

Age Rather Than Supplementation with Oat β-Glucan Influences Development of the Intestinal Microbiota and SCFA Concentrations in Suckling Piglets

 , , ,
, , ,  ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals
2.3. Housing and Management
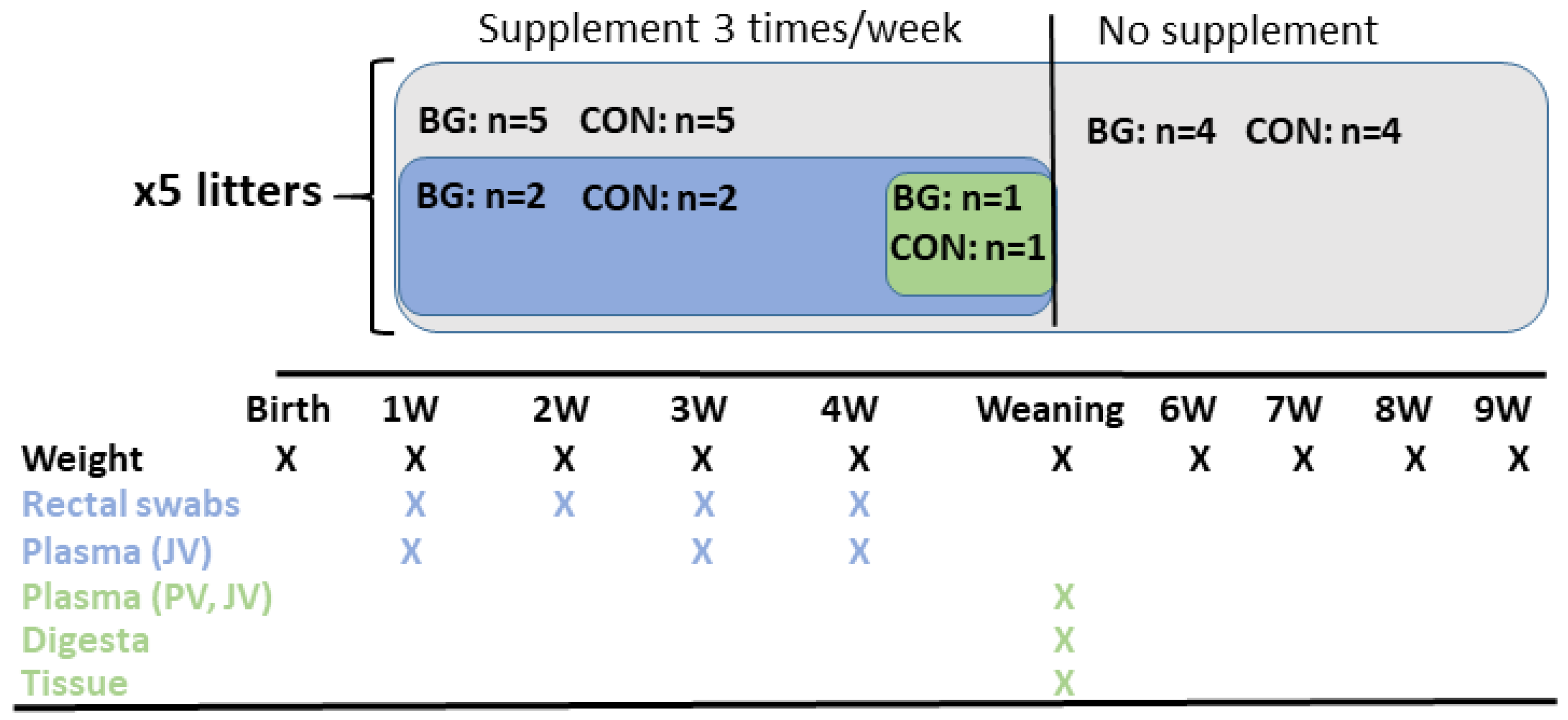
2.4. Treatments
2.5. Experimental Procedures
2.6. Post-Mortem Sampling Procedures
2.7. Histology of Intestinal Tissue
2.8. DNA Extraction, PCR Conditions and Sequence Analysis
2.9. Determination of SCFAs in Plasma and Digesta
2.10. Statistical Analysis
3. Results
3.1. Animal Growth and Performance
3.2. Development of the Microbiota
3.3. SCFA and Caproic Acid Concentrations in Plasma and Colon Digesta
3.4. Gut Histological Measurements
3.5. Correlations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aluthge, N.D.; Van Sambeek, D.M.; Carney-Hinkle, E.E.; Li, Y.S.; Fernando, S.C.; Burkey, T.E. The pig microbiota and the potential for harnessing the power of the microbiome to improve growth and health1. J. Anim. Sci. 2019, 97, 3741–3757. [Google Scholar] [CrossRef] [PubMed]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut Microbiota Dysbiosis in Postweaning Piglets: Understanding the Keys to Health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.K.; Kye, Y.C.; Kim, G.; Kim, H.W.; Gu, M.J.; Umboh, J.; Maaruf, K.; Kim, S.W.; Yun, C.H. Stress, Nutrition, and Intestinal Immune Responses in Pigs—A Review. Asian-Australas J. Anim. Sci. 2016, 29, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Xiong, K.; Fang, R.; Li, M. Weaning stress and intestinal health of piglets: A review. Front. Immunol. 2022, 13, 1042778. [Google Scholar] [CrossRef]
- Rhouma, M.; Fairbrother, J.M.; Beaudry, F.; Letellier, A. Post weaning diarrhea in pigs: Risk factors and non-colistin-based control strategies. Acta Vet. Scand. 2017, 59, 31. [Google Scholar] [CrossRef]
- Van Hees, H.M.J.; Davids, M.; Maes, D.; Millet, S.; Possemiers, S.; den Hartog, L.A.; van Kempen, T.; Janssens, G.P.J. Dietary fibre enrichment of supplemental feed modulates the development of the intestinal tract in suckling piglets. J. Anim. Sci. Biotechnol. 2019, 10, 83. [Google Scholar] [CrossRef]
- Vetvicka, V.; Vannucci, L.; Sima, P.; Richter, J. Beta Glucan: Supplement or Drug? From Laboratory to Clinical Trials. Molecules 2019, 24, 1251. [Google Scholar] [CrossRef]
- Jayachandran, M.; Chen, J.; Chung, S.S.M.; Xu, B. A critical review on the impacts of β-glucans on gut microbiota and human health. J. Nutr. Biochem. 2018, 61, 101–110. [Google Scholar] [CrossRef]
- Jha, R.; Fouhse, J.M.; Tiwari, U.P.; Li, L.; Willing, B.P. Dietary Fiber and Intestinal Health of Monogastric Animals. Front. Vet. Sci. 2019, 6, 48. [Google Scholar] [CrossRef]
- Jha, R.; Berrocoso, J.D. Review: Dietary fiber utilization and its effects on physiological functions and gut health of swine. Animal 2015, 9, 1441–1452. [Google Scholar] [CrossRef]
- Tian, G.; Wu, X.; Chen, D.; Yu, B.; He, J. Adaptation of gut microbiome to different dietary nonstarch polysaccharide fractions in a porcine model. Mol. Nutr. Food Res. 2017, 61, 10. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Li, P.; Wang, F. β-glucans as potential immunoadjuvants: A review on the adjuvanticity, structure-activity relationship and receptor recognition properties. Vaccine 2018, 36, 5235–5244. [Google Scholar] [CrossRef] [PubMed]
- Bedford, A.; Gong, J. Implications of butyrate and its derivatives for gut health and animal production. Anim. Nutr. 2018, 4, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, P.; Araújo, J.R.; Di Santo, J.P. A Cross-Talk Between Microbiota-Derived Short-Chain Fatty Acids and the Host Mucosal Immune System Regulates Intestinal Homeostasis and Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2018, 24, 558–572. [Google Scholar] [CrossRef]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; de Los Reyes-Gavilán, C.G.; Salazar, N. Intestinal Short Chain Fatty Acids and their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef]
- Calderón-Pérez, L.; Gosalbes, M.J.; Yuste, S.; Valls, R.M.; Pedret, A.; Llauradó, E.; Jimenez-Hernandez, N.; Artacho, A.; Pla-Pagà, L.; Companys, J.; et al. Gut metagenomic and short chain fatty acids signature in hypertension: A cross-sectional study. Sci. Rep. 2020, 10, 6436. [Google Scholar] [CrossRef]
- Knudsen, K.E.B.; Jørgensen, H.; Theil, P.K. Changes in short-chain fatty acid plasma profile incurred by dietary fiber composition. J. Anim. Sci. 2016, 94, 476–479. [Google Scholar] [CrossRef]
- Verbeek, E.; Keeling, L.; Landberg, R.; Lindberg, J.E.; Dicksved, J. The gut microbiota and microbial metabolites are associated with tail biting in pigs. Sci. Rep. 2021, 11, 20547. [Google Scholar] [CrossRef]
- Muller, M.; Hernandez, M.A.G.; Goossens, G.H.; Reijnders, D.; Holst, J.J.; Jocken, J.W.E.; van Eijk, H.; Canfora, E.E.; Blaak, E.E. Circulating but not faecal short-chain fatty acids are related to insulin sensitivity, lipolysis and GLP-1 concentrations in humans. Sci. Rep. 2019, 9, 12515. [Google Scholar] [CrossRef]
- Choudhury, R.; Middelkoop, A.; Boekhorst, J.; Gerrits, W.J.J.; Kemp, B.; Bolhuis, J.E.; Kleerebezem, M. Early life feeding accelerates gut microbiome maturation and suppresses acute post-weaning stress in piglets. Environ. Microbiol. 2021, 23, 7201–7213. [Google Scholar] [CrossRef]
- Choudhury, R.; Middelkoop, A.; De Souza, J.G.; Van Veen, L.A.; Gerrits, W.J.; Kemp, B.; Bolhuis, J.E.; Kleerebezem, M. Impact of early-life feeding on local intestinal microbiota and digestive system development in piglets. Sci. Rep. 2021, 11, 1–16. [Google Scholar]
- du Sert, N.P.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar]
- Wallgren, P.; Vallgårda, J. Seroproduktion—Presentation, definition och kravlista. Sven. Vet. Tidn. 1993, 45, 733–735. [Google Scholar]
- Müller, B.; Sun, L.; Westerholm, M.; Schnürer, A. Bacterial community composition and fhs profiles of low- and high-ammonia biogas digesters reveal novel syntrophic acetate-oxidising bacteria. Biotechnol. Biofuels 2016, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. PyNAST: A flexible tool for aligning sequences to a template alignment. Bioinformatics 2010, 26, 266–267. [Google Scholar] [CrossRef]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classsifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Han, J.; Lin, K.; Sequeira, C.; Borchers, C.H. An isotope-labeled chemical derivatization method for the quantitation of short-chain fatty acids in human feces by liquid chromatography-tandem mass spectrometry. Anal. Chim. Acta 2015, 854, 86–94. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological Software Package for Education and Data Analysis. Paleontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Tremblay Antoine, R.J. LMER Convenience Functions: Model Selection and Post-Hoc Analysis for (G)LMER Models, R Package Version 3.0; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Lenth, R.V. Estimated Marginal Means, aka Least-Squares Means, R Package Version 1.5.1; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple hypothesis testing. J. Roy. Statist. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Qi, R.; Qiu, X.; Du, L.; Wang, J.; Wang, Q.; Huang, J.; Liu, Z. Changes of Gut Microbiota and Its Correlation With Short Chain Fatty Acids and Bioamine in Piglets at the Early Growth Stage. Front. Vet. Sci. 2020, 7, 617259. [Google Scholar] [CrossRef] [PubMed]
- Bian, G.; Ma, S.; Zhu, Z.; Su, Y.; Zoetendal, E.G.; Mackie, R.; Liu, J.; Mu, C.; Huang, R.; Smidt, H.; et al. Age, Introduction of Solid Feed and Weaning Are More Important Determinants of Gut Bacterial Succession in Piglets than Breed and Nursing Mother as Revealed by a Reciprocal Cross-Fostering Model: Gut Bacterial Succession in Piglets. Environ. Microbiol. 2016, 18, 1566. [Google Scholar] [CrossRef] [PubMed]
- Frese, S.A.; Parker, K.; Calvert, C.C.; Mills, D.A. Diet shapes the gut microbiome of pigs during nursing and weaning. Microbiome 2015, 3, 28. [Google Scholar] [CrossRef]
- Mach, N.; Berri, M.; Estellé, J.; Levenez, F.; Lemonnier, G.; Denis, C.; Leplat, J.-J.; Chevaleyre, C.; Billon, Y.; Doré, J.; et al. Early-life establishment of the swine gut microbiome and impact on host phenotypes. Environ. Microbiol. Rep. 2015, 7, 554–569. [Google Scholar] [CrossRef]
- Wang, X.; Tsai, T.; Deng, F.; Wei, X.; Chai, J.; Knapp, J.; Apple, J.; Maxwell, C.V.; Lee, J.A.; Li, Y.; et al. Longitudinal investigation of the swine gut microbiome from birth to market reveals stage and growth performance associated bacteria. Microbiome 2019, 7, 109. [Google Scholar] [CrossRef]
- Ke, S.; Fang, S.; He, M.; Huang, X.; Yang, H.; Yang, B.; Chen, C.; Huang, L. Age-based dynamic changes of phylogenetic composition and interaction networks of health pig gut microbiome feeding in a uniformed condition. BMC Vet. Res. 2019, 15, 172. [Google Scholar] [CrossRef]
- Guevarra, R.B.; Hong, S.H.; Cho, J.H.; Kim, B.-R.; Shin, J.; Lee, J.H.; Kang, B.N.; Kim, Y.H.; Wattanaphansak, S.; Isaacson, R.E.; et al. The dynamics of the piglet gut microbiome during the weaning transition in association with health and nutrition. J. Anim. Sci. Biotechnol. 2018, 9, 54. [Google Scholar] [CrossRef]
- Guevarra, R.B.; Lee, J.H.; Lee, S.H.; Seok, M.-J.; Kim, D.W.; Kang, B.N.; Johnson, T.J.; Isaacson, R.E.; Kim, H.B. Piglet gut microbial shifts early in life: Causes and effects. J. Anim. Sci. Biotechnol. 2019, 10, 1. [Google Scholar] [CrossRef]
- Ivarsson, E.; Roos, S.; Liu, H.Y.; Lindberg, J.E. Fermentable non-starch polysaccharides increases the abundance of Bacteroides–Prevotella–Porphyromonas in ileal microbial community of growing pigs. Animal 2014, 8, 1777–1787. [Google Scholar] [CrossRef]
- Fraser, D.; Pajor, E.A.; Feddes, J.J.R. The relationship between creep feeding behavior of piglets and adaptation to weaning: Effect of diet quality. Can. J. Anim. Sci. 1994, 74, 1–6. [Google Scholar] [CrossRef]
- Zhou, Z.; Zheng, W.; Shang, W.; Du, H.; Li, G.; Yao, W. How host gender affects the bacterial community in pig feces and its correlation to skatole production. Ann. Microbiol. 2015, 65, 2379–2386. [Google Scholar] [CrossRef]
- He, M.; Gao, J.; Wu, J.; Zhou, Y.; Fu, H.; Ke, S.; Yang, H.; Chen, C.; Huang, L. Host Gender and Androgen Levels Regulate Gut Bacterial Taxa in Pigs Leading to Sex-Biased Serum Metabolite Profiles. Front. Microbiol. 2019, 10, 1359. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yin, J.; Tan, B.; Chen, J.; Zhang, H.; Li, Z.; Ma, X. Physiological function and application of dietary fiber in pig nutrition: A review. Anim. Nutr. 2021, 7, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Wegh, C.A.M.; Geerlings, S.Y.; Knol, J.; Roeselers, G.; Belzer, C. Postbiotics and Their Potential Applications in Early Life Nutrition and Beyond. Int. J. Mol. Sci. 2019, 20, 4673. [Google Scholar] [CrossRef] [PubMed]
- Hume, I.D. Flow dynamics of digesta and colonic fermentation. In Physiological and Clinical Aspects of Short-Chain Fatty Acids; Cummings, J.H., Rombeau, J.L., Sakata, T., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 119–132. [Google Scholar]
- Fernández, J.; Redondo-Blanco, S.; Gutiérrez-del-Río, I.; Miguélez, E.M.; Villar, C.J.; Lombo, F. Colon microbiota fermentation of dietary prebiotics towards short-chain fatty acids and their roles as anti-inflammatory and antitumour agents: A review. J. Func. Foods 2016, 25, 511–522. [Google Scholar] [CrossRef]
- Feng, W.; Ao, H.; Peng, C. Gut Microbiota, Short-Chain Fatty Acids, and Herbal Medicines. Front. Pharmacol. 2018, 9, 1354. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- van Beers-Schreurs, H.M.; Nabuurs, M.J.; Vellenga, L.; Kalsbeek-van der Valk, H.J.; Wensing, T.; Breukink, H.J. Weaning and the weanling diet influence the villous height and crypt depth in the small intestine of pigs and alter the concentrations of short-chain fatty acids in the large intestine and blood. J. Nutr. 1998, 128, 947–953. [Google Scholar] [CrossRef]
- Müller, M.; Hermes, G.D.A.; Canfora, E.E.; Smidt, H.; Masclee, A.A.M.; Zoetendal, E.G.; Blaak, E.E. Distal colonic transit is linked to gut microbiota diversity and microbial fermentation in humans with slow colonic transit. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, 361–369. [Google Scholar] [CrossRef]
- Nakatani, M.; Inoue, R.; Tomonaga, S.; Fukuta, K.; Tsukahara, T. Production, Absorption, and Blood Flow Dynamics of Short-Chain Fatty Acids Produced by Fermentation in Piglet Hindgut during the Suckling-Weaning Period. Nutrients 2018, 10, 1220. [Google Scholar] [CrossRef]
- Metzler-Zebeli, B.U.; Zijlstra, R.T.; Mosenthin, R.; Gänzle, M.G. Dietary calcium phosphate content and oat β-glucan influence gastrointestinal microbiota, butyrate-producing bacteria and butyrate fermentation in weaned pigs. FEMS Microbiol. Ecol. 2011, 75, 402–413. [Google Scholar] [CrossRef]
- He, B.; Bai, Y.; Jiang, L.; Wang, W.; Li, T.; Liu, P.; Tao, S.; Zhao, J.; Han, D.; Wang, J. Effects of Oat Bran on Nutrient Digestibility, Intestinal Microbiota, and Inflammatory Responses in the Hindgut of Growing Pigs. Int. J. Mol. Sci. 2018, 19, 2407. [Google Scholar] [CrossRef]
- Pieper, R.; Jha, R.; Rossnagel, B.; Van Kessel, A.G.; Souffrant, W.B.; Leterme, P. Effect of barley and oat cultivars with different carbohydrate compositions on the intestinal bacterial communities in weaned piglets. FEMS Microbiol. Ecol. 2008, 66, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Hiss, S.; Sauerwein, H. Influence of dietary ß-glucan on growth performance, lymphocyte proliferation, specific immune response and haptoglobin plasma concentrations in pigs. J. Anim. Physiol. Anim. Nutr. 2003, 87, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Feng, M.; Chu, Y.; Wang, S.; Shete, V.; Tuohy, K.M.; Liu, F.; Zhou, X.; Kamil, A.; Pan, D.; et al. The Prebiotic Effects of Oats on Blood Lipids, Gut Microbiota, and Short-Chain Fatty Acids in Mildly Hypercholesterolemic Subjects Compared With Rice: A Randomized, Controlled Trial. Front. Immunol. 2021, 12, 787797. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, U.P.; Singh, A.K.; Jha, R. Fermentation characteristics of resistant starch, arabinoxylan, and β-glucan and their effects on the gut microbial ecology of pigs: A review. Anim. Nutr. 2019, 5, 217–226. [Google Scholar] [CrossRef]
- Zhou, Y.; Luo, Y.; Yu, B.; Zheng, P.; Yu, J.; Huang, Z.; Mao, X.; Luo, J.; Yan, H.; He, J. Effect of β-Glucan Supplementation on Growth Performance and Intestinal Epithelium Functions in Weaned Pigs Challenged by Enterotoxigenic Escherichia coli. Antibiotics 2022, 11, 519. [Google Scholar] [CrossRef]
- Vetvicka, V.; Vetvickova, J. Physiological Effects of Different Types of Beta-Glucan. Biomed. Pap. 2007, 151, 225–231. [Google Scholar]
- Dong, J.L.; Yu, X.; Dong, L.E.; Shen, R.L. In vitro fermentation of oat [beta]-glucan and hydrolysates by fecal microbiota and selected probiotic strains. J. Sci. Food Agric. 2017, 97, 4198. [Google Scholar] [CrossRef]
- Gong, L.; Cao, W.; Chi, H.; Wang, J.; Zhang, H.; Liu, J.; Sun, B. Whole cereal grains and potential health effects: Involvement of the gut microbiota. Food Res. Int. 2018, 103, 84–102. [Google Scholar] [CrossRef] [PubMed]
- Volman, J.J.; Ramakers, J.D.; Plat, J. Dietary modulation of immune function by beta-glucans. Physiol. Behav. 2008, 94, 276–284. [Google Scholar] [CrossRef]
- Stier, H.; Ebbeskotte, V.; Gruenwald, J. Immune-modulatory effects of dietary Yeast Beta-1,3/1,6-D-glucan. Nutr. J. 2014, 13, 38. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, J.; Schloermann, W.; Trautvetter, U.; Glei, M. The effects of β-glucans on intestinal health. Ernahr. Umsch. 2020, 67, 52–59. [Google Scholar]
- Geervliet, M.; Lute, L.C.; Jansen, C.A.; Rutten, V.P.; Savelkoul, H.F.; Tijhaar, E. Differential immunomodulation of porcine bone marrow derived dendritic cells by E. coli Nissle 1917 and β-glucans. PLoS ONE 2020, 15, e0233773. [Google Scholar] [CrossRef] [PubMed]
- Sonck, E.; Devriendt, B.; Goddeeris, B.; Cox, E. Varying effects of different β-glucans on the maturation of porcine monocyte-derived dendritic cells. Clin. Vacc. Immunol. 2011, 18, 1441–1446. [Google Scholar] [CrossRef]
- de Vries, H.; Geervliet, M.; Jansen, C.A.; Rutten, V.; van Hees, H.; Groothuis, N.; Wells, J.M.; Savelkoul, H.F.J.; Tijhaar, E.; Smidt, H. Impact of Yeast-Derived β-Glucans on the Porcine Gut Microbiota and Immune System in Early Life. Microorganisms 2020, 8, 157. [Google Scholar] [CrossRef] [PubMed]
- Mejía, S.M.V.; de Francisco, A.; Bohrer, B. A comprehensive review on cereal β-glucan: Extraction, characterization, causes of degradation, and food application. Crit. Rev. Food Sci. Nutr. 2020, 60, 3693–3704. [Google Scholar] [CrossRef]
- Shen, R.L.; Dang, X.Y.; Dong, J.L.; Hu, X.Z. Effects of oat β-glucan and barley β-glucan on fecal characteristics, intestinal microflora, and intestinal bacterial metabolites in rats. J. Agric. Food Chem. 2012, 60, 11301–11308. [Google Scholar] [CrossRef]
- Boes, J.; Johansen, M.V.; Eriksen, L.; Bogh, H.O.; Nansen, P.; Stephenson, L.S. False-positive Trichuris suis egg counts in pigs in relation to coprophagia. Parasite 1998, 5, 91–93. [Google Scholar] [CrossRef]
- Luo, J.; Chen, D.; Mao, X.; He, J.; Yu, B.; Cheng, L.; Zeng, D. Purified β-glucans of Different Molecular Weights Enhance Growth Performance of LPS-challenged Piglets via Improved Gut Barrier Function and Microbiota. Animals 2019, 9, 602. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Song, P.; Fan, P.; Hou, C.; Thacker, P.; Ma, X. Dietary Sodium Butyrate Decreases Postweaning Diarrhea by Modulating Intestinal Permeability and Changing the Bacterial Communities in Weaned Piglets. J. Nutr. 2015, 145, 2774–2780. [Google Scholar] [CrossRef]
- Guilloteau, P.; Martin, L.; Eeckhaut, V.; Ducatelle, R.; Zabielski, R.; Van Immerseel, F. From the gut to the peripheral tissues: The multiple effects of butyrate. Nutr. Res. Rev. 2010, 23, 366–384. [Google Scholar] [CrossRef] [PubMed]
- Lalles, J.P.; Boudry, G.; Favier, C.; Le Floc’H, N.; Luron, I.; Montagne, L.; Oswald, I.P.; Pié, S.; Piel, C.; Sève, B. Gut function and dysfunction in young pigs: Physiology. Anim. Res. 2004, 53, 301–316. [Google Scholar] [CrossRef]
- Inoue, R.; Tsukahara, T.; Nakatani, M.; Okutani, M.; Nishibayashi, R.; Ogawa, S.; Harayama, T.; Nagino, T.; Hatanaka, H.; Fukuta, K.; et al. Weaning Markedly Affects Transcriptome Profiles and Peyer’s Patch Development in Piglet Ileum. Front. Immunol. 2015, 6, 630. [Google Scholar] [CrossRef]
- Alizadeh, A.; Akbari, P.; Difilippo, E.; Schols, H.A.; Ulfman, L.H.; Schoterman, M.H.C.; Garssen, J.; Fink-Gremmels, J.; Braber, S. The piglet as a model for studying dietary components in infant diets: Effects of galacto-oligosaccharides on intestinal functions. Br. J. Nutr. 2016, 115, 605–618. [Google Scholar] [CrossRef]
- Bruininx, E.M.; Binnendijk, G.P.; van der Peet-Schwering, C.M.; Schrama, J.W.; den Hartog, L.A.; Everts, H.; Beynen, A.C. Effect of creep feed consumption on individual feed intake characteristics and performance of group-housed weanling pigs. J. Anim. Sci. 2002, 80, 1413–1418. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BG | CON | T | A | A:S | T:A | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1w | 2w | 3w | 4w | 1w | 2w | 3w | 4w | |||||
| Shannon index | 3.26 ± 0.13 | 3.59 ± 0.19 | 3.63 ± 0.14 | 4.19 ± 0.21 | 3.13 ± 0.17 | 3.68 ± 0.14 | 3.73 ± 0.19 | 4.21 ± 0.26 a | ns | * | * | ns |
| Chao-1 | 538 ± 38 | 803 ± 35 | 910 ± 40 | 980 ± 50 | 535 ± 30 | 798 ± 39 | 939 ± 38 | 977 ± 62 a | ns | * | * | ns |
| Bacteria | BG | CON | T | A | T:A | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1w | 2w | 3w | 4w | 1w | 2w | 3w | 4w | ||||
| Phylum | |||||||||||
| Actinobacteria | 0.88 ± 0.40 a | 0.35 ± 0.08 b | 1.01 ± 0.35 b | 0.35 ± 0.13 b | 2.01 ± 0.66 a | 0.41 ± 0.19 b | 0.43 ± 0.15 b | 0.30 ± 0.19 b | ns | * | ns |
| Bacteroidetes | 40.1 ± 4.67 a | 26.3 ± 3.07 | 30.8 ± 4.47 | 33.7 ± 4.16 | 27.6 ± 6.38 a | 31.6 ± 3.59 | 33.2 ± 4.58 | 42.6 ± 3.79 | ns | ns | ns |
| Firmicutes | 34.6 ± 3.32 | 41.7 ± 3.71 a | 33.3 ± 2.10 b | 32.3 ± 3.62 b | 34.8 ± 3.20 | 39.1 ± 2.84 a | 32.0 ± 2.83 b | 28.5 ± 2.77 b | ns | * | ns |
| Fusobacteria | 1.55 ± 0.34 a | 0.11 ± 0.05 b | 0.01 ± 0.01 b | 0.08 ± 0.03 b | 1.29 ± 0.33 a | 0.21 ± 0.11 b | 0.03 ± 0.01 b | 0.42 ± 0.30 b | ns | * | ns |
| Proteobacteria | 22.9 ± 5.08 | 27.7 ± 5.23 | 30.1 ± 4.45 | 27.6 ± 5.59 | 34.3 ± 6.51 | 23.2 ± 3.26 | 29.4 ± 6.09 | 18.5 ± 4.93 | ns | ns | ns |
| Genera | |||||||||||
| Escherichia | 19.9 ± 5.15 | 25.6 ± 5.27 | 30.2 ± 3.75 | 23.7 ± 5.79 A | 32.0 ± 6.46 | 21.6 ± 3.22 | 24.5 ± 5.35 | 13.5 ± 4.86 A | ns | ns | * |
| Bacteroides | 28.4 ± 4.75 A | 13.9 ± 3.24 B | 14.4 ± 3.03 | 10.6 ± 2.06 | 17.4 ± 4.27 A | 23.9 ± 3.57 B | 14.1 ± 1.97 | 11.4 ± 3.24 | ns | ns | * |
| Prevotella | 3.64 ± 1.60 a | 7.31 ± 2.28 b,c,A | 9.46 ± 3.46 b,d | 12.1 ± 3.70 b,d | 2.0 ± 0.61 a | 1.99 ± 0.43 b,c,A | 10.6 ± 3.10 b,d | 18.3 ± 5.65 b,d | ns | * | * |
| Lactobacillus | 9.44 ± 1.95 a | 8.45 ± 1.82 c | 4.65 ± 1.53 b,d,e | 2.63 ± 0.79 b,d,f | 12.5 ± 3.12 a | 12.1 ± 2.60 c | 1.67 ± 0.37 b,d,e | 2.95 ± 0.60 b,d,f | ns | * | ns |
| Lachnospiraceae | 7.73 ± 2.64 | 7.44 ± 2.96 | 7.33 ± 2.60 | 4.45 ± 1.67 | 3.10 ± 1.56 | 4.33 ± 1.36 | 7.05 ± 3.50 | 3.49 ± 0.99 | ns | ns | ns |
| Ruminococcaceae; other | 1.93 ± 0.37 a | 1.53 ± 0.70 b,c | 1.00 ± 0.15 | 0.97 ± 0.21 d | 2.39 ± 0.51 a | 0.63 ± 0.09 b,c | 2.03 ± 0.82 | 0.99 ± 0.36 d | ns | * | ns |
| Oscillospira | 1.78 ± 0.37 a | 3.82 ± 0.74 b | 2.74 ± 0.47 b | 3.43 ± 0.84 | 1.46 ± 0.45 a | 4.03 ± 0.64 b | 3.21 ± 0.79 b | 1.89 ± 0.28 | ns | * | ns |
| S24_7:g | 0.27 ± 0.12 a | 2.11 ± 0.68 b,c | 1.94 ± 0.41 b | 3.11 ± 0.91 b,d | 0.29 ± 0.13 a | 1.31 ± 0.36 b,c | 2.00 ± 0.48 b | 2.72 ± 0.55 b,d | ns | * | ns |
| [Prevotella] | 0.53 ± 0.44 a | 1.21 ± 0.48 | 0.88 ± 0.40 b,c | 1.93 ± 0.44 b,d | 0.29 ± 0.16 a | 0.94 ± 0.34 | 2.56 ± 1.06 b,c | 3.66 ± 1.14 b,d | ns | * | ns |
| Ruminococcus | 1.93 ± 0.37 a | 1.53 ± 0.70 b | 1.00 ± 0.15 c | 0.97 ± 0.21 d | 2.39 ± 0.51 a | 0.63 ± 0.09 b | 2.03 ± 0.82 c | 0.99 ± 0.36 d | ns | * | ns |
| SCFA | BG | CON | T | A | T:A | ||||
|---|---|---|---|---|---|---|---|---|---|
| 1w | 3w | 4w | 1w | 3w | 4w | ||||
| Acetic acid | 10.9 ± 0.69 a | 17.1 ± 0.85 b,c | 18.9 ± 1.10 b,d | 13.0 ± 0.76 a | 18.4 ± 1.44 b,c | 21.7 ± 1.27 b,d | * | * | ns |
| Propionic acid | 0.88 ± 0.12 | 1.19 ± 0.20 a | 1.27 ± 0.23 | 1.03 ± 0.09 | 1.82 ± 0.34 a | 1.21 ± 0.21 | ns | * | ns |
| Butyric acid | 0.15 ± 0.30 | 0.38 ± 0.71 a | 0.68 ± 0.10 a | 0.21 ± 0.05 | 0.34 ± 0.11 a | .045 ± 0.08 a | ns | * | ns |
| Formic acid | 576 ± 20.3 a | 447 ± 34.7 b,c | 346 ± 30.1 b,d | 546 ± 17.6 a | 494 ± 53.4 b,c | 419 ± 50.5 b,d | ns | * | ns |
| Iso-butyric acid | 0.42 ± 0.04 a | 0.69 ± 0.16 b | 0.81 ± 0.12 b | 0.43 ± 0.06 a | 0.83 ± 0.13 b | 0.57 ± 0.08 b | ns | * | ns |
| Valeric acid | 0.08 ± 0.00 a | 0.14 ± 0.03 b | 0.16 ± 0.03 b | 0.06 ± 0.02 a | 0.08 ± 0.02 b | 0.11 ± 0.01 b | ns | * | ns |
| Succinic acid | 9.07 ± 1.66 a | 7.62 ± 0.94 | 7.28 ± 1.36 b | 11.89 ± 1.80 a | 9.34 ± 1.38 | 7.52 ± 0.96 b | ns | * | ns |
| Caproic acid | 0.27 ± 0.02 | 0.29 ± 0.02 | 0.24 ± 0.03 | 0.23 ± 0.03 | 0.23 ± 0.04 | 0.20 ± 0.03 | ns | ns | ns |
| N (1) | CON | BG | p-Value | |
|---|---|---|---|---|
| Ileum (µm) | ||||
| Villus height | 10 | 346 ± 19.6 | 360 ± 25.1 | 0.58 |
| Crypt depth | 5–9 | 234 ± 15.3 | 238 ± 21.9 | 0.82 |
| Thickness mucosa | 10 | 598 ± 30.7 | 619 ± 36.0 | 0.62 |
| Thickness muscularis externa | 10 | 543 ± 33.3 | 480± 24.5 | 0.34 |
| Thickness total gut | 5 | 1925 ± 70.1 | 2199 ± 274 | 0.65 |
| Colon (µm) | ||||
| Crypt depth | 10 | 410 ± 13.0 | 376 ± 17.2 | 0.42 |
| Thickness mucosa | 10 | 483 ± 14.0 | 445 ± 9.4 | 0.61 |
| Thickness muscularis externa | 10 | 400 ± 20.5 | 366 ± 20.4 | 0.57 |
| Thickness total gut | 5–6 | 1152 ± 80.0 | 1042 ± 76.5 | 0.57 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arapovic, L.; Huang, Y.; Manell, E.; Verbeek, E.; Keeling, L.; Sun, L.; Landberg, R.; Lundh, T.; Lindberg, J.E.; Dicksved, J. Age Rather Than Supplementation with Oat β-Glucan Influences Development of the Intestinal Microbiota and SCFA Concentrations in Suckling Piglets. Animals 2023, 13, 1349. https://doi.org/10.3390/ani13081349
Arapovic L, Huang Y, Manell E, Verbeek E, Keeling L, Sun L, Landberg R, Lundh T, Lindberg JE, Dicksved J. Age Rather Than Supplementation with Oat β-Glucan Influences Development of the Intestinal Microbiota and SCFA Concentrations in Suckling Piglets. Animals. 2023; 13(8):1349. https://doi.org/10.3390/ani13081349
Chicago/Turabian StyleArapovic, Lidija, Yi Huang, Elin Manell, Else Verbeek, Linda Keeling, Li Sun, Rikard Landberg, Torbjörn Lundh, Jan Erik Lindberg, and Johan Dicksved. 2023. "Age Rather Than Supplementation with Oat β-Glucan Influences Development of the Intestinal Microbiota and SCFA Concentrations in Suckling Piglets" Animals 13, no. 8: 1349. https://doi.org/10.3390/ani13081349
APA StyleArapovic, L., Huang, Y., Manell, E., Verbeek, E., Keeling, L., Sun, L., Landberg, R., Lundh, T., Lindberg, J. E., & Dicksved, J. (2023). Age Rather Than Supplementation with Oat β-Glucan Influences Development of the Intestinal Microbiota and SCFA Concentrations in Suckling Piglets. Animals, 13(8), 1349. https://doi.org/10.3390/ani13081349