
How Sanctuary Chimpanzees (Pan troglodytes) Use Space after Being Introduced to a Large Outdoor Habitat


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
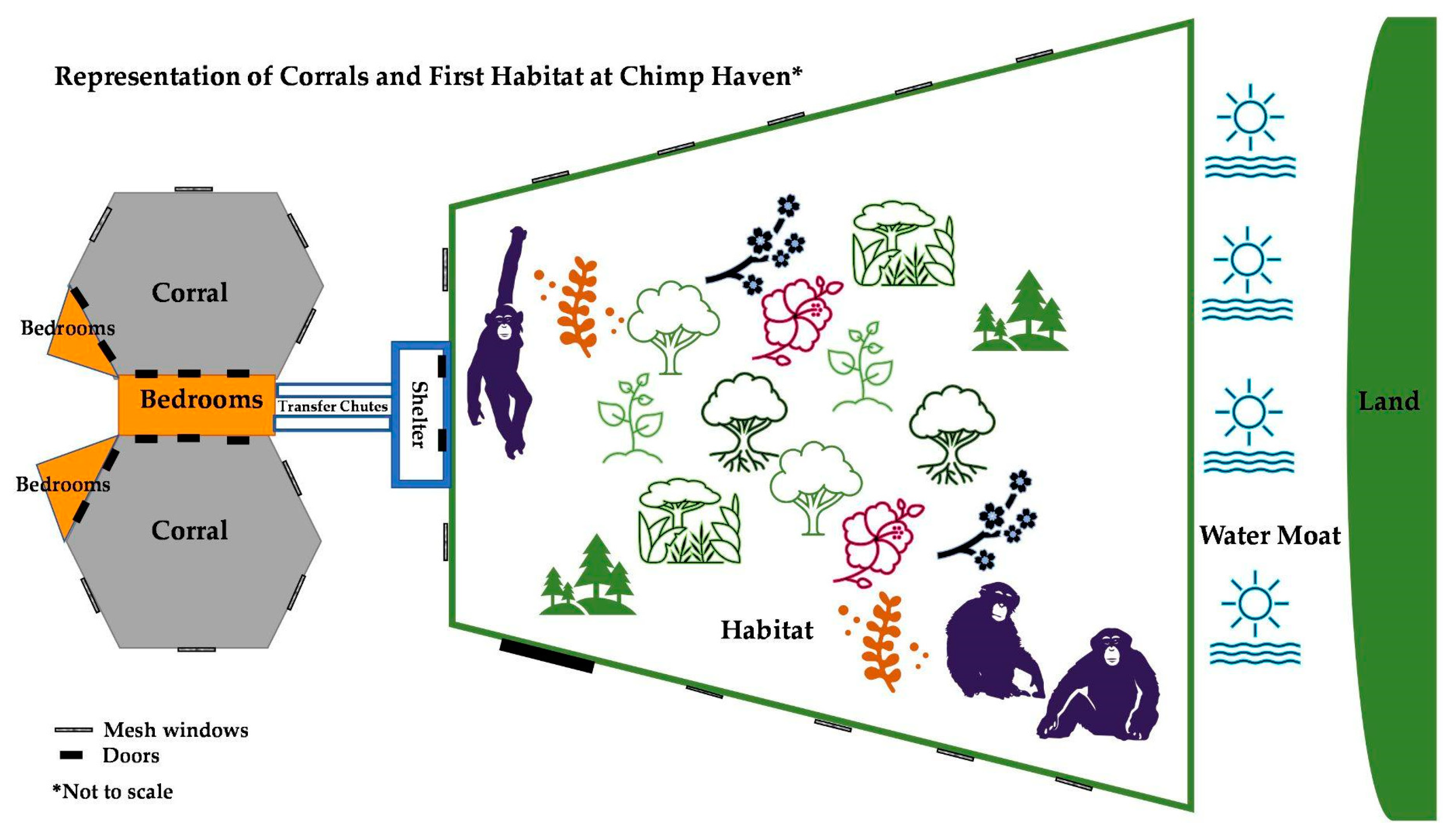
2.1. Facilities
2.2. Subjects
2.3. Data Collection
2.4. Data Analysis
Statistical Analysis
3. Results
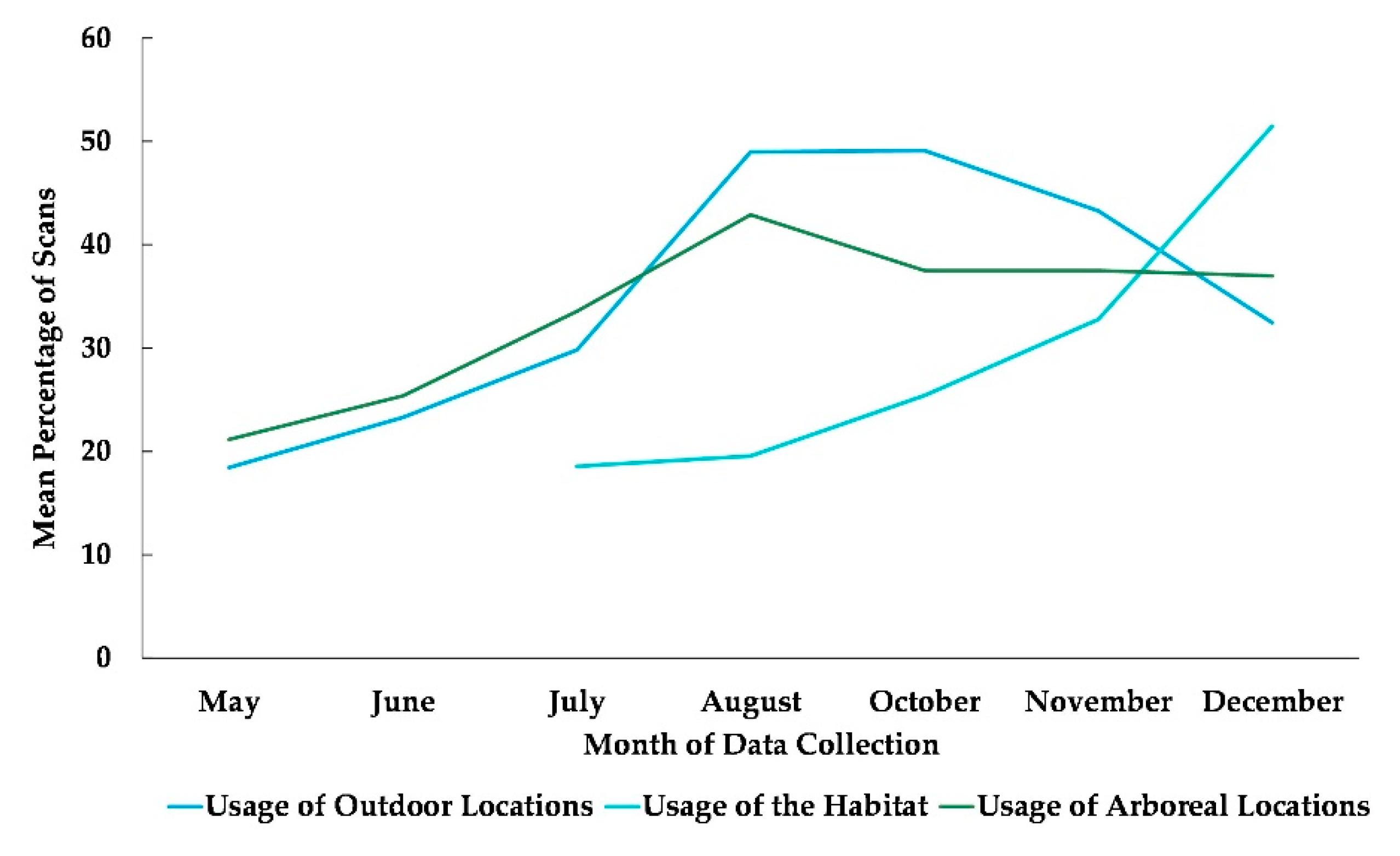
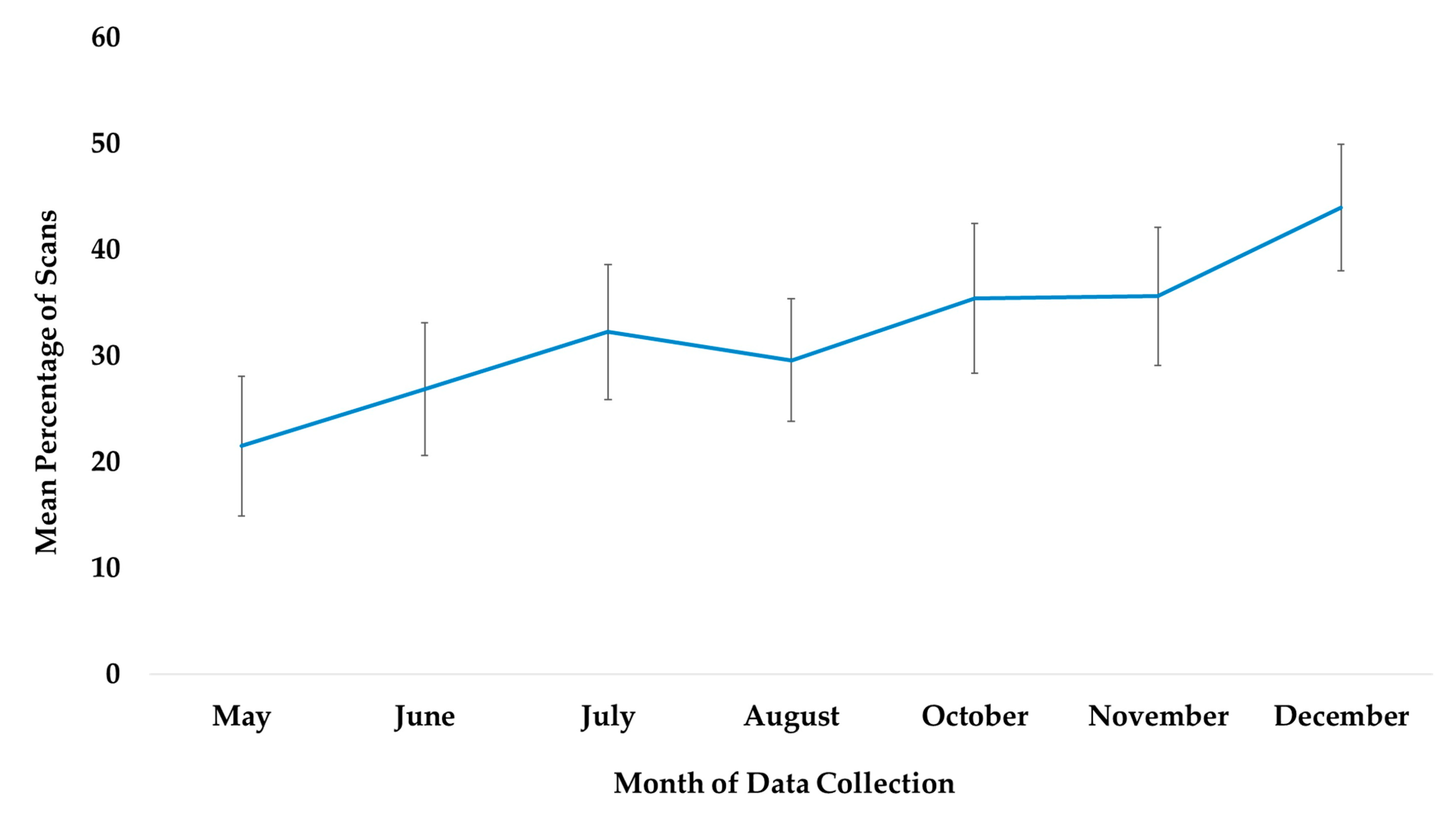
3.1. Use of All Outdoor Space
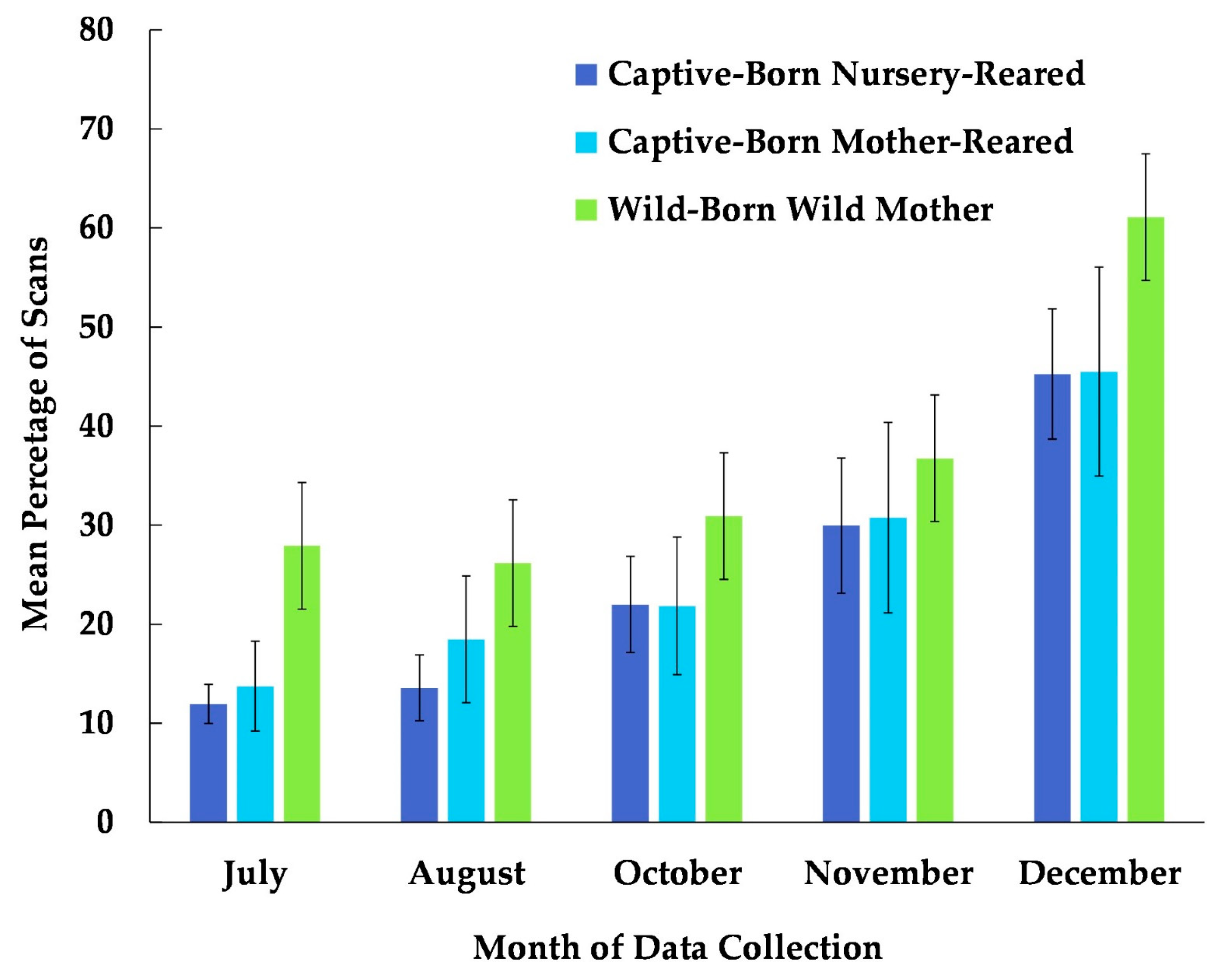
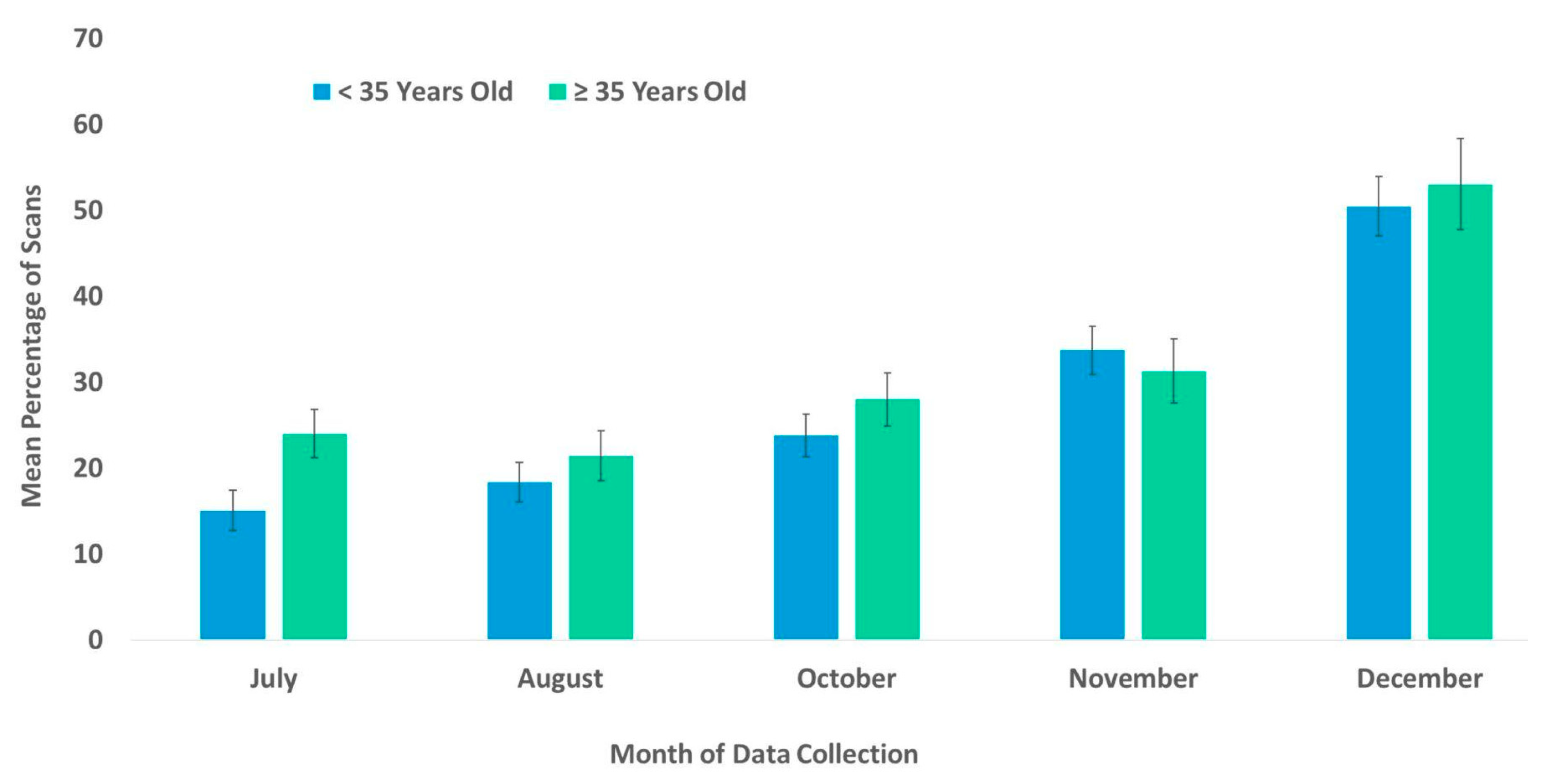
3.2. Habitat Use
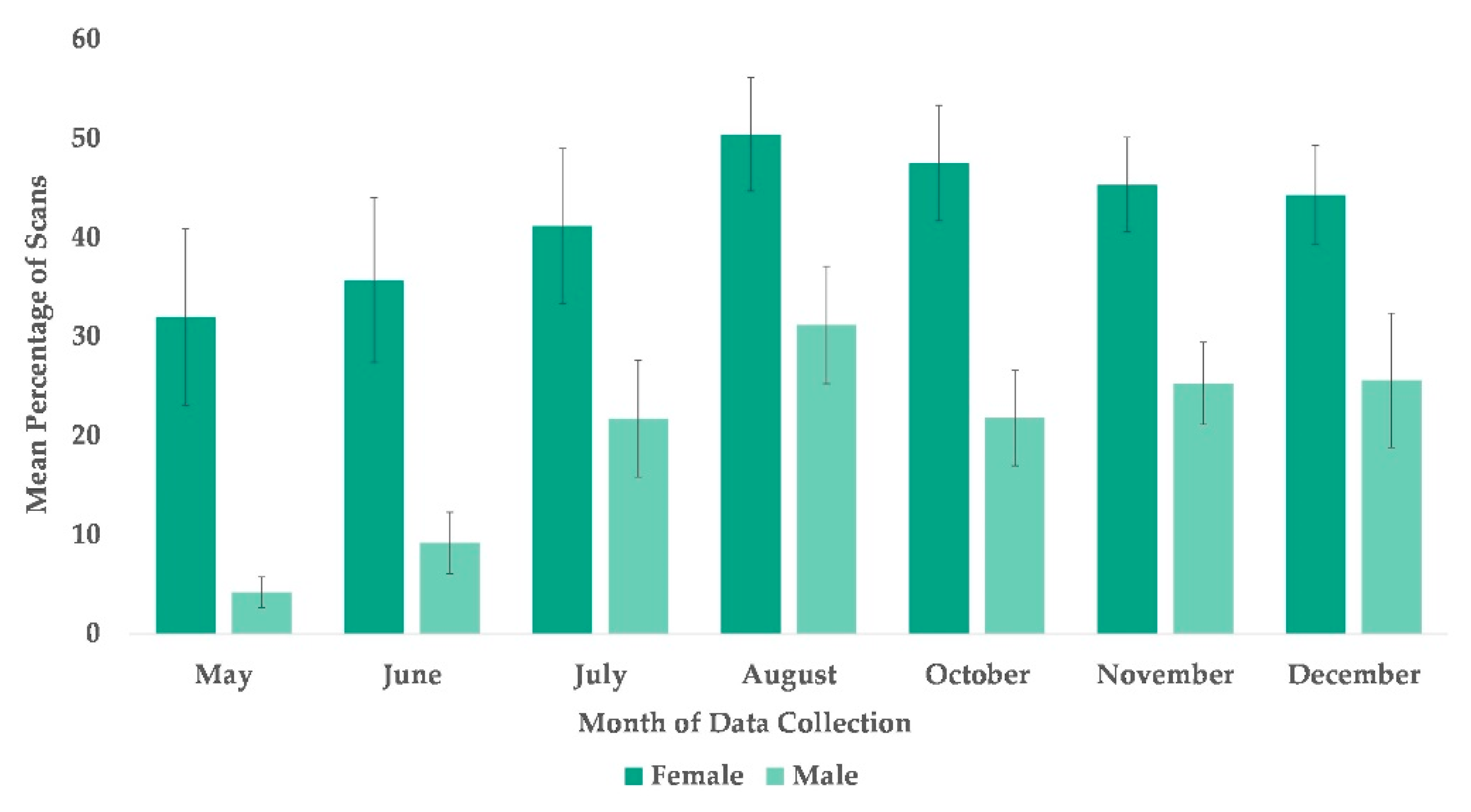
3.3. Use of Arboreal Space
3.4. Use of Indoor Arboreal Space
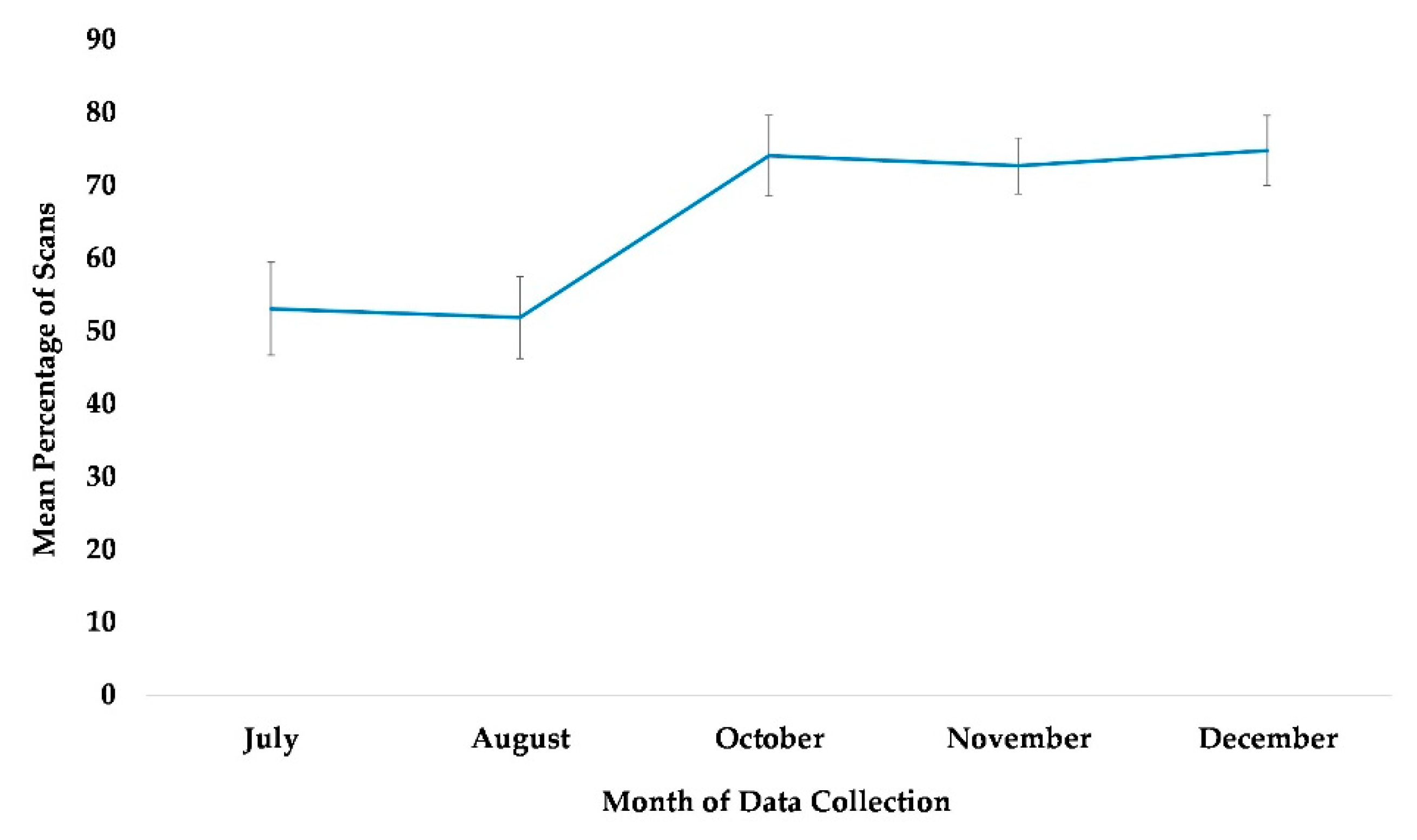
3.5. Use of Terrestrial Space (Habitat)
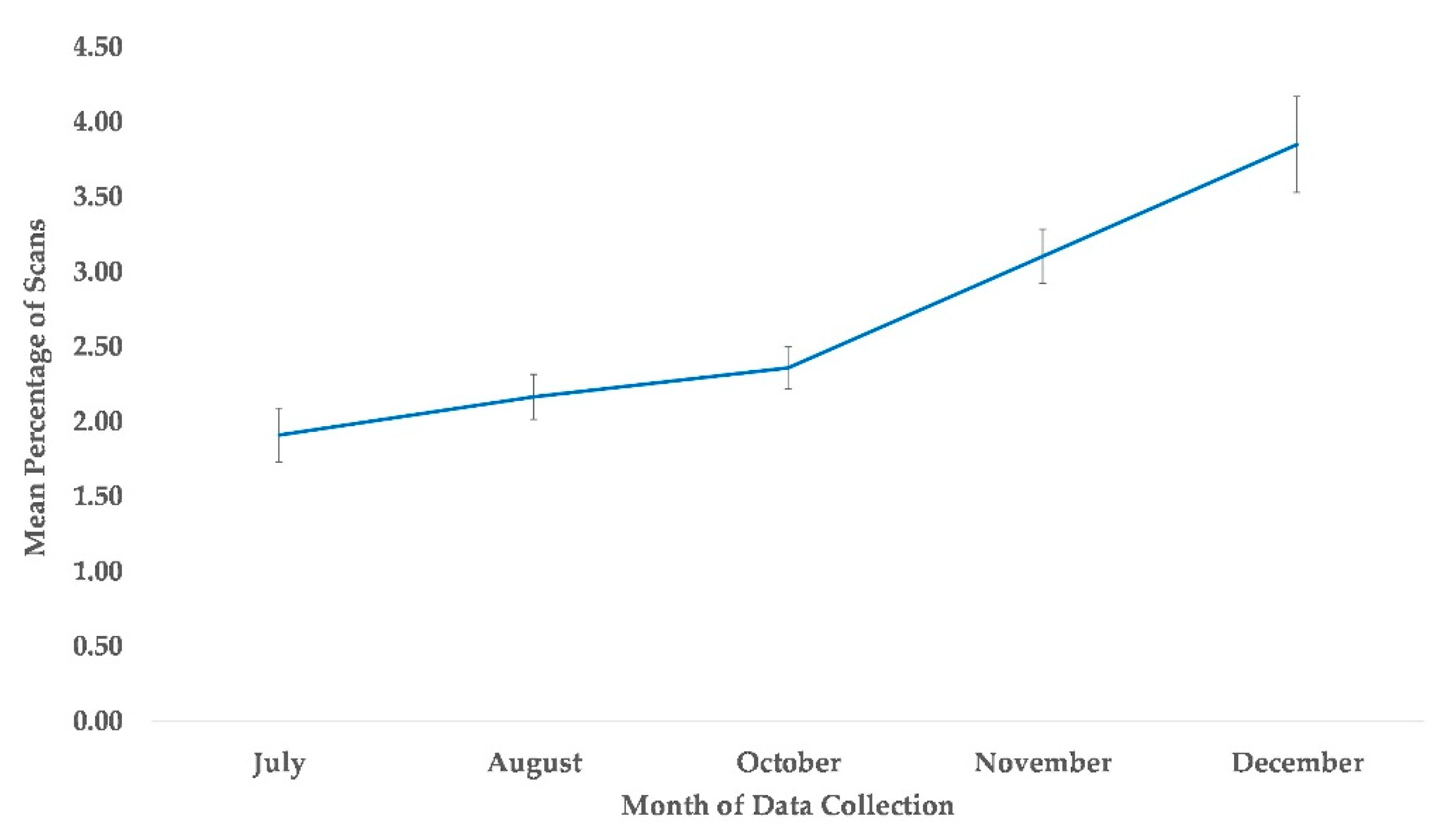
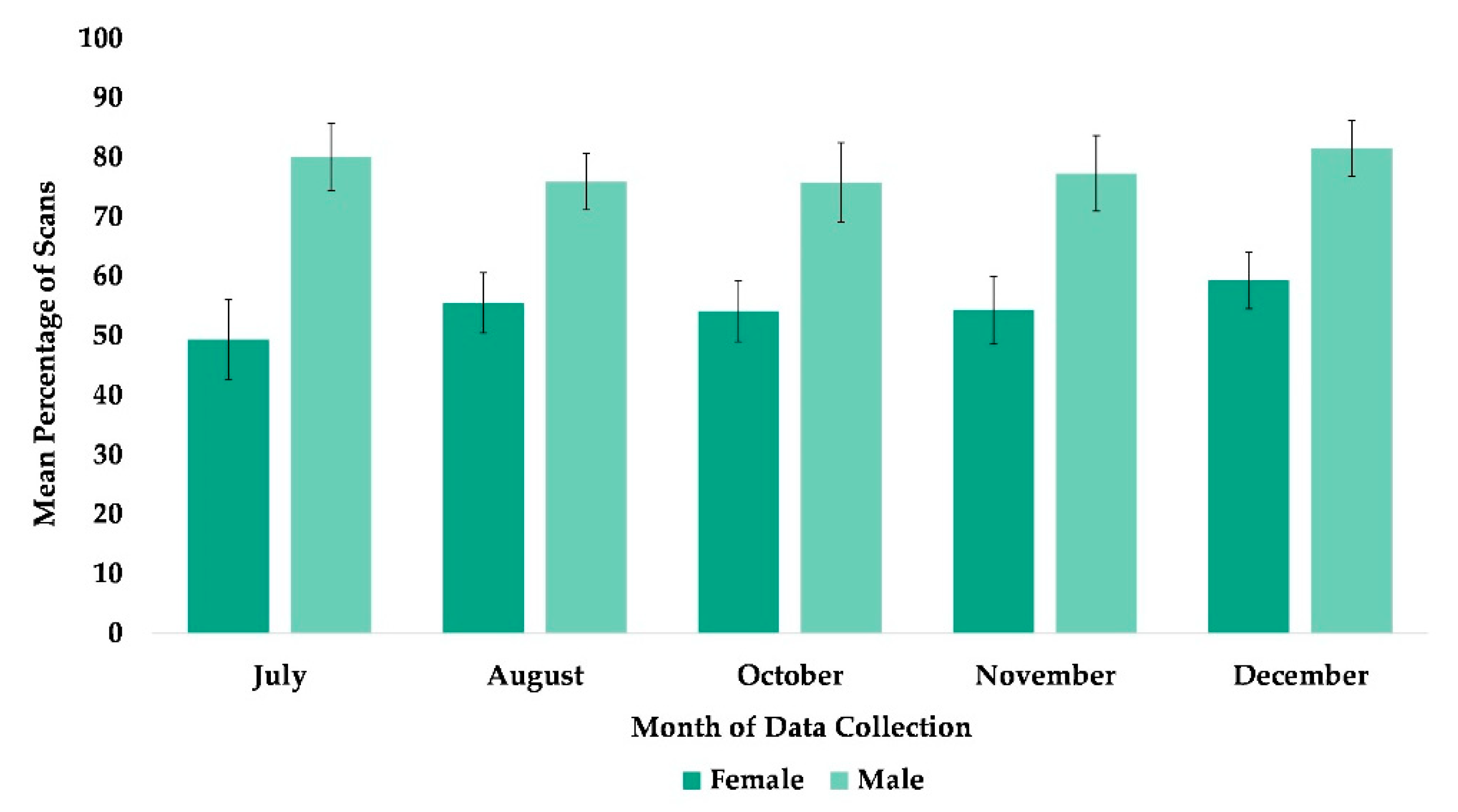
3.6. Proximity to All Group Members
3.7. Proximity to Unfamiliar Group Members
4. Discussion
4.1. Use of All Outdoor Space
4.2. Habitat Use
4.3. Use of Arboreal Space
4.4. Use of Terrestrial Space (Habitat)
4.5. Proximity to Group Members
4.6. Future Considerations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doran, D.M. Comparative positional behavior of the African apes. In Great Ape Societies; McGrew, W.C., Marchant, L.F., Nishida, T., Eds.; Cambridge University Press: Cambridge, UK, 1996; pp. 213–234. [Google Scholar]
- Nishida, T. Chimpanzees of the Lakeshore: Natural History and Culture at Mahale; Cambridge University Press: Cambridge, UK, 2012; ISBN 978-1-107-60178-9. [Google Scholar]
- Preutz, J.D.; McGrew, W.C. What does a chimpanzee need? Using behavior to guide the care of captive populations. In The Care and Management of Captive Chimpanzees, Special Topics in Primatology; Brent, L., Ed.; The American Society of Primatologists: San Antonio, TX, USA, 2001; Volume 2, pp. 16–37. [Google Scholar]
- Reynolds, V. The Chimpanzees of the Budongo Forest: Ecology, Behaviour, and Conservation; Oxford University Press: New York, NY, USA, 2005. [Google Scholar] [CrossRef]
- Stanford, C. Apes of the Impenetrable Forest: The Behavioral Ecology of Sympatric Chimpanzees and Gorillas; Pearson Prentice Hall: Hoboken, NJ, USA, 2008. [Google Scholar] [CrossRef]
- Lehmann, J.; Boesch, C. Sexual differences in chimpanzee sociality. Int. J. Primatol. 2008, 29, 65–81. [Google Scholar] [CrossRef]
- Gilby, I.C.; Wrangham, R.W. Association patterns among wild chimpanzees (Pan troglodytes schweinfurthii) reflect sex differences in cooperation. Behav. Ecol. Sociobiol. 2008, 62, 1831. [Google Scholar] [CrossRef]
- Goodall, J. The Chimpanzees of Gombe: Patterns of Behavior; Belknap Press of Harvard University Press: Cambridge, MA, USA, 1986; ISBN 0-674-11649-6. [Google Scholar]
- Mitani, J.C.; Watts, D.P.; Amsler, S.J. Lethal intergroup aggression leads to territorial expansion in wild chimpanzees. Curr. Biol. 2010, 20, R507–R508. [Google Scholar] [CrossRef] [PubMed]
- Boesch, C.; Head, J.; Tagg, N.; Arandjelovic, M.; Vigilant, L.; Robbins, M.M. Fatal chimpanzee attack in Loango National Park, Gabon. Int. J. Primatol. 2007, 28, 1025–1034. [Google Scholar] [CrossRef]
- Humle, T.; Maisels, F.; Oates, J.F.; Plumptre, A.; Williamson, E.A. Pan troglodytes; (Errata Version Published in 2018). The IUCN Red List of Threatened Species [IUCN Website], International Union for Conservation of Nature and Natural Resources. 2016. Available online: https://www.iucnredlist.org/species/15933/129038584 (accessed on 16 January 2023).
- Kosheleff, V.; Anderson, C. Temperature’s influence on the activity budget, terrestriality, and sun exposure of chimpanzees in the Budongo Forest, Uganda. Am. J. Phys. Anthropol. 2009, 139, 172–181. [Google Scholar] [CrossRef] [PubMed]
- National Research Council of the National Academies. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Animals; National Academies Press: Washington, DC, USA, 2011. Available online: https://www.ncbi.nlm.nih.gov/books/NBK54050/ (accessed on 10 January 2023). [CrossRef]
- Else, J.G. A Review of Literature and Animal Welfare/Regulatory Requirements and Guidance Pertaining to the Space Density Needs of Captive Research Chimpanzees. Unpublished Manuscript. 2013. Available online: https://dpcpsi.nih.gov/sites/default/files/ElseLitReviewFinal-110713.pdf (accessed on 12 January 2023).
- Ross, S.R.; Calcutt, S.; Schapiro, S.J.; Hau, J. Space use selectivity by chimpanzees and gorillas in an indoor-outdoor enclosure. Am. J. Primatol. 2011, 73, 197–208. [Google Scholar] [CrossRef]
- Jensvold, M.L.A.; Sanz, C.M.; Fouts, R.S.; Fouts, D.H. Effect of enclosure size and complexity on the behaviors of captive chimpanzees (Pan troglodytes). J. Appl. Anim. Welf. Sci. 2001, 4, 53–69. [Google Scholar] [CrossRef]
- Ross, S.R.; Shender, M.A. Daily travel distances of zoo-housed chimpanzees and gorillas: Implications for welfare assessments and space requirements. Primates 2016, 57, 395–401. [Google Scholar] [CrossRef]
- Wehnelt, S.; Bird, S.; Lenihan, A. Chimpanzee forest exhibit at Chester Zoo. Int. Zoo Yearb. 2006, 40, 313–322. [Google Scholar] [CrossRef]
- Ross, S.R.; Wagner, K.E.; Schapiro, S.J.; Hau, J. Ape behavior in two alternating environments: Comparing exhibit and short-term holding areas. Am. J. Primatol. 2010, 72, 951–959. [Google Scholar] [CrossRef]
- Brent, L.; Fultz, A.; DelaHaya, R.; Bettinger, T. Acclimation of captive chimpanzees to a 5- acre wooded habitat at Chimp Haven: Expression of natural behaviors. Int. J. Primatol. 2006, 27, 56. [Google Scholar]
- Fultz, A.; Brent, L. Emergence of natural behaviors in sanctuary chimpanzees (Pan troglodytes). Am. J. Primatol. 2008, 70 (Suppl. 1), 45. [Google Scholar]
- Fultz, A.; Brent, L.; Breaux, S.D.; Grand, A.P. An evaluation of nest-building behavior by sanctuary chimpanzees with access to forested habitats. Folia Primatol. 2013, 84, 405–420. [Google Scholar] [CrossRef] [PubMed]
- Ross, S.R.; Wagner, K.E.; Schapiro, S.J.; Hau, J.; Lukas, K.E. Transfer and acclimatization effects on the behavior of two species of African great ape (Pan troglodytes and Gorilla gorilla gorilla) moved to a novel and naturalistic zoo environment. Int. J. Primatol. 2011, 32, 99–117. [Google Scholar] [CrossRef]
- Videan, E.N.; Fritz, J. Effects of short- and long-term changes in spatial density on the social behavior of captive chimpanzees (Pan troglodytes). J. Appl. Anim. Welf. Sci. 2007, 102, 95–105. [Google Scholar] [CrossRef]
- Hosey, G.R. How does the zoo environment affect the behavior of captive primates? J. Appl. Anim. Welf. Sci. 2005, 90, 107–129. [Google Scholar] [CrossRef]
- Perkins, L.A. Variables that influence the activity of captive orangutans. Zoo Biol. 1992, 11, 177–186. [Google Scholar] [CrossRef]
- Ross, S.R.; Lukas, K.E. Use of space in a non-naturalistic environment by chimpanzees (Pan troglodytes) and lowland gorillas (Gorilla gorilla gorilla). Appl. Anim. Behav. Sci. 2006, 96, 143–152. [Google Scholar] [CrossRef]
- Wilson, S.F. Environmental influences on the activity of captive apes. Zoo Biol. 1982, 1, 201–209. [Google Scholar] [CrossRef]
- Brent, L.; Lee, R.D.; Eichberg, J.W. Evaluation of a chimpanzee enrichment enclosure. J. Med. Primatol. 1991, 20, 29–34. [Google Scholar] [CrossRef]
- Goff, C.; Howell, S.M.; Fritz, J.; Nankivell, B. Space use and proximity of captive chimpanzee (Pan troglodytes) mother/offspring pairs. Zoo Biol. 1994, 13, 61–68. [Google Scholar] [CrossRef]
- Murray, C.M.; Mane, S.V.; Pusey, A.E. Dominance rank influences female space use in wild chimpanzees, Pan troglodytes: Towards an ideal despotic distribution. Anim. Behav. 2007, 74, 1795–1804. [Google Scholar] [CrossRef]
- Doran, D.M.; Hunt, K.D. Comparative locomotor behavior of chimpanzees and bonobos: Species and habitat differences. In Chimpanzee Cultures; Wrangham, R.W., McGrew, W.C., de Waal, F.B.M., Heltne, P.G., Eds.; Chicago Academy of Sciences: Chicago, IL, USA, 1994; pp. 93–108. [Google Scholar]
- Clark, F.E. Space to choose: Network analysis of social preferences in a captive chimpanzee community, and implications for management. Am. J. Primatol. 2011, 73, 748–757. [Google Scholar] [CrossRef]
- Ross, S.R.; Schapiro, S.J.; Hau, J.; Lukas, K.E. Space use as an indicator of enclosure appropriateness: A novel measure of captive animal welfare. Appl. Anim. Behav. Sci. 2009, 121, 42–50. [Google Scholar] [CrossRef]
- Sherwen, S.L.; Hemsworth, P.H. The Visitor Effect on Zoo Animals: Implications and Opportunities for Zoo Animal Welfare. Animals 2019, 9, 366. [Google Scholar] [CrossRef]
- Curry, B.A.; Drane, A.L.; Atencia, R.; Feltrer, Y.; Howatson, G.; Calvi, T.; Palmer, C.; Moittie, S.; Unwin, S.; Tremblay, J.C.; et al. Body mass and growth rates in captive chimpanzees (Pan troglodytes) cared for in African wildlife sanctuaries, zoological institutions, and research facilities. Zoo Biol. 2023, 42, 98–106. [Google Scholar] [CrossRef]
- Leigh, S.R.; Shea, B.T. Ontogeny of body size variation in African apes. Am. J. Phys. Anthropol. 1996, 99, 43–65. [Google Scholar] [CrossRef]
- Uehara, S.; Nishida, T. Body weights of wild chimpanzees (Pan troglodytes schweinfurthii) of the Mahale Mountains National Park, Tanzania. Am. J. Phys. Anthropol. 1987, 72, 315–321. [Google Scholar] [CrossRef]
- Fultz, A.; Panu, L.D.; Landon, F.; Brent, L. The preliminary response of retired captive chimpanzees to increases in enclosure and group size. Inter. J. Primatol. 2006, 27, 38. [Google Scholar]
- Fultz, A.; Yanagi, A.; Breaux, S.; Beaupre, L. Aggressive, submissive, and affiliative behavior in sanctuary chimpanzees (Pan troglodytes) during social integration. Animals 2002, 12, 2421. [Google Scholar] [CrossRef]
- Hellaye, Y.L.; Goossens, B.; Jamart, A.; Curtis, D.J. Acquisition of fission-fusion social organization in a chimpanzee (Pan troglodytes troglodytes) community released into the wild. Behav. Ecol. Sociobiol. 2010, 64, 349–360. [Google Scholar] [CrossRef]
- Pruetz, J.D.; Fulton, S.J.; Marchant, L.F.; McGrew, W.C.; Schiel, M.; Waller, M. Arboreal nesting as antipredator adaptation by savanna chimpanzees (Pan troglodytes verus) in southeastern Senegal. Am. J. Primatol. 2008, 70, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Clay, A.W.; Ross, S.R.; Lambeth, S.; Vazquez, M.; Breaux, S.; Pietsch, R.; Fultz, A.; Lammey, M.; Jacobson, S.L.; Perlman, J.E.; et al. Chimpanzees (Pan troglodytes) in U.S. Zoos, Sanctuaries, and Research Facilities: A Survey-Based Comparison of Species-Typical Behaviors. Animals 2023, 13, 251. [Google Scholar] [CrossRef] [PubMed]
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 49, 227–266. [Google Scholar] [CrossRef]
- Laurence, H.; Kumar, S.; Owston, M.A.; Lanford, R.E.; Hubbard, G.B.; Dick, E.J., Jr. Natural mortality and cause of death analysis of the captive chimpanzee (Pan troglodytes): A 35-year review. J. Med. Primatol. 2017, 46, 106–115. [Google Scholar] [CrossRef]
- Che-Castaldo, J.; Havercamp, K.; Watanuki, K.; Matsuzawa, T.; Hirata, S.; Ross, S.R. Comparative survival analyses among captive chimpanzees (Pan troglodytes) in America and Japan. PeerJ 2021, 9, e11913. [Google Scholar] [CrossRef]
- Neal Webb, S.J.; Hau, J.; Lambeth, S.P.; Schapiro, S.J. Differences in behavior between elderly and nonelderly captive chimpanzees and the effects of the social environment. J. Am. Assoc. Lab. Anim. Sci. 2019, 58, 783–789. [Google Scholar] [CrossRef]
- Ross, S.R.; Joshi, P.B.; Terio, K.A.; Gamble, K.C. A 25-year retrospective review of mortality in chimpanzees (Pan troglodytes) in accredited U.S. zoos from a management and welfare perspective. Animals 2022, 12, 1878. [Google Scholar] [CrossRef]
- Duncan, L.M.; D’Egidio Kotze, C.; Pillay, N. Long-Term Spatial Restriction Generates Deferred Limited Space Use in a Zoo-Housed Chimpanzee Group. Animals 2022, 12, 2207. [Google Scholar] [CrossRef]
- Bloomsmith, M.A.; Lambeth, S.P.; Haberstroh, M.D. Chimpanzee use of enclosures. Am. J. Primatol. 1999, 49, 36. [Google Scholar]
- Alp, R. Meat eating and ant dipping by wild chimpanzees in Sierra Leone. Primates 1993, 34, 463–468. [Google Scholar] [CrossRef]
- Pruetz, J.D.; Bertolani, P. Chimpanzee (Pan troglodytes verus) behavioral responses to stresses associated with living in a savanna-mosaic environment: Implications for hominin adaptations to open habitats. PaleoAnthropology 2009, 2009, 252–262. [Google Scholar] [CrossRef]
- Herrelko, E.; Buchanan-Smith, H.; Vick, S. Perception of available space during chimpanzee introductions: Number of accessible areas is more important than enclosure size. Zoo Biol. 2015, 34, 397–405. [Google Scholar] [CrossRef]
- National Institutes of Health. Costs for Maintaining Humane Care and Welfare of Chimpanzees. Available online: https://orip.nih.gov/comparative-medicine/programs/vertebrate-models/chimpanzee-management-reports (accessed on 10 January 2023).
- Nunamaker, E.A.; Lee, D.R.; Lammey, M.L. Chronic diseases in captive geriatric female chimpanzees (Pan troglodytes). Comp. Med. 2012, 62, 131–136. [Google Scholar]
- Koops, K.; McGrew, W.C.; DeVries, H.; Matsuzawa, T. Nest-building by chimpanzees (Pan troglodytes verus) at Seringbara, Nimba Mountains: Anti predation, thermoregulation, and antivector hypotheses. Int. J. Primatol. 2012, 33, 356–380. [Google Scholar] [CrossRef]
- Anderson, J.R. Chimpanzees and death. Phil. Trans. R. Soc. 2018, B373, 20170257. [Google Scholar] [CrossRef]
- Bloomsmith, M.A.; Clay, A.W.; Ross, S.R.; Lambeth, S.P.; Lutz, C.K.; Breaux, S.D.; Pietsch, R.; Fultz, A.; Lammey, M.L.; Jacobson, S.L.; et al. Chimpanzees in U.S. zoos, sanctuaries, and research facilities: A survey-based comparison of atypical behaviors. In Chimpanzees in Context: A Comparative Perspective on Chimpanzee Behavior, Cognition, Conservation and Welfare; Hopper, L.M., Ross, S.R., Eds.; University of Chicago Press: Chicago, IL, USA, 2020. [Google Scholar]
- Persad-Clem, R. Adaptation of Captive Chimpanzees (Pan troglodytes) to Free Ranging in a Natural Temperate Environment. Doctoral Dissertation, Miami University, Oxford, OH, USA, 2009. Available online: http://rave.ohiolink.edu/etdc/view?acc_num=miami1250172406/ (accessed on 24 November 2022).
- Neal Webb, S.J.; Hau, J.; Schapiro, S.J. Captive chimpanzee (Pan troglodytes) behavior as a function of space per animal and enclosure type. Am. J. Primatol. 2018, 80, e22749. [Google Scholar] [CrossRef]
- Binding, S.; Farmer, H.; Krusin, L.; Cronin, K. Status of animal welfare research in zoos and aquariums: Where are we, where to next? JZAR 2020, 8, 166–174. [Google Scholar] [CrossRef]
- Ross, S.R. Chimpanzee Welfare in the Context of Science, Policy, and Practice. In Chimpanzees in Context; Hopper, L.M., Ross, S.R., Eds.; University of Chicago Press: Chicago, IL, USA, 2020; pp. 552–582. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chimpanzee | Sex | Age | Rearing History |
|---|---|---|---|
| Aug | Female | 30 | Captive-Born Nursery-Reared |
| Con | Male | 19 | Captive-Born Mother-Reared |
| Dee | Female | 19 | Wild-Born Wild Mother |
| Dea | Female | 38 | Wild-Born Wild Mother |
| Ell | Female | 33 | Captive-Born Mother-Reared |
| Gya | Female | 38 | Wild-Born Wild Mother |
| Joh | Male | 41 | Wild-Born Wild Mother |
| Jun | Male | 37 | Captive-Born Nursery-Reared |
| Lta | Female | 46 | Wild-Born Wild Mother |
| Mgn | Male | 16 | Captive-Born Nursery-Reared |
| Msn | Male | 21 | Captive-Born Nursery-Reared |
| Mrv | Male | 29 | Captive-Born Nursery-Reared |
| Pdn | Male | 26 | Captive-Born Nursery-Reared |
| Rta | Female | 41 | Wild-Born Wild Mother |
| Sra | Female | 20 | Captive-Born Mother-Reared |
| Shi | Female | 33 | Captive-Born Mother-Reared |
| Sza | Female | 29 | Captive-Born Nursery-Reared |
| Tra | Female | 41 | Wild-Born Wild Mother |
| Prediction | Factor | Factor Level b | Estimate | S.E. | Z | p Value | Lower Ci c | Upper Ci c |
|---|---|---|---|---|---|---|---|---|
| Use of all outdoor space | Time | May | Reference | |||||
| June * | 0.349 | 0.164 | 2.13 | 0.0330 | 0.028 | 0.670 | ||
| July *** | 0.705 | 0.16 | 4.4 | 0.0000 | 0.391 | 1.020 | ||
| August *** | 1.551 | 0.155 | 10.03 | 0.0000 | 1.248 | 1.855 | ||
| October *** | 1.566 | 0.157 | 9.99 | 0.0000 | 1.259 | 1.873 | ||
| November *** | 1.322 | 0.158 | 8.36 | 0.0000 | 1.012 | 1.632 | ||
| December *** | 0.838 | 0.162 | 5.17 | 0.0000 | 0.521 | 1.156 | ||
| Habitat use | Time | July | Reference | |||||
| August | 0.0916 | 0.144 | 0.64 | 0.5245 | −0.190 | 0.374 | ||
| October ** | 0.4401 | 0.14 | 3.15 | 0.0016 | 0.167 | 0.714 | ||
| November *** | 0.8047 | 0.136 | 5.91 | 0.0000 | 0.538 | 1.072 | ||
| December *** | 1.623 | 0.135 | 12.03 | 0.0000 | 1.359 | 1.888 | ||
| Age | (Continuous) * | −0.0396 | 0.018 | −2.27 | 0.0235 | −0.074 | −0.005 | |
| Rearing History | Captive-Born Mother-Reared | Reference | ||||||
| Captive-Born Nursery-Reared | −0.022 | 0.332 | −0.07 | 0.9470 | −0.672 | 0.628 | ||
| Wild-Born Wild-Mother-Reared ** | 1.1175 | 0.388 | 2.88 | 0.0040 | 0.357 | 1.878 | ||
| Use of all arboreal space | Time | May | Reference | |||||
| June | 0.327 | 0.19 | 1.72 | 0.0853 | −0.045 | 0.699 | ||
| July *** | 0.8 | 0.189 | 4.24 | 0.0000 | 0.430 | 1.170 | ||
| August *** | 1.282 | 0.19 | 6.75 | 0.0000 | 0.910 | 1.654 | ||
| October *** | 1.02 | 0.189 | 5.39 | 0.0000 | 0.649 | 1.391 | ||
| November *** | 1.032 | 0.193 | 5.36 | 0.0000 | 0.654 | 1.409 | ||
| December *** | 0.968 | 0.194 | 4.99 | 0.0000 | 0.588 | 1.348 | ||
| Sex | Female | Reference | ||||||
| Male ** | −1.103 | 0.359 | −3.07 | 0.0021 | −1.807 | −0.400 | ||
| Use of indoor arboreal space | Time | May | Reference | |||||
| June | 0.417 | 0.232 | 1.79 | 0.0729 | −0.039 | 0.872 | ||
| July ** | 0.741 | 0.231 | 3.2 | 0.0014 | 0.288 | 1.195 | ||
| August ** | 0.625 | 0.233 | 2.68 | 0.0074 | 0.168 | 1.082 | ||
| October *** | 0.936 | 0.224 | 4.17 | 0.0000 | 0.497 | 1.376 | ||
| November *** | 0.957 | 0.229 | 4.17 | 0.0000 | 0.507 | 1.406 | ||
| December *** | 1.354 | 0.234 | 5.8 | 0.0000 | 0.897 | 1.812 | ||
| Sex | Female | Reference | ||||||
| Male * | −1.042 | 0.49 | −2.13 | 0.0333 | −2.002 | −0.083 | ||
| Use of terrestrial space (habitat) | Time | July | Reference | |||||
| August | −0.114 | 0.255 | −0.45 | 0.6553 | −0.613 | 0.385 | ||
| October *** | 0.94 | 0.265 | 3.55 | 0.0004 | 0.421 | 1.459 | ||
| November ** | 0.884 | 0.276 | 3.2 | 0.0014 | 0.342 | 1.425 | ||
| December *** | 1.048 | 0.279 | 3.76 | 0.0002 | 0.501 | 1.594 | ||
| Proximity to all group members | Time | July | Reference | |||||
| August | 0.176 | 0.114 | 1.55 | 0.1220 | −0.047 | 0.399 | ||
| October | 0.189 | 0.114 | 1.66 | 0.0960 | −0.034 | 0.411 | ||
| November *** | 0.493 | 0.107 | 4.6 | 0.0000 | 0.283 | 0.703 | ||
| December *** | 0.717 | 0.104 | 6.93 | 0.0000 | 0.514 | 0.920 | ||
| Sex | Female | Reference | ||||||
| Male * | −0.247 | 0.101 | −2.46 | 0.0140 | −0.444 | −0.050 | ||
| Proximity to unfamiliar group members | Time | July | Reference | |||||
| August | 0.086 | 0.132 | 0.65 | 0.5130 | −0.172 | 0.344 | ||
| October | 0.039 | 0.131 | 0.29 | 0.7680 | −0.217 | 0.294 | ||
| November | 0.077 | 0.13 | 0.59 | 0.5519 | −0.178 | 0.332 | ||
| December * | 0.31 | 0.133 | 2.33 | 0.0198 | 0.049 | 0.570 | ||
| Sex | Female | Reference | ||||||
| Male *** | 1.186 | 0.351 | 3.38 | 0.0007 | 0.499 | 1.874 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fultz, A.; Yanagi, A.; Breaux, S.; Beaupre, L.; Naitove, N. How Sanctuary Chimpanzees (Pan troglodytes) Use Space after Being Introduced to a Large Outdoor Habitat. Animals 2023, 13, 961. https://doi.org/10.3390/ani13060961
Fultz A, Yanagi A, Breaux S, Beaupre L, Naitove N. How Sanctuary Chimpanzees (Pan troglodytes) Use Space after Being Introduced to a Large Outdoor Habitat. Animals. 2023; 13(6):961. https://doi.org/10.3390/ani13060961
Chicago/Turabian StyleFultz, Amy, Akie Yanagi, Sarah Breaux, Leilani Beaupre, and Nick Naitove. 2023. "How Sanctuary Chimpanzees (Pan troglodytes) Use Space after Being Introduced to a Large Outdoor Habitat" Animals 13, no. 6: 961. https://doi.org/10.3390/ani13060961
APA StyleFultz, A., Yanagi, A., Breaux, S., Beaupre, L., & Naitove, N. (2023). How Sanctuary Chimpanzees (Pan troglodytes) Use Space after Being Introduced to a Large Outdoor Habitat. Animals, 13(6), 961. https://doi.org/10.3390/ani13060961