonald, T.T.; Pallone, F.; Monteleone, G. Aryl hydrocarbon receptor-induced signals up-regulate IL-22 production and inhibit inflammation in the gastrointestinal tract. Gastroenterology 2011, 141, 237–248.e1. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Perdew, G.H. The aryl hydrocarbon receptor as a mediator of host-microbiota interplay. Gut Microbes 2020, 12, 1859812. [Google Scholar] [CrossRef] [PubMed]
- Reigstad, C.S.; Salmonson, C.E.; Rainey, J.F., III; Szurszewski, J.H.; Linden, D.R.; Sonnenburg, J.L.; Farrugia, G.; Kashyap, P.C. Gut microbes promote colonic serotonin production through an effect of short-chain fatty acids on enterochromaffin cells. FASEB J. 2015, 29, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- Marsilio, S.; Chow, B.; Hill, S.L.; Ackermann, M.R.; Estep, J.S.; Sarawichitr, B.; Pilla, R.; Lidbury, J.A.; Steiner, J.M.; Suchodolski, J.S. Untargeted metabolomic analysis in cats with naturally occurring inflammatory bowel disease and alimentary small cell lymphoma. Sci. Rep. 2021, 11, 9198. [Google Scholar] [CrossRef]
- Blake, A.B.; Guard, B.C.; Honneffer, J.B.; Lidbury, J.A.; Steiner, J.M.; Suchodolski, J.S. Altered microbiota, fecal lactate, and fecal bile acids in dogs with gastrointestinal disease. PLoS ONE 2019, 14, e0224454. [Google Scholar] [CrossRef]
- Mawe, G.M.; Hoffman, J.M. Serotonin Signaling in the Gastrointestinal Tract: Functions, dysfunctions, and therapeutic targets. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 473–486. [Google Scholar] [CrossRef]
- Marsilio, S.; Ackermann, M.R.; Lidbury, J.A.; Suchodolski, J.S.; Steiner, J.M. Results of histopathology, immunohistochemistry, and molecular clonality testing of small intestinal biopsy specimens from clinically healthy client-owned cats. J. Vet. Intern. Med. 2019, 33, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.H.; Jha, A.R.; Do, S.; Scarsella, E.; Shmalberg, J.; Schauwecker, A.; Steelman, A.J.; Honaker, R.W.; Swanson, K.S. Dietary enrichment of resistant starches or fibers differentially alter the feline fecal microbiome and metabolite profile. Anim. Microbiome 2022, 4, 61. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Healthy (n = 26) | CIE (n = 44) | LGITL (n = 31) | p-Value | |
|---|---|---|---|---|
| Age (years) | 0.010 | |||
| Median | 10.0 a | 10.5 a,b | 12.0 b | |
| IQR | 8.0–11.0 | 7.0–13.0 | 8.0–13.0 | |
| Min–Max | 3–14 | 2–17 | 7–15 | |
| Sex | 0.85 | |||
| Spayed Female | 15 (57.7%) | 23 (52.3%) | 18 (58.1%) | |
| Neutered Male | 11 (42.3%) | 21 (47.7%) | 13 (41.9%) | |
| BCS (1–9) | <0.001 | |||
| Median | 6 a | 5 b | 4 b | |
| IQR | 5–8 | 4–5 | 4–5 | |
| Min–Max | 5–9 | 3–7 | 1–9 | |
| Cobalamin (ng/L) | <0.001 | |||
| Median | 1000.0 a | 743.0 b | 615.0 b | |
| IQR | 826.3–1000.0 | 378.8–891.3 | 209.0–906.5 | |
| Min–Max | 311.0–1000.0 | 150.0–1000.0 | 150.0–1000.0 | |
| Folate (µg/L) | 0.66 | |||
| Median | 15.8 | 16.1 | 16.0 | |
| IQR | 14.1–17.9 | 10.7–19.7 | 13.2–21.2 | |
| Min–Max | 11.0–21.5 | 5.2–38.0 | 9.2–48.0 | |
| fPLI (µg/L) | 0.045 | |||
| Median | 2.0 a | 2.0 a | 2.7 a | |
| IQR | 1.4–2.5 | 1.3–2.98 | 1.5–5.98 | |
| Min–Max | 0.9–3.2 | 0.5–21.6 | 1.0–51.0 | |
| fTLI (µg/L) | <0.001 | |||
| Median | 32.6 a | 32.2 a | 56.8 b | |
| IQR | 23.1–39.0 | 27.8–47.5 | 38.5–84.4 | |
| Min–Max | 15.9–63.7 | 14.3–231.0 | 15.4–243.7 | |
| Healthy (n = 26) | CIE (n = 44) | LGITL (n = 31) | Kruskal–Wallis p-Value | |
|---|---|---|---|---|
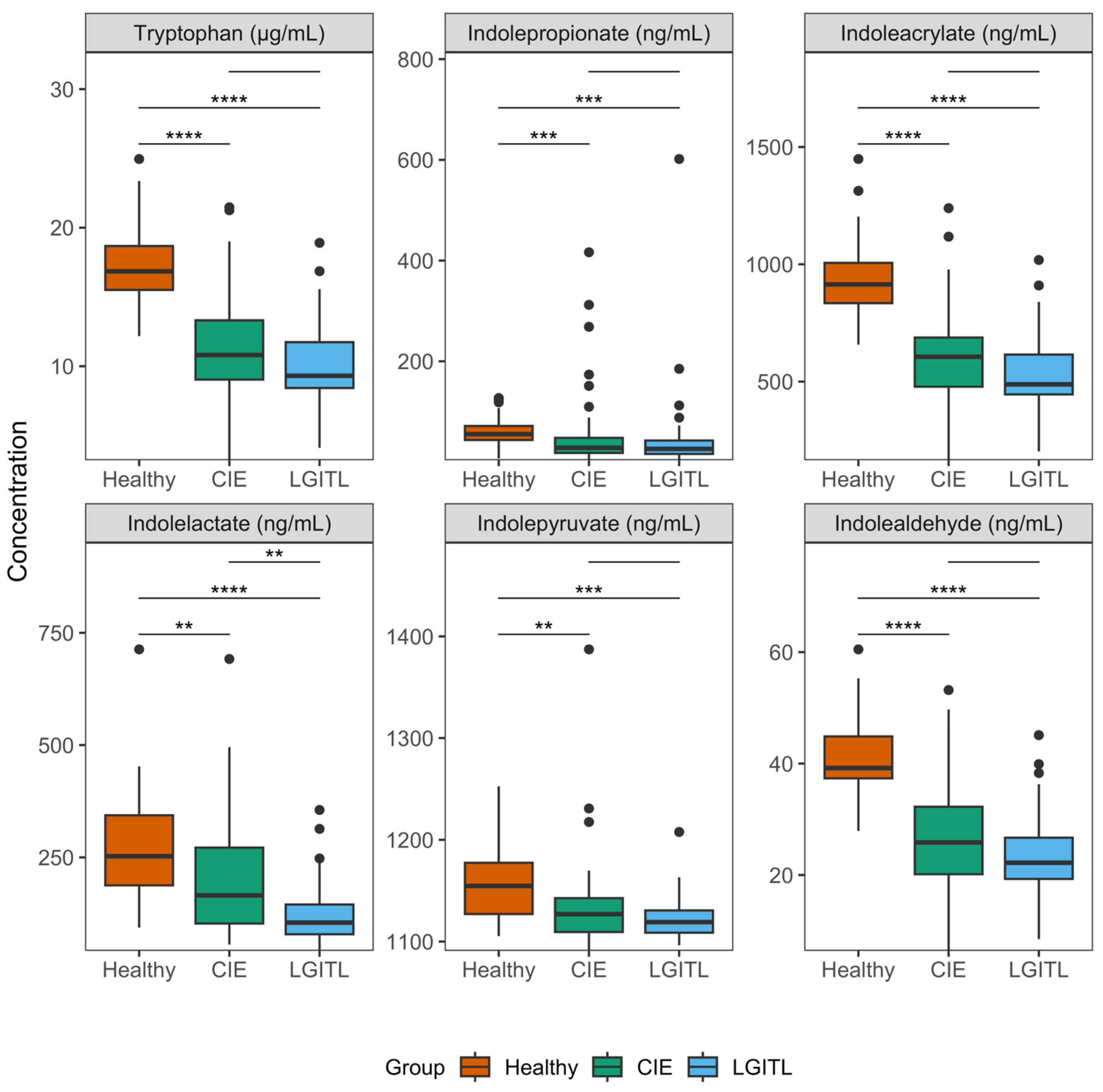
| Tryptophan (µg/mL) | <0.001 | |||
| Median | 16.9 a | 10.8 b | 9.3 b | |
| IQR | 15.5–18.7 | 9.0–13.3 | 8.4–11.7 | |
| Min–Max | 12.2–25.0 | 3.2–21.5 | 4.1–18.9 | |
| Kynurenine (µg/mL) | 0.23 | |||
| Median | 1.04 | 1.14 | 1.26 | |
| IQR | 0.888–1.237 | 0.843–1.57 | 1.01–1.53 | |
| Min–Max | 085–1.94 | 0.513–9.81 | 0.601–2.52 | |
| Kynurenate (ng/mL) | 0.63 | |||
| Median | 8.9 | 10.3 | 9.0 | |
| IQR | 7.9–10.9 | 6.5–14.9 | 6.6–13.8 | |
| Min–Max | 4.6–22.7 | 3.7–136.2 | 4.6–37.3 | |
| Serotonin (ng/mL) | 0.15 | |||
| Median | 888.0 | 760.9 | 954.2 | |
| IQR | 677.8–1160.0 | 516.0–1038.8 | 759.5–1235.9 | |
| Min–Max | 293.9–1439.3 | 12.8–3214.1 | 13.4–2158.5 | |
| Indoleacetate (ng/mL) | 0.21 | |||
| Median | 182.8 | 267.3 | 210.0 | |
| IQR | 139.9–300.4 | 172.9–420.0 | 95.7–418.1 | |
| Min–Max | 62.7–980.1 | 22.1–3219.3 | 51.8–1946.7 | |
| Indolepropionate (ng/mL) | <0.001 | |||
| Median | 55.4 a | 28.2 b | 26.1 b | |
| IQR | 43.7–71.5 | 18.0–48.0 | 16.1–42.7 | |
| Min–Max | 7.2–127.0 | 3.9–416.5 | 3.6–601.7 | |
| Indoleacetamide (ng/mL) | 0.034 | |||
| Median | 1.4 a | 1.4 a | 1.4 a | |
| IQR | 1.4–1.4 | 1.4–1.4 | 1.4–1.4 | |
| Min–Max | 1.3–2.0 | 1.3–2.1 | 1.3–1.5 | |
| Indoleacrylate (ng/mL) | <0.001 | |||
| Median | 914.2 a | 606.0 b | 488.3 b | |
| IQR | 834.5–1006.2 | 478.3–687.8 | 445.5–615.3 | |
| Min–Max | 657.5–1448.8 | 165.0–1239.3 | 202.7–1018.4 | |
| Indolelactate (ng/mL) | <0.001 | |||
| Median | 252.7 a | 165.5 b | 105.0 c | |
| IQR | 187.8–343.8 | 102.9–271.8 | 79.0–145.2 | |
| Min–Max | 93.8–713.1 | 56.0–691.7 | 42.1–355.6 | |
| Indolepyruvate (ng/mL) | 0.001 | |||
| Median | 1154.7 a | 1127.0 b | 1119.1 b | |
| IQR | 1127.1–1177.5 | 1109.4–1142.7 | 1108.7–1130.6 | |
| Min–Max | 1105.4–1252.6 | 1090.9–1387.2 | 1096.3–1207.7 | |
| Indolealdehyde (ng/mL) | <0.001 | |||
| Median | 39.2 a | 25.9 b | 22.2 b | |
| IQR | 37.4–44.9 | 20.2–32.3 | 19.3–26.7 | |
| Min–Max | 27.9–60.5 | 6.4–53.2 | 8.5–45.1 | |
| Tryptamine (ng/mL) | 0.160 | |||
| Median | 3.5 | 3.4 | 3.6 | |
| IQR | 3.2–4.0 | 3.0–3.7 | 3.3–3.9 | |
| Min–Max | 2.6–4.2 | 0.0–6.3 | 2.4–4.8 |
| Variable | Contrast | Dunn’s Z Statistic | padj Value |
|---|---|---|---|
| Tryptophan | Healthy vs. CIE | −4.96 | <0.001 |
| Healthy vs. LGITL | −5.83 | <0.001 | |
| CIE vs. LGITL | −1.37 | 0.17 | |
| Indolepropionate | Healthy vs. CIE | −3.37 | 0.001 |
| Healthy vs. LGITL | −3.60 | <0.001 | |
| CIE vs. LGITL | −0.53 | 0.60 | |
| Indolepyruvate | Healthy vs. CIE | −3.14 | 0.003 |
| Healthy vs. LGITL | −3.43 | 0.002 | |
| CIE vs. LGITL | −0.58 | 0.56 | |
| Indolelactate | Healthy vs. CIE | −2.60 | 0.009 |
| Healthy vs. LGITL | −4.87 | <0.001 | |
| CIE vs. LGITL | −2.77 | 0.008 | |
| Indolealdehyde | Healthy vs. CIE | −4.88 | <0.001 |
| Healthy vs. LGITL | −5.74 | <0.001 | |
| CIE vs. LGITL | −1.36 | 0.17 | |
| Indoleacrylate | Healthy vs. CIE | −5.08 | <0.001 |
| Healthy vs. LGITL | −5.93 | <0.001 | |
| CIE vs. LGITL | −1.36 | 0.17 |
| Clinical Variable | Tryptophan Catabolite | Spearman’s Rho | padj |
|---|---|---|---|
| Cobalamin | Indolealdehyde | 0.39 | <0.001 |
| Tryptophan | 0.39 | 0.001 | |
| Indoleacrylate | 0.37 | 0.001 | |
| Indolepyruvate | 0.31 | 0.007 | |
| Indolelactate | 0.27 | 0.02 | |
| Kynurenine | −0.25 | 0.03 | |
| fPLI | Kynurenine | 0.28 | 0.016 |
| Tryptophan | −0.28 | 0.016 | |
| Indoleacrylate | −0.29 | 0.016 | |
| Indolealdehyde | −0.29 | 0.016 | |
| fTLI | Kynurenine | 0.29 | 0.013 |
| Indolelactate | −0.28 | 0.013 | |
| Indoleacrylate | −0.38 | 0.001 | |
| Indolealdehyde | −0.38 | 0.001 | |
| Tryptophan | −0.38 | 0.001 |
| Spearman’s Rho | padj | ||
|---|---|---|---|
| Tryptamine | Serotonin | 0.98 | <0.001 |
| Tryptophan | Indolealdehyde | 0.99 | <0.001 |
| Indoleacrylate | 0.99 | <0.001 | |
| Indolelactate | 0.64 | <0.001 | |
| Indolepyruvate | 0.42 | <0.001 | |
| Serotonin | Indoleacetate | 0.26 | 0.034 |
| Kynurenate | Kynurenine | 0.72 | <0.001 |
| Indolepropionate | Indoleacetate | 0.27 | 0.026 |
| Indolepyruvate | Indoleacrylate | 0.41 | <0.001 |
| Indolelactate | 0.34 | 0.003 | |
| Indolelactate | Indoleacrylate | 0.62 | <0.001 |
| Indolepropionate | 0.34 | 0.0023 | |
| Indoleacetamide | 0.28 | 0.02 | |
| Indolealdehyde | Indoleacrylate | 0.99 | <0.001 |
| Indolelactate | 0.65 | <0.001 | |
| Indolepyruvate | 0.41 | <0.001 | |
| Indoleacetamide | Indoleacetate | 0.3 | 0.01 |
| Indolepropionate | 0.27 | 0.027 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barko, P.C.; Williams, D.A.; Wu, Y.-A.; Steiner, J.M.; Suchodolski, J.S.; Gal, A.; Marsilio, S. Chronic Inflammatory Enteropathy and Low-Grade Intestinal T-Cell Lymphoma Are Associated with Altered Microbial Tryptophan Catabolism in Cats. Animals 2024, 14, 67. https://doi.org/10.3390/ani14010067
Barko PC, Williams DA, Wu Y-A, Steiner JM, Suchodolski JS, Gal A, Marsilio S. Chronic Inflammatory Enteropathy and Low-Grade Intestinal T-Cell Lymphoma Are Associated with Altered Microbial Tryptophan Catabolism in Cats. Animals. 2024; 14(1):67. https://doi.org/10.3390/ani14010067
Chicago/Turabian StyleBarko, Patrick C., David A. Williams, Yu-An Wu, Joerg M. Steiner, Jan S. Suchodolski, Arnon Gal, and Sina Marsilio. 2024. "Chronic Inflammatory Enteropathy and Low-Grade Intestinal T-Cell Lymphoma Are Associated with Altered Microbial Tryptophan Catabolism in Cats" Animals 14, no. 1: 67. https://doi.org/10.3390/ani14010067
APA StyleBarko, P. C., Williams, D. A., Wu, Y.-A., Steiner, J. M., Suchodolski, J. S., Gal, A., & Marsilio, S. (2024). Chronic Inflammatory Enteropathy and Low-Grade Intestinal T-Cell Lymphoma Are Associated with Altered Microbial Tryptophan Catabolism in Cats. Animals, 14(1), 67. https://doi.org/10.3390/ani14010067