
A One-Year Retrospective Analysis of Viral and Parasitological Agents in Wildlife Animals Admitted to a First Aid Hospital

 , , ,
, , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
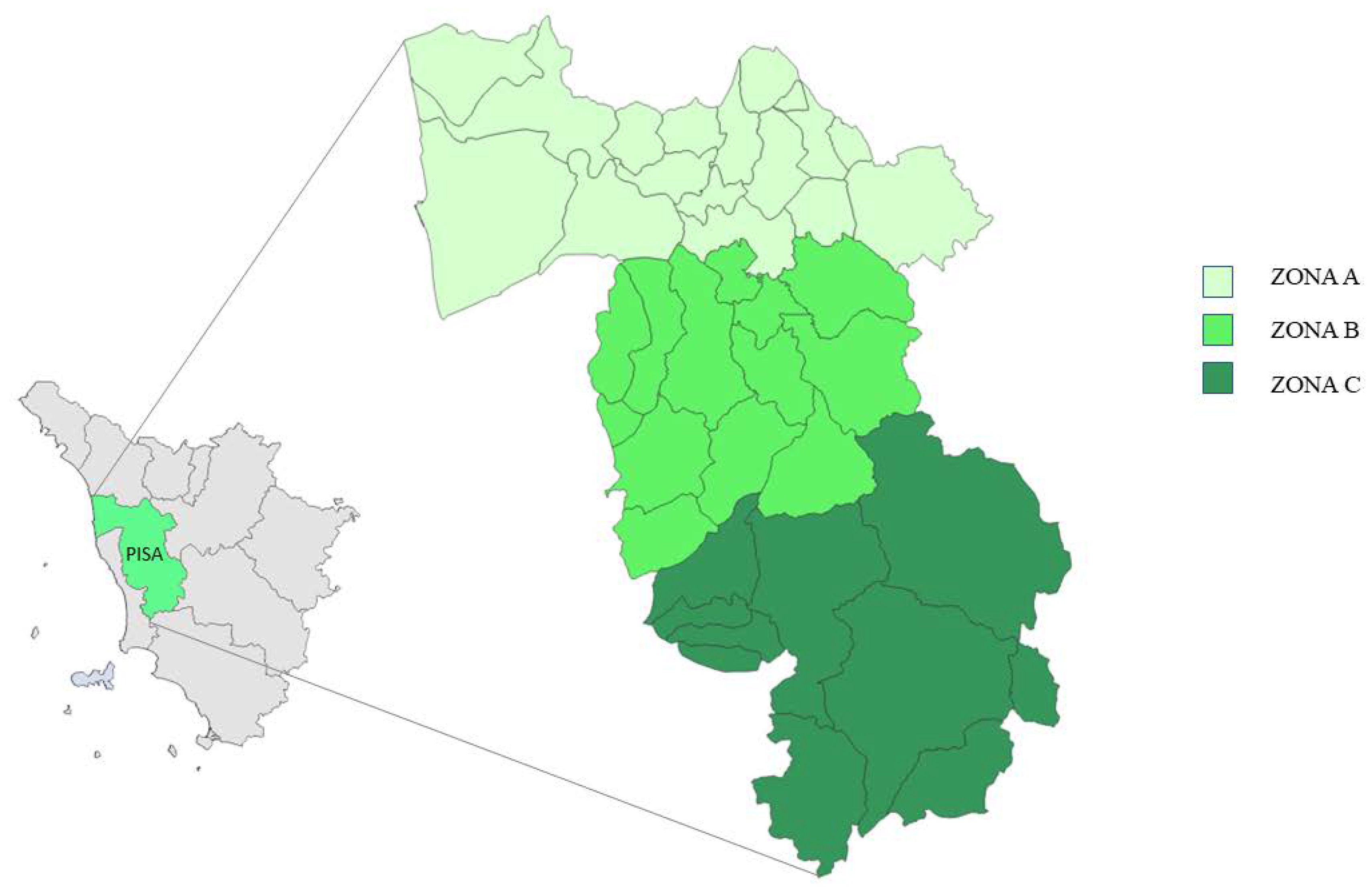
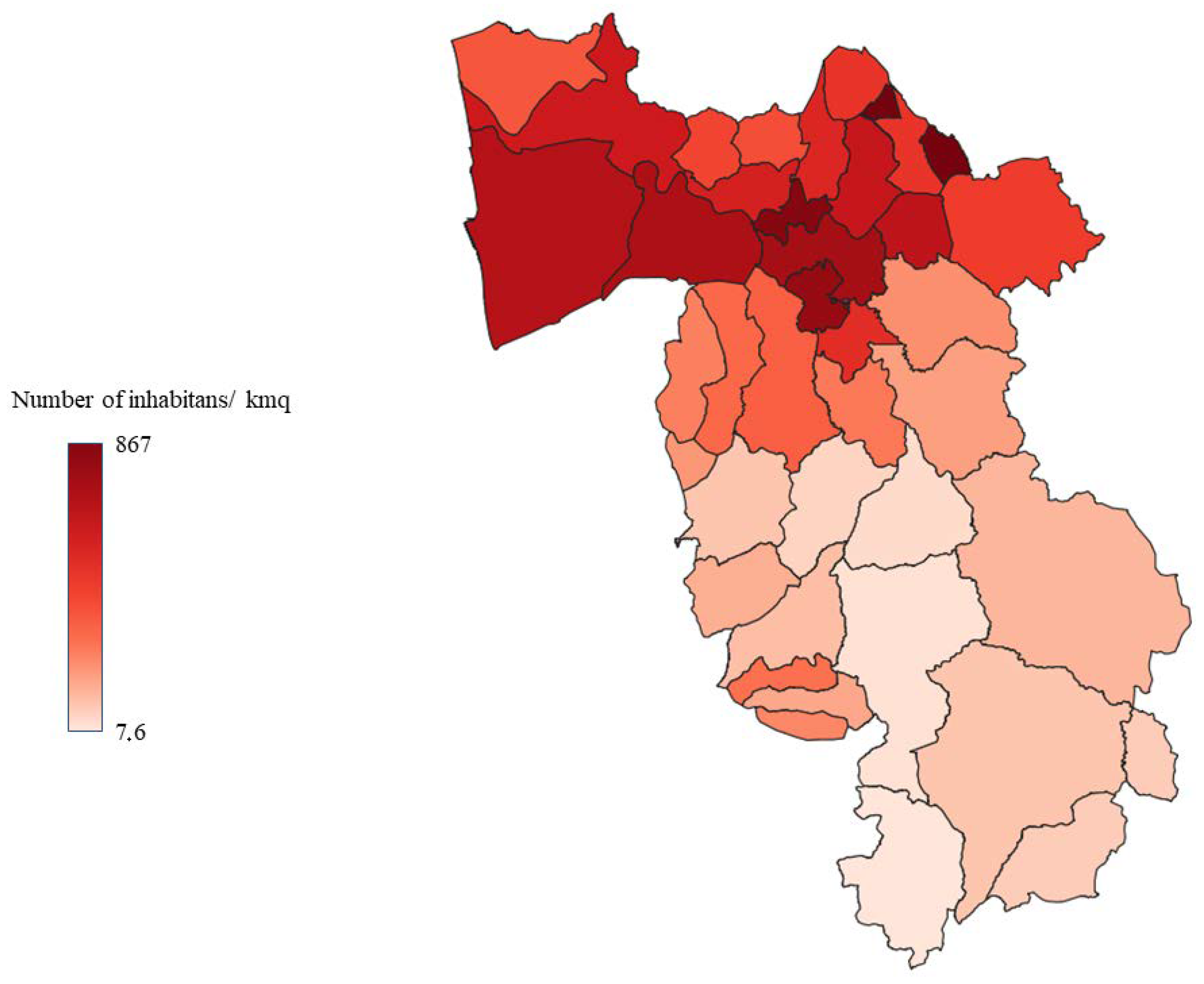
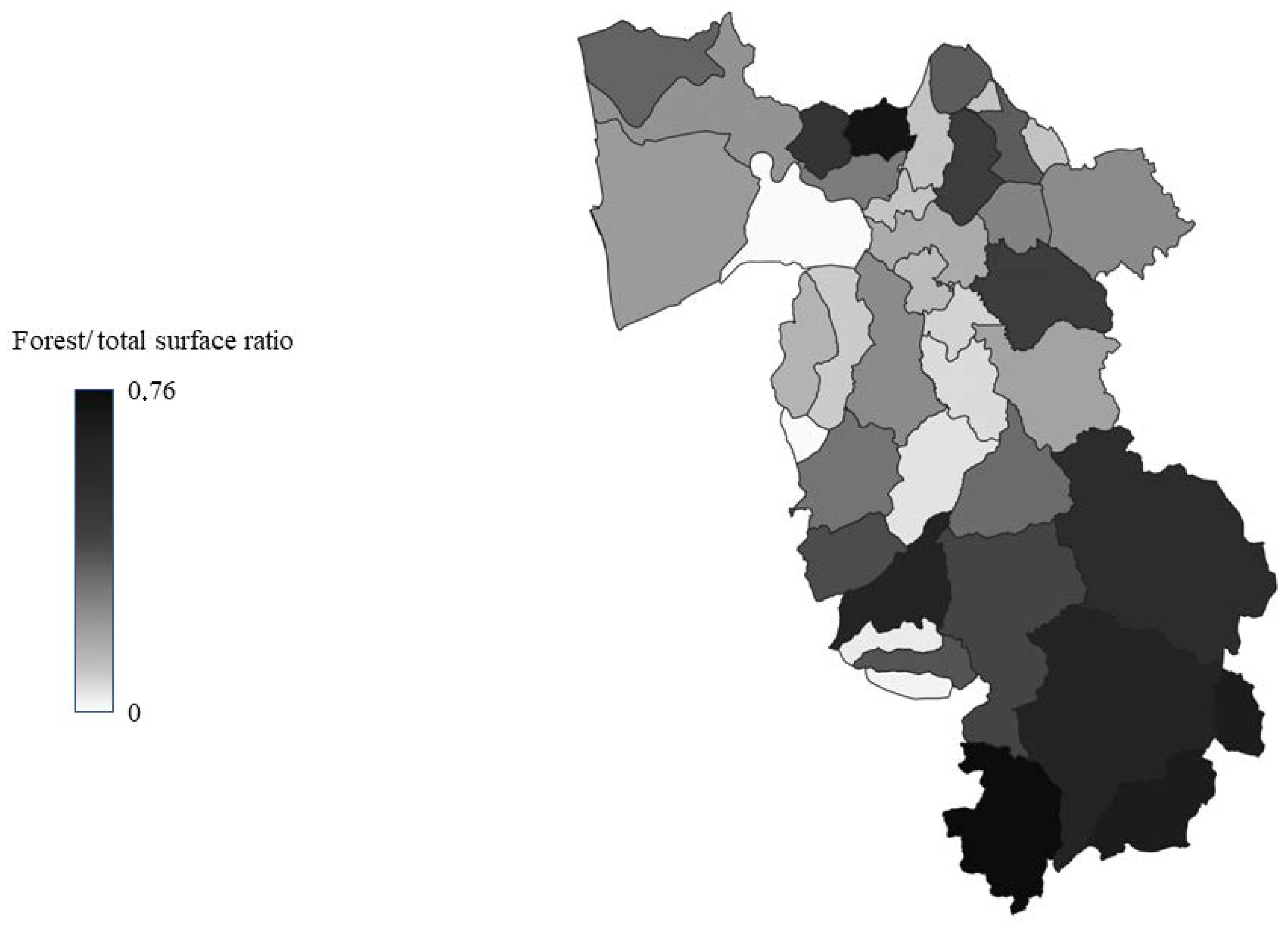
2.1. Study Area
2.2. Enrolled Animals
2.3. Sample Collection
2.4. Viral Pathogens
2.4.1. Serological Analysis
2.4.2. Molecular Analysis
2.5. Parasitological Investigations
2.5.1. Faecal Examinations
2.5.2. Commercial Assays
2.5.3. Transtracheal Wash Technique
2.5.4. Molecular Analysis for Giardia duodenalis Genotyping and Cryptosporidium Species Identification
2.5.5. Molecular Identification of Adult Lungworms
2.6. Statistical Analysis
3. Results
3.1. Viral Pathogens
3.1.1. Serological Analysis
3.1.2. Molecular Examination
3.2. Parasitological Investigations
3.2.1. Faecal Examinations
3.2.2. Commercial Assays
3.2.3. TTW Results
3.2.4. Molecular Identification of Giardia duodenalis and Cryptosporidium spp.
3.2.5. Molecular Identification of Adult Lungworms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging Infectious Diseases of Wildlife—Threats to Biodiversity and Human Health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef]
- Dobson, A.; Foufopoulos, J. Emerging Infectious Pathogens of Wildlife. Philos. Trans. R. Soc. B Biol. Sci. 2001, 356, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.A. A Walk on the Wild Side—Emerging Wildlife Diseases. BMJ 2005, 331, 1214–1215. [Google Scholar] [CrossRef]
- Cleaveland, S.; Haydon, D.T.; Taylor, L. Overviews of Pathogen Emergence: Which Pathogens Emerge, When and Why? Curr. Top. Microbiol. Immunol. 2007, 315, 85–111. [Google Scholar]
- Artois, M.; Delahay, R.; Guberti, V.; Cheeseman, C. Control of Infectious Diseases of Wildlife in Europe. Vet. J. 2001, 162, 141–152. [Google Scholar] [CrossRef]
- Morner, T.; Obendorf, D.L.; Artois, M.; Woodford, M.H. Diseases of Wildlife Occur in Many Different Forms in a Wide Range of Animal Species and Populations. Rev. Sci. Tech. 2002, 21, 67–76. [Google Scholar] [CrossRef]
- Artois, M.; Bengis, R.; Delahay, R.J.; Duchêne, M.J.; Duff, J.P.; Ferroglio, E.; Gortazar, C.; Hutchings, M.R.; Kock, R.A.; Leighton, F.A.; et al. Wildlife Disease Surveillance and Monitoring. In Management of Disease in Wild Mammals; Springer: Berlin/Heidelberg, Germany, 2009; pp. 187–213. [Google Scholar] [CrossRef]
- Bengis, R.G.; Leighton, F.A.; Fischer, J.R.; Artois, M.; Mörner, T.; Tate, C.M. The Role of Wildlife in Emerging and Re-Emerging Zoonoses. OIE Rev. Sci. Tech. 2004, 23, 497–511. [Google Scholar] [CrossRef]
- Ryser-Degiorgis, M.P. Wildlife Health Investigations: Needs, Challenges and Recommendations. BMC Vet. Res. 2013, 9, 223. [Google Scholar] [CrossRef]
- Wobeser, G.A. Disease in Wild Animals: Investigation and Management; Springer: Berlin/Heidelberg, Germany, 2007; pp. 1–393. [Google Scholar] [CrossRef]
- Stitt, T.; Mountifield, J.; Stephen, C. Opportunities and Obstacles to Collecting Wildlife Disease Data for Public Health Purposes: Results of a Pilot Study on Vancouver Island, British Columbia. Can. Vet. J. 2007, 48, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Sleeman, J.M. Use of Wildlife Rehabilitation Centers as Monitors of Ecosystem Health. In Zoo and Wild Animal; Elsevier: Amsterdam, The Netherlands, 2008; pp. 97–104. [Google Scholar] [CrossRef]
- Trocini, S.; Pacioni, C.; Warren, K.; Butcher, J.; Robertson, I. Wildlife Disease Passive Surveillance: The Potential Role of Wildlife Rehabilitation Centres. In Proceedings of the 6th National Wildlife Rehabilitation Conference, Canberra, Australia, 9–13 August 2008. [Google Scholar]
- Jiménez-Ruiz, S.; Vicente, J.; García-Bocanegra, I.; Cabezón, Ó.; Arnal, M.C.; Balseiro, A.; Ruiz-Fons, F.; Gómez-Guillamón, F.; Lázaro, S.; Escribano, F.; et al. Distribution of Pestivirus Exposure in Wild Ruminants in Spain. Transbound. Emerg. Dis. 2020, 68, 1577–1585. [Google Scholar] [CrossRef]
- Olde Riekerink, R.G.M.; Dominici, A.; Barkema, H.W.; De Smit, A.J. Seroprevalence of Pestivirus in Four Species of Alpine Wild Ungulates in the High Valley of Susa, Italy. Vet. Microbiol. 2005, 108, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Sirera, L.; Cabezón, O.; Dematteis, A.; Rossi, L.; Meneguz, P.G.; Gennero, M.S.; Allepuz, A.; Rosell, R.; Lavín, S.; Marco, I. Survey of Pestivirus Infection in Wild and Domestic Ungulates from South-Western Italian Alps. Eur. J. Wildl. Res. 2012, 58, 425–431. [Google Scholar] [CrossRef]
- Kaba, J.; Strzałkowska, N.; Jóźwik, A.; Krzyzewski, J.; Bagnicka, E. Twelve-Year Cohort Study on the Influence of Caprine Arthritis-Encephalitis Virus Infection on Milk Yield and Composition. J. Dairy Sci. 2012, 95, 1617–1622. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Navalón, B.; Peris, C.; Gómez, E.A.; Peris, B.; Roche, M.L.; Caballero, C.; Goyena, E.; Berriatua, E. Quantitative Estimation of the Impact of Caprine Arthritis Encephalitis Virus Infection on Milk Production by Dairy Goats. Vet. J. 2013, 197, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Huaman, J.L.; Pacioni, C.; Sarker, S.; Doyle, M.; Forsyth, D.M.; Pople, A.; Carvalho, T.G.; Helbig, K.J. Novel Picornavirus Detected in Wild Deer: Identification, Genomic Characterisation, and Prevalence in Australia. Viruses 2021, 13, 2412. [Google Scholar] [CrossRef] [PubMed]
- László, Z.; Pankovics, P.; Reuter, G.; Cságola, A.; Bálint, Á.; Albert, M.; Boros, Á. Multiple Types of Novel Enteric Bopiviruses (Picornaviridae) with the Possibility of Interspecies Transmission Identified from Cloven-Hoofed Domestic Livestock (Ovine, Caprine and Bovine) in Hungary. Viruses 2021, 13, 66. [Google Scholar] [CrossRef]
- Snodgrass, D.R.; Gray, E.W. Detection and Transmission of 30 Nm Virus Particles (Astroviruses) in Faeces of Lambs with Diarrhoea. Arch. Virol. 1977, 55, 287–291. [Google Scholar] [CrossRef]
- Woode, G.N.; Bridger, J.C. Isolation of Small Viruses Resembling Astroviruses and Caliciviruses from Acute Enteritis of Calves. J. Med. Microbiol. 1978, 11, 441–452. [Google Scholar] [CrossRef]
- Campbell, S.J.; Ashley, W.; Gil-Fernandez, M.; Newsome, T.M.; Di Giallonardo, F.; Ortiz-Baez, A.S.; Mahar, J.E.; Towerton, A.L.; Gillings, M.; Holmes, E.C.; et al. Red Fox Viromes in Urban and Rural Landscapes. Virus Evol. 2020, 6, veaa065. [Google Scholar] [CrossRef]
- Wang, L.; Shen, H.; Zheng, Y.; Schumacher, L.; Li, G. Astrovirus in White-Tailed Deer, United States, 2018. Emerg. Infect. Dis. 2020, 26, 374. [Google Scholar] [CrossRef]
- Hoshino, Y.; Zimmer, J.F.; Moise, N.S.; Scott, F.W. Detection of Astroviruses in Feces of a Cat with Diarrhea. Arch. Virol. 1981, 70, 373–376. [Google Scholar] [CrossRef]
- Kurtz, J.B.; Lee, T.W. Astroviruses: Human and Animal. Ciba Found. Symp. 1987, 128, 92–107. [Google Scholar] [CrossRef] [PubMed]
- Toffan, A.; Jonassen, C.M.; De Battisti, C.; Schiavon, E.; Kofstad, T.; Capua, I.; Cattoli, G. Genetic Characterization of a New Astrovirus Detected in Dogs Suffering from Diarrhoea. Vet. Microbiol. 2009, 139, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.E.; Mitchell, D.K. Astrovirus infection in children. Curr. Opin. Infect. Dis. 2003, 16, 247–253. [Google Scholar] [CrossRef] [PubMed]
- . Reuter, G.; Nemes, C.; Boros, Á.; Kapusinszky, B.; Delwart, E.; Pankovics, P. Astrovirus in Wild Boars (Sus scrofa) in Hungary. Arch. Virol. 2012, 157, 1143–1147. [Google Scholar] [CrossRef]
- Pankovics, P.; Boros, Á.; Kiss, T.; Delwart, E.; Reuter, G. Detection of a Mammalian-like Astrovirus in Bird, European Roller (Coracias garrulus). Infect. Genet. Evol. 2015, 34, 114–121. [Google Scholar] [CrossRef]
- Kowalczyk, M.; Jakubczak, A. Mink Astrovirus Diseases—Cause, Symptoms, Diagnosis. Wiadomości Zootech. 2016, 54, 96–101. [Google Scholar]
- Guix, S.; Bosch, A.; Pintó, R.M. Astrovirus Taxonomy. In Astrovirus Research: Essential Ideas, Everyday Impacts, Future Directions; Springer: Berlin/Heidelberg, Germany, 2013; pp. 97–118. ISBN 978-1-4614-4735-1. [Google Scholar] [CrossRef]
- Di Martino, B.; Di Profio, F.; Melegari, I.; Robetto, S.; Di Felice, E.; Orusa, R.; Marsilio, F. Molecular Evidence of Kobuviruses in Free-Ranging Red Foxes (Vulpes vulpes). Arch. Virol. 2014, 159, 1803–1806. [Google Scholar] [CrossRef]
- Reuter, G.; Egyed, L. Bovine Kobuvirus in Europe. Emerg. Infect. Dis. 2009, 15, 822. [Google Scholar] [CrossRef]
- Reuter, G.; Boros, Á.; Pankovics, P.; Egyed, L. Kobuvirus in Domestic Sheep, Hungary. Emerg. Infect. Dis. 2010, 16, 869. [Google Scholar] [CrossRef]
- Reuter, G.; Boros, Á.; Pankovics, P. Kobuviruses—A Comprehensive Review. Rev. Med. Virol. 2011, 21, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G.; Nemes, C.; Boros, Á.; Kapusinszky, B.; Delwart, E.; Pankovics, P. Porcine Kobuvirus in Wild Boars (Sus scrofa). Arch. Virol. 2013, 158, 281–282. [Google Scholar] [CrossRef] [PubMed]
- Melegari, I.; Di Profio, F.; Sarchese, V.; Martella, V.; Marsilio, F.; Di Martino, B. First Molecular Evidence of Kobuviruses in Goats in Italy. Arch. Virol. 2016, 161, 3245–3248. [Google Scholar] [CrossRef]
- Melegari, I.; Sarchese, V.; Di Profio, F.; Robetto, S.; Carella, E.; Bermudez Sanchez, S.; Orusa, R.; Martella, V.; Marsilio, F.; Di Martino, B. First Molecular Identification of Kobuviruses in Wolves (Canis lupus) in Italy. Arch. Virol. 2018, 163, 509–513. [Google Scholar] [CrossRef]
- Di Martino, B.; Di Felice, E.; Ceci, C.; Di Profio, F.; Marsilio, F. Canine Kobuviruses in Diarrhoeic Dogs in Italy. Vet. Microbiol. 2013, 166, 246–249. [Google Scholar] [CrossRef]
- Carmona-Vicente, N.; Buesa, J.; Brown, P.A.; Merga, J.Y.; Darby, A.C.; Stavisky, J.; Sadler, L.; Gaskell, R.M.; Dawson, S.; Radford, A.D. Phylogeny and Prevalence of Kobuviruses in Dogs and Cats in the UK. Vet. Microbiol. 2013, 164, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Olarte-Castillo, X.A.; Heeger, F.; Mazzoni, C.J.; Greenwood, A.D.; Fyumagwa, R.; Moehlman, P.D.; Hofer, H.; East, M.L. Molecular Characterization of Canine Kobuvirus in Wild Carnivores and the Domestic Dog in Africa. Virology 2015, 477, 89–97. [Google Scholar] [CrossRef]
- Woods, L.W. Adenoviral Diseases. In Infectious Diseases of Wild Mammals; Wiley: Hoboken, NJ, USA, 2008; pp. 202–213. [Google Scholar] [CrossRef]
- Åkerstedt, J.; Lillehaug, A.; Larsen, I.L.; Eide, N.E.; Arnemo, J.M.; Handeland, K. Serosurvey for Canine Distemper Virus, Canine Adenovirus, Leptospira Interrogans, and Toxoplasma Gondii in Free-Ranging Canids in Scandinavia and Svalbard. J. Wildl. Dis. 2010, 46, 474–480. [Google Scholar] [CrossRef]
- Thompson, H.; Philbey, A.W.; Lewis, J.C.M.; Stocker, L.R.; Laurenson, M.K. Infectious Canine Hepatitis in Red Foxes (Vulpes vulpes) in the United Kingdom. Vet. Rec. 2010, 166, 111–114. [Google Scholar] [CrossRef]
- Millán, J.; López-Bao, J.V.; Garcıá, E.J.; Oleaga, Á.; Llaneza, L.; Palacios, V.; De La Torre, A.; Rodríguez, A.; Dubovi, E.J.; Esperón, F. Patterns of Exposure of Iberian Wolves (Canis lupus) to Canine Viruses in Human-Dominated Landscapes. Ecohealth 2016, 13, 123–134. [Google Scholar] [CrossRef]
- Knowles, S.; Bodenstein, B.L.; Hamon, T.; Saxton, M.W.; Hall, J.S. Infectious Canine Hepatitis in a Brown Bear (Ursus Arctos Horribilis) from Alaska, USA. J. Wildl. Dis. 2018, 54, 642–645. [Google Scholar] [CrossRef]
- Pyziel, A. Studies on Eimerians (Apicomplexa: Eimeriidae) of Wild Ruminants. Ann. Parasitol. 2012, 58, 105–106. [Google Scholar] [PubMed]
- Duszynski, D.W.; Couch, L.; Upton, S.J. Coccidia (Eimeriidae) of Canidae and Felidae. Available online: http://eimeria.unl.edu/carniv1.html (accessed on 2 January 2023).
- Newman, C.; Macdonald, D.W.; Anwar, M.A. Coccidiosis in the European Badger, Meles Meles in Wytham Woods: Infection and Consequences for Growth and Survival. Parasitology 2001, 123, 133–142. [Google Scholar] [CrossRef]
- Burlinski, P.; Janiszewski, P.; Anna, K.; Gonkowski, S. Parasitofauna in the Gastrointestinal Tract of the Cervids (Cervidae) in Northern Poland. Acta Vet. 2011, 61, 269–282. [Google Scholar] [CrossRef]
- Borgsteede, F.H.M.; Jansen, J.; van Nispen tot Pannerden, H.P.M.; van der Burg, W.P.J.; Noorman, N.; Poutsma, J.; Kotter, J.F. A Study of the Endoparasitic Helminth Fauna of Roe Deer (Capreolus capreolus L.) in the Netherlands. Z. Jagdwiss. 1990, 36, 104–109. [Google Scholar] [CrossRef]
- Bolukbas, C.S.; Gurler, A.T.; Beyhan, Y.E.; Acici, M.; Umur, S. Helminths of Roe Deer (Capreolus capreolus) in the Middle Black Sea Region of Turkey. Parasitol. Int. 2012, 61, 729–730. [Google Scholar] [CrossRef]
- Gibbons, L.; Höglund, J. Dictyocaulus Capreolus n. sp. (Nematoda: Trichostrongyloidea) from Roe Deer, Capreolus Capreolus and Moose, Alces Alces in Sweden. J. Helminthol. 2002, 76, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Jurankova, J.; Jirsova, D.; Pafco, B.; Forejtek, P. The Molecular and Morphometric Identification of Dictyocaulus Capreolus in Clinically Affected Roe Deer (Capreolus capreolus L.). Vet. Med. 2019, 64, 386–391. [Google Scholar] [CrossRef]
- Fiocchi, A.; Gustinelli, A.; Gelmini, L.; Rugna, G.; Renzi, M.; Fontana, M.C.; Poglayen, G. Helminth Parasites of the Red Fox Vulpes vulpes (L., 1758) and the Wolf Canis lupus italicus Altobello, 1921 in Emilia-Romagna, Italy. Ital. J. Zool. 2016, 83, 503–513. [Google Scholar] [CrossRef]
- Magi, M.; Guardone, L.; Prati, M.C.; Mignone, W.; Macchioni, F. Extraintestinal Nematodes of the Red Fox Vulpes Vulpes in North-West Italy. J. Helminthol. 2015, 89, 506–511. [Google Scholar] [CrossRef]
- Veronesi, F.; Morganti, G.; di Cesare, A.; Lepri, E.; Cassini, R.; Zanet, S.; Deni, D.; Chiari, M.; Ferroglio, E. Eucoleus Boehmi Infection in Red Fox (Vulpes vulpes) from Italy. Vet. Parasitol. 2014, 206, 232–239. [Google Scholar] [CrossRef]
- Seguel, M.; Gottdenker, N. The Diversity and Impact of Hookworm Infections in Wildlife. Int. J. Parasitol. Parasites Wildl. 2017, 6, 177–194. [Google Scholar] [CrossRef]
- András, T.; László, S.; Attila, T.A.; Tamás, T.P.; Miklós, H. Data on the Parasitological State of the Eurasian Badger (Meles meles) in Hungary. Magy. Állatorvosok Lapja 2012, 134, 106–110. [Google Scholar]
- Torres, J.; Miquel, J.; Motjé, M. Helminth Parasites of the Eurasian Badger (Meles meles L.) in Spain: A Biogeographic Approach. Parasitol. Res. 2001, 87, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Millán, J.; Sevilla, I.; Gerrikagoitia, X.; García-Pérez, A.L.; Barral, M. Helminth Parasites of the Eurasian Badger (Meles meles L.) in the Basque Country (Spain). Eur. J. Wildl. Res. 2004, 50, 37–40. [Google Scholar] [CrossRef]
- Byrne, R.L.; Fogarty, U.; Mooney, A.; Harris, E.; Good, M.; Marples, N.M.; Holland, C.V. The Helminth Parasite Community of European Badgers (Meles meles) in Ireland. J. Helminthol. 2019, 94, e37. [Google Scholar] [CrossRef]
- Latrofa, M.S.; Lia, R.P.; Giannelli, A.; Colella, V.; Santoro, M.; D’Alessio, N.; Campbell, B.E.; Parisi, A.; Dantas-Torres, F.; Mutafchiev, Y.; et al. Crenosoma Vulpis in Wild and Domestic Carnivores from Italy: A Morphological and Molecular Study. Parasitol. Res. 2015, 114, 3611–3617. [Google Scholar] [CrossRef]
- Ko, P.P.; Suzuki, K.; Canales-Ramos, M.; Aung, M.P.P.T.H.H.; Htike, W.W.; Yoshida, A.; Montes, M.; Morishita, K.; Gotuzzo, E.; Maruyama, H.; et al. Phylogenetic Relationships of Strongyloides Species in Carnivore Hosts. Parasitol. Int. 2020, 78, 102151. [Google Scholar] [CrossRef]
- Ionică, A.M.; Deak, G.; Boncea, R.; Gherman, C.M.; Mihalca, A.D. The European Badger as a New Host for Dirofilaria Immitis and an Update on the Distribution of the Heartworm in Wild Carnivores from Romania. Pathogens 2022, 11, 420. [Google Scholar] [CrossRef] [PubMed]
- Nagy, E.; Benedek, I.; Zsolnai, A.; Halász, T.; Csivincsik, Á.; Ács, V.; Nagy, G.; Tari, T. Habitat Characteristics as Potential Drivers of the Angiostrongylus Daskalovi Infection in European Badger (Meles meles) Populations. Pathogens 2021, 10, 715. [Google Scholar] [CrossRef]
- Gherman, C.M.; Deak, G.; Matei, I.A.; IonicǍ, A.M.; D’Amico, G.; Taulescu, M.; Barbu-Tudoran, L.; Sarmaşi, A.; Mihalca, A.D.; Cozma, V. A Rare Cardiopulmonary Parasite of the European Badger, Meles meles: First Description of the Larvae, Ultrastructure, Pathological Changes and Molecular Identification of Angiostrongylus Daskalovi Janchev & Genov 1988. Parasites Vectors 2016, 9, 1–10. [Google Scholar] [CrossRef]
- Almería, S. Neospora Caninum and Wildlife. ISRN Parasitol. 2013, 2013. [Google Scholar] [CrossRef]
- Ryan, U.; Cacciò, S.M. Zoonotic Potential of Giardia. Int. J. Parasitol. 2013, 43, 943–956. [Google Scholar] [CrossRef]
- Cai, W.; Ryan, U.; Xiao, L.; Feng, Y. Zoonotic Giardiasis: An Update. Parasitol. Res. 2021, 120, 4199–4218. [Google Scholar] [CrossRef] [PubMed]
- Regione Toscana. Risultati di Applicazione Della Legge Regionale n. 10 del 9 Febbraio 2016 “Legge Obiettivo per la Gestione Degli Ungulati in Toscana”. 2018. Available online: https://www.consiglio.regione.toscana.it/upload/COCCOINA/documenti/nota%20informativa%2040_pub.pdf (accessed on 10 September 2022).
- Piano Locale di Sviluppo Rurale (PLSR) Della Provincia di Pisa. 2008. Available online: https://www.regione.toscana.it/documents/10180/11243391/pisa/9b71a602-921f-44ab-8f1c-5454527ffed4 (accessed on 10 September 2022).
- Statistiche Demografiche. Provincia Di PISA, Densità Abitativa, Popolazione, Età Media, Famiglie, Stranieri. Available online: https://ugeo.urbistat.com/AdminStat/it/it/demografia/dati-sintesi/pisa/50/3 (accessed on 5 December 2022).
- Pacini, M.I.; Bonelli, F.; Briganti, A.; Citi, S.; Papini, R.A.; Sgorbini, M. Wildlife Ungulate Rescue and Emergency Services in the Pisa Area (Tuscany, Italy): Evaluation of a 9-Years Period (2010–2018). Front. Vet. Sci. 2020, 7, 626. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, B.; Di Profio, F.; Melegari, I.; Di Felice, E.; Robetto, S.; Guidetti, C.; Orusa, R.; Martella, V.; Marsilio, F. Molecular Detection of Kobuviruses in European Roe Deer (Capreolus capreolus) in Italy. Arch. Virol. 2015, 160, 2083–2086. [Google Scholar] [CrossRef]
- Conceição-Neto, N.; Godinho, R.; Álvares, F.; Yinda, C.K.; Deboutte, W.; Zeller, M.; Laenen, L.; Heylen, E.; Roque, S.; Petrucci-Fonseca, F.; et al. Viral Gut Metagenomics of Sympatric Wild and Domestic Canids, and Monitoring of Viruses: Insights from an Endangered Wolf Population. Ecol. Evol. 2017, 7, 4135–4146. [Google Scholar] [CrossRef] [PubMed]
- Ricci, S.; Bartolini, S.; Morandi, F.; Cuteri, V.; Preziuso, S. Genotyping of Pestivirus A (Bovine Viral Diarrhea Virus 1) Detected in Faeces and in Other Specimens of Domestic and Wild Ruminants at the Wildlife-Livestock Interface. Vet. Microbiol. 2019, 235, 180–187. [Google Scholar] [CrossRef]
- van den Brand, J.M.A.; van Leeuwen, M.; Schapendonk, C.M.; Simon, J.H.; Haagmans, B.L.; Osterhaus, A.D.M.E.; Smits, S.L. Metagenomic Analysis of the Viral Flora of Pine Marten and European Badger Feces. J. Virol. 2012, 86, 2360–2365. [Google Scholar] [CrossRef]
- Walker, D.; Gregory, W.F.; Turnbull, D.; Rocchi, M.; Meredith, A.L.; Philbey, A.W.; Sharp, C.P. Novel Adenoviruses Detected in British Mustelids, Including a Unique Aviadenovirus in the Tissues of Pine Martens (Martes martes). J. Med. Microbiol. 2017, 66, 1177. [Google Scholar] [CrossRef]
- Di Sabatino, D.; Di Francesco, G.; Zaccaria, G.; Malatesta, D.; Brugnola, L.; Marcacci, M.; Portanti, O.; De Massis, F.; Savini, G.; Teodori, L.; et al. Lethal Distemper in Badgers (Meles meles) Following Epidemic in Dogs and Wolves. Infect. Genet. Evol. 2016, 46, 130–137. [Google Scholar] [CrossRef]
- Bodewes, R.; Ruiz-Gonzalez, A.; Schapendonk, C.M.E.; Van Den Brand, J.M.A.; Osterhaus, A.D.M.E.; Smits, S.L. Viral Metagenomic Analysis of Feces of Wild Small Carnivores. Virol. J. 2014, 11, 89. [Google Scholar] [CrossRef]
- Balboni, A.; Verin, R.; Morandi, F.; Poli, A.; Prosperi, S.; Battilani, M. Molecular Epidemiology of Canine Adenovirus Type 1 and Type 2 in Free-Ranging Red Foxes (Vulpes vulpes) in Italy. Vet. Microbiol. 2013, 162, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Pratelli, A.; Cirone, F.; Zizzo, N.; Decaro, N.; Tinelli, A.; Foti, M.; Buonavoglia, C. Detection and Genetic Characterization of Canine Distemper Virus (CDV) from Free-Ranging Red Foxes in Italy. Mol. Cell. Probes 2002, 16, 77–83. [Google Scholar] [CrossRef]
- Ndiana, L.A.; Lanave, G.; Desario, C.; Berjaoui, S.; Alfano, F.; Puglia, I.; Fusco, G.; Colaianni, M.L.; Vincifori, G.; Camarda, A.; et al. Circulation of Diverse Protoparvoviruses in Wild Carnivores, Italy. Transbound. Emerg. Dis. 2021, 68, 2489–2502. [Google Scholar] [CrossRef]
- Hu, R.L.; Huang, G.; Qiu, W.; Zhong, Z.H.; Xia, X.Z.; Yin, Z. Detection and Differentiation of CAV-1 and CAV-2 by Polymerase Chain Reaction. Vet. Res. Commun. 2001, 25, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.K.W.; Leung, C.Y.H.; Gilbert, M.; Joyner, P.H.; Ng, E.M.; Tse, T.M.; Guan, Y.; Peiris, J.S.M.; Poon, L.L.M. Avian Coronavirus in Wild Aquatic Birds. J. Virol. 2011, 85, 12815–12820. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.K.W.; Poon, L.L.M.; Guan, Y.; Peiris, J.S.M. Novel Astroviruses in Insectivorous Bats. J. Virol. 2008, 82, 9107. [Google Scholar] [CrossRef]
- Schatzberg, S.J.; Haley, N.J.; Barr, S.C.; Parrish, C.; Steingold, S.; Summers, B.A.; Delahunta, A.; Kornegay, J.N.; Sharp, N.J.H. Polymerase Chain Reaction (PCR) Amplification of Parvoviral DNA from the Brains of Dogs and Cats with Cerebellar Hypoplasia. J. Vet. Intern. Med. 2003, 17, 538–544. [Google Scholar] [CrossRef]
- Kapoor, A.; Mehta, N.; Dubovi, E.J.; Simmonds, P.; Govindasamy, L.; Medina, J.L.; Street, C.; Shields, S.; Ian Lipkin, W. Characterization of Novel Canine Bocaviruses and Their Association with Respiratory Disease. J. Gen. Virol. 2012, 93, 341. [Google Scholar] [CrossRef]
- Lau, S.K.P.; Woo, P.C.Y.; Yeung, H.C.; Teng, J.L.L.; Wu, Y.; Bai, R.; Fan, R.Y.Y.; Chan, K.H.; Yuen, K.Y. Identification and Characterization of Bocaviruses in Cats and Dogs Reveals a Novel Feline Bocavirus and a Novel Genetic Group of Canine Bocavirus. J. Gen. Virol. 2012, 93, 1573–1582. [Google Scholar] [CrossRef]
- Li, L.; Pesavento, P.A.; Leutenegger, C.M.; Estrada, M.; Coffey, L.L.; Naccache, S.N.; Samayoa, E.; Chiu, C.; Qiu, J.; Wang, C.; et al. A Novel Bocavirus in Canine Liver. Virol. J. 2013, 10, 54. [Google Scholar] [CrossRef]
- Segalés, J.; Martínez-Guinó, L.; Cortey, M.; Navarro, N.; Huerta, E.; Sibila, M.; Pujols, J.; Kekarainen, T. Retrospective Study on Swine Torque Teno Virus Genogroups 1 and 2 Infection from 1985 to 2005 in Spain. Vet. Microbiol. 2009, 134, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Vilček, S.; Herring, A.J.; Herring, J.A.; Nettleton, P.F.; Lowings, J.P.; Paton, D.J. Pestiviruses Isolated from Pigs, Cattle and Sheep Can Be Allocated into at Least Three Genogroups Using Polymerase Chain Reaction and Restriction Endonuclease Analysis. Arch. Virol. 1994, 136, 309–323. [Google Scholar] [CrossRef]
- Reuter, G.; Boldizsár, Á.; Pankovics, P. Complete Nucleotide and Amino Acid Sequences and Genetic Organization of Porcine Kobuvirus, a Member of a New Species in the Genus Kobuvirus, Family Picornaviridae. Arch. Virol. 2009, 154, 101–108. [Google Scholar] [CrossRef]
- Sheehan, M.; Markey, B.; Cassidy, J.; Ball, H.; Duane, M.; Doherty, M.L. New Transtracheal Bronchoalveolar Lavage Technique for the Diagnosis of Respiratory Disease in Sheep. Vet. Rec. 2005, 157, 309–313. [Google Scholar] [CrossRef]
- Sulaiman, I.M.; Fayer, R.; Bern, C.; Gilman, R.H.; Trout, J.M.; Schantz, P.M.; Das, P.; Lal, A.A.; Xiao, L. Triosephosphate Isomerase Gene Characterization and Potential Zoonotic Transmission of Giardia Duodenalis. Emerg. Infect. Dis. 2003, 9, 1444–1452. [Google Scholar] [CrossRef] [PubMed]
- Cacciò, S.M.; De Giacomo, M.; Pozio, E. Sequence Analysis of the β-Giardin Gene and Development of a Polymerase Chain Reaction-Restriction Fragment Length Polymorphism Assay to Genotype Giardia Duodenalis Cysts from Human Faecal Samples. Int. J. Parasitol. 2002, 32, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.; Xiao, L.; Sulaiman, I.; Lal, A.A.; Matos, O.; Antunes, F. Subgenotype Analysis of Cryptosporidium Isolates from Humans, Cattle, and Zoo Ruminants in Portugal. J. Clin. Microbiol. 2003, 41, 2744–2747. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A Greedy Algorithm for Aligning DNA Sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Morgulis, A.; Coulouris, G.; Raytselis, Y.; Madden, T.L.; Agarwala, R.; Schäffer, A.A. Database Indexing for Production MegaBLAST Searches. Bioinformatics 2008, 24, 1757–1764. [Google Scholar] [CrossRef]
- Nielsen, S.S.; Roensholt, L.; Bitsch, V. Bovine Virus Diarrhea Virus in Free-Living Deer from Denmark. J. Wildl. Dis. 2000, 36, 584–587. [Google Scholar] [CrossRef] [PubMed]
- Lillehaug, A.; Vikøren, T.; Larsen, I.-L.; Kerstedt, J.A.; Tharaldsen, J.; Handeland, K. Antibodies to ruminant alpha-herpesviruses and pestiviruses in norwegian cervids. J. Wildl. Dis. 2003, 39, 779–786. [Google Scholar] [CrossRef]
- Böhm, M.; White, P.C.L.; Chambers, J.; Smith, L.; Hutchings, M.R. Wild Deer as a Source of Infection for Livestock and Humans in the UK. Vet. J. 2007, 174, 260–276. [Google Scholar] [CrossRef]
- Tavernier, P.; Sys, S.U.; De Clercq, K.; De Leeuw, I.; Caij, A.B.; De Baere, M.; De Regge, N.; Fretin, D.; Roupie, V.; Govaerts, M.; et al. Serologic Screening for 13 Infectious Agents in Roe Deer (Capreolus capreolus) in Flanders. Infect. Ecol. Epidemiol. 2015, 5, 29862. [Google Scholar] [CrossRef]
- Fischer, S.; Weiland, E.; Frolich, K. Characterization of a Bovine Viral Virus from. J. Wildl. Dis. 1998, 34, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Franklin, S.P.; Troyer, J.L.; Terwee, J.A.; Lyren, L.M.; Boyce, W.M.; Riley, S.P.D.; Roelke, M.E.; Crooks, K.R.; VandeWoude, S. Frequent Transmission of Immunodeficiency Viruses among Bobcats and Pumas. J. Virol. 2007, 81, 10961–10969. [Google Scholar] [CrossRef] [PubMed]
- da Cruz, J.C.M.; Singh, D.K.; Lamara, A.; Chebloune, Y. Small Ruminant Lentiviruses (SRLVs) Break the Species Barrier to Acquire New Host Range. Viruses 2013, 5, 1867–1884. [Google Scholar] [CrossRef] [PubMed]
- Patton, K.M.; Bildfell, R.J.; Anderson, M.L.; Cebra, C.K.; Valentine, B.A. Fatal Caprine Arthritis Encephalitis Virus-like Infection in 4 Rocky Mountain Goats (Oreamnos americanus). J. Vet. Diagn. Investig. 2012, 24, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Olech, M.; Osiński, Z.; Kuźmak, J. Seroprevalence of Small Ruminant Lentivirus (SRLV) Infection in Wild Cervids in Poland. Prev. Vet. Med. 2020, 176, 104905. [Google Scholar] [CrossRef]
- Kuhar, U.; Vengušt, D.Ž.; Vengušt, G. Serological Survey of Small Ruminant Lentivirus Infections in Free-Ranging Mouflon and Chamois in Slovenia. Animals 2022, 12, 1032. [Google Scholar] [CrossRef] [PubMed]
- Palombieri, A.; Fruci, P.; Di Profio, F.; Sarchese, V.; Robetto, S.; Martella, V.; Di Martino, B. Detection and Characterization of Bopiviruses in Domestic and Wild Ruminants. Transbound. Emerg. Dis. 2022, 69, 3972–3978. [Google Scholar] [CrossRef]
- Smits, S.L.; Van Leeuwen, M.; Kuiken, T.; Hammer, A.S.; Simon, J.H.; Osterhaus, A.D.M.E. Identification and Characterization of Deer Astroviruses. J. Gen. Virol. 2010, 91, 2719–2722. [Google Scholar] [CrossRef]
- Jamnikar-Ciglenecki, U.; Civnik, V.; Kirbis, A.; Kuhar, U. A Molecular Survey, Whole Genome Sequencing and Phylogenetic Analysis of Astroviruses from Roe Deer. BMC Vet. Res. 2020, 16, 68. [Google Scholar] [CrossRef] [PubMed]
- Moser, L.A.; Schultz-Cherry, S. Pathogenesis of Astrovirus Infection. Viral Immunol. 2005, 18, 4–10. [Google Scholar] [CrossRef]
- Blomström, A.L.; Widén, F.; Hammer, A.S.; Belák, S.; Berg, M. Detection of a Novel Astrovirus in Brain Tissue of Mink Suffering from Shaking Mink Syndrome by Use of Viral Metagenomics. J. Clin. Microbiol. 2010, 48, 4392–4396. [Google Scholar] [CrossRef]
- Li, L.; Diab, S.; McGraw, S.; Barr, B.; Traslavina, R.; Higgins, R.; Talbot, T.; Blanchard, P.; Rimoldi, G.; Fahsbender, E.; et al. Divergent Astrovirus Associated with Neurologic Disease in Cattle. Emerg. Infect. Dis. 2013, 19, 1385–1392. [Google Scholar] [CrossRef]
- Naccache, S.N.; Peggs, K.S.; Mattes, F.M.; Phadke, R.; Garson, J.A.; Grant, P.; Samayoa, E.; Federman, S.; Miller, S.; Lunn, M.P.; et al. Diagnosis of Neuroinvasive Astrovirus Infection in an Immunocompromised Adult with Encephalitis by Unbiased Next-Generation Sequencing. Clin. Infect. Dis. 2015, 60, 919–923. [Google Scholar] [CrossRef]
- Pfaff, F.; Schlottau, K.; Scholes, S.; Courtenay, A.; Hoffmann, B.; Höper, D.; Beer, M. A Novel Astrovirus Associated with Encephalitis and Ganglionitis in Domestic Sheep. Transbound. Emerg. Dis. 2017, 64, 677–682. [Google Scholar] [CrossRef]
- Di Bartolo, I.; Angeloni, G.; Tofani, S.; Monini, M.; Ruggeri, F.M. Infection of Farmed Pigs with Porcine Kobuviruses in Italy. Arch. Virol. 2015, 160, 1533–1536. [Google Scholar] [CrossRef] [PubMed]
- von Messling, V.; Springfeld, C.; Devaux, P.; Cattaneo, R. A Ferret Model of Canine Distemper Virus Virulence and Immunosuppression. J. Virol. 2003, 77, 12579–12591. [Google Scholar] [CrossRef]
- Miyabe, F.M.; Ribeiro, J.; Alfieri, A.F.; Alfieri, A.A. Detection of Canine Kobuvirus RNA in Diarrheic Fecal Samples of Dogs with Parvoviruses. Braz. J. Microbiol. 2019, 50, 871–874. [Google Scholar] [CrossRef]
- Kaiser, F.K.; van Dyck, L.; Jo, W.K.; Schreiner, T.; Pfankuche, V.M.; Wohlsein, P.; Baumann, I.; Peters, M.; Baumgärtner, W.; Osterhaus, A.D.M.E.; et al. Detection of Systemic Canine Kobuvirus Infection in Peripheral Tissues and the Central Nervous System of a Fox Infected with Canine Distemper Virus. Microorganisms 2021, 9, 2521. [Google Scholar] [CrossRef] [PubMed]
- Oem, J.K.; Choi, J.W.; Lee, M.H.; Lee, K.K.; Choi, K.S. Canine Kobuvirus Infections in Korean Dogs. Arch. Virol. 2014, 159, 2751–2755. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Cui, Y.; Jiang, S.; Liu, H.; Wang, J.; Li, Y. Duplex SYBR Green I-Based Real-Time PCR Assay for the Rapid Detection of Canine Kobuvirus and Canine Astrovirus. J. Virol. Methods 2021, 290, 114066. [Google Scholar] [CrossRef] [PubMed]
- Smits, S.L.; Raj, V.S.; Oduber, M.D.; Schapendonk, C.M.E.; Bodewes, R.; Provacia, L.; Stittelaar, K.J.; Osterhaus, A.D.M.E.; Haagmans, B.L. Metagenomic Analysis of the Ferret Fecal Viral Flora. PLoS ONE 2013, 8, e71595. [Google Scholar] [CrossRef]
- Phan, T.G.; Kapusinszky, B.; Wang, C.; Rose, R.K.; Lipton, H.L.; Delwart, E.L. The Fecal Viral Flora of Wild Rodents. PLOS Pathog. 2011, 7, e1002218. [Google Scholar] [CrossRef] [PubMed]
- Dastjerdi, A.; Benfield, C.; Everest, D.; Stidworthy, M.F.; Zell, R. Novel Enteric Viruses in Fatal Enteritis of Grey Squirrels. J. Gen. Virol. 2020, 101, 746–750. [Google Scholar] [CrossRef]
- Zhang, M.; You, F.; Wu, F.; He, H.; Li, Q.; Chen, Q. Epidemiology and Genetic Characteristics of Murine Kobuvirus from Faecal Samples of Rattus Losea, Rattus Tanezumi and Rattus Norvegicus in Southern China. J. Gen. Virol. 2021, 102, 1646. [Google Scholar] [CrossRef]
- Green, R.G.; Ziegler, N.R.; Green, B.B.; Dewey, E.T. Epizootic Fox Encephalitis. I. General Description. Am. J. Epidemiol. 1930, 12, 109–129. [Google Scholar] [CrossRef]
- Walker, D.; Abbondati, E.; Cox, A.L.; Mitchell, G.B.B.; Pizzi, R.; Sharp, C.P.; Philbey, A.W. Infectious Canine Hepatitis in Red Foxes (Vulpes vulpes) in Wildlife Rescue Centres in the UK. Vet. Rec. 2016, 178, 421. [Google Scholar] [CrossRef]
- Verin, R.; Forzan, M.; Schulze, C.; Rocchigiani, G.; Balboni, A.; Poli, A.; Mazzei, M. Multicentric Molecular and Pathologic Study On Canine Adenovirus Type 1 in Red Foxes (Vulpes vulpes) in Three European Countries. J. Wildl. Dis. 2019, 55, 935–939. [Google Scholar] [CrossRef]
- Robinson, A.J.; Crerar, S.K.; Sharma, W.; Müller, W.J.; Bradley, M.P.; Jackson, A.; Anderson, N.; Browning, G.; Carmel, B.; Condron, R.; et al. Prevalence of Serum Antibodies to Canine Adenovirus and Canine Herpesvirus in the European Red Fox (Vulpes vulpes) in Australia. Aust. Vet. J. 2005, 83, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.; Fee, S.A.; Hartley, G.; Learmount, J.; O’Hagan, M.J.H.; Meredith, A.L.; Bronsvoort, B.M.D.C.; Porphyre, T.; Sharp, C.P.; Philbey, A.W. Serological and Molecular Epidemiology of Canine Adenovirus Type 1 in Red Foxes (Vulpes vulpes) in the United Kingdom. Sci. Rep. 2016, 6, 36051. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Campolo, M.; Elia, G.; Buonavoglia, D.; Colaianni, M.L.; Lorusso, A.; Mari, V.; Buonavoglia, C. Infectious Canine Hepatitis: An “Old” Disease Reemerging in Italy. Res. Vet. Sci. 2007, 83, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Park, N.Y.; Lee, M.C.; Kurkure, N.V.; Cho, H.S. Canine Adenovirus Type 1 Infection of a Eurasian River Otter (Lutra lutra). Vet. Pathol. 2007, 44, 536–539. [Google Scholar] [CrossRef] [PubMed]
- Kozak, R.A.; Ackford, J.G.; Slaine, P.; Li, A.; Carman, S.; Campbell, D.; Welch, M.K.; Kropinski, A.M.; Nagy, É. Characterization of a Novel Adenovirus Isolated from a Skunk. Virology 2015, 485, 16–24. [Google Scholar] [CrossRef]
- ADM Technical Bulletin—Internal Parasites of Deer; ADM Monograms; ADM: Chicago, IL, USA, 2005.
- Vervaeke, M.; Dorny, P.; De Bruyn, L.; Vercammen, F.; Jordaens, K.; Van Den Berge, K.; Verhagen, R. A Survey of Intestinal Helminths of Red Foxes (Vulpes vulpes) in Northern Belgium. Acta Parasitol. 2005, 50, 221–227. [Google Scholar]
- Eira, C.; Vingada, J.; Torres, J.; Miquel, J. The Helminth Community of the Red Fox, Vulpes vulpes, In Dunas de Mira (Portugal) and Its Effect on Host Condition. Wildl. Biol. Pract. 2006, 2, 26–36. [Google Scholar] [CrossRef]
- Hernández, F.A.; Manqui, J.; Mejías, C.; Acosta-Jamett, G. Domestic Dogs and Wild Foxes Interactions in a Wildlife-Domestic Interface of North-Central Chile: Implications for Multi-Host Pathogen Transmission. Front. Vet. Sci. 2021, 8, 631788. [Google Scholar] [CrossRef]
- Karamon, J.; Dabrowska, J.; Kochanowski, M.; Samorek-Pieróg, M.; Sroka, J.; Różycki, M.; Bilska-Zajac, E.; Zdybel, J.; Cencek, T. Prevalence of Intestinal Helminths of Red Foxes (Vulpes vulpes) in Central Europe (Poland): A Significant Zoonotic Threat. Parasites Vectors 2018, 11, 436. [Google Scholar] [CrossRef] [PubMed]
- Traversa, D.; Torbidone, A.; Malatesta, D.; Guglielmini, C. Occurrence of Fatal Canine Angiostrongylus Vasorum Infection in Italy. Vet. Parasitol. 2008, 152, 162–166. [Google Scholar] [CrossRef]
- Morchón, R.; Carretón, E.; González-Miguel, J.; Mellado-Hernández, I. Heartworm Disease (Dirofilaria immitis) and Their Vectors in Europe—New Distribution Trends. Front. Physiol. 2012, 3, 196. [Google Scholar] [CrossRef] [PubMed]
- Fuehrer, H.P. An Overview of the Host Spectrum and Distribution of Calodium Hepaticum (Syn. Capillaria Hepatica): Part 1—Muroidea. Parasitol. Res. 2014, 113, 619–640. [Google Scholar] [CrossRef]
- Khan, W.; Nisa, N.N.; Pervez, S.; Ahmed, S.; Ahmed, M.S.; Alfarraj, S.; Ali, A.; Tahreem, S. Occurrence of Hymenolepis diminuta: A Potential Helminth of Zoonotic Importance in Murid Rodents. Braz. J. Biol. 2022, 82, e242089. [Google Scholar] [CrossRef] [PubMed]
- Eleni, C.; Mariacher, A.; Grifoni, G.; Cardini, E.; Tonon, S.; Lombardo, A.; Barone, A.; Fichi, G. Pathology of Urinary Bladder in Pearsonema spp. Infected Wildlife from Central Italy. Pathogens 2021, 10, 474. [Google Scholar] [CrossRef]
- Rosypal, A.C.; Alexander, A.; Byrd, D.; Weaver, M.; Stewart, R.; Gerhold, R.; Houston, A.; Van Why, K.; Dubey, J.P. Survey of Antibodies to Leishmania spp. in Wild Canids from Pennsylvania and Tennessee. J. Zoo Wildl. Med. 2013, 44, 1131–1133. [Google Scholar] [CrossRef]
- Ferreira, S.C.M.; Torelli, F.; Klein, S.; Fyumagwa, R.; Karesh, W.B.; Hofer, H.; Seeber, F.; East, M.L. Evidence of High Exposure to Toxoplasma Gondii in Free-Ranging and Captive African Carnivores. Int. J. Parasitol. Parasites Wildl. 2019, 8, 111–117. [Google Scholar] [CrossRef]
- De La Torre, J.R.; Bautista-Piña, C.; Alfonso Ortega, S.J.; Cantu-Covarruvias, A.; Genoveva Alvarez-Ojeda, M.; Romero-Salas, D.; Henke, S.E.; Hilton, C.D.; Hewitt, D.G.; De Young, R.W.; et al. Neospora Caninum in Axis Deer (Axis Axis) and Fallow Deer (Dama Dama) in Northern Mexico. J. Wildl. Dis. 2017, 53, 186–187. [Google Scholar] [CrossRef]
- Rocchigiani, G.; Nardoni, S.; D’Ascenzi, C.; Nicoloso, S.; Picciolli, F.; Amerigo Papini, R.; Mancianti, F. Seroprevalence of Toxoplasma Gondii and Neospora Caninum in Red Deer from Central Italy. Ann. Agric. Environ. Med. 2016, 23, 699–701. [Google Scholar] [CrossRef] [PubMed]
- Bartley, P.M.; Wright, S.E.; Zimmer, I.A.; Roy, S.; Kitchener, A.C.; Meredith, A.; Innes, E.A.; Katzer, F. Detection of Neospora Caninum in Wild Carnivorans in Great Britain. Vet. Parasitol. 2013, 192, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Almería, S.; Ferrer, D.; Pabón, M.; Castellà, J.; Mañas, S. Red Foxes (Vulpes vulpes) Are a Natural Intermediate Host of Neospora Caninum. Vet. Parasitol. 2002, 107, 287–294. [Google Scholar] [CrossRef]
- Marco, I.; Ferroglio, E.; López-Olvera, J.R.; Montané, J.; Lavín, S. High Seroprevalence of Neospora Caninum in the Red Fox (Vulpes vulpes) in the Pyrenees (NE Spain). Vet. Parasitol. 2008, 152, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Hamnes, I.S.; Enemark, H.L.; Henriksen, K.; Madslien, K.; Er, C. The Surveillance Programme for Angiostrongylus Vasorum in Red Foxes (Vulpes vulpes) in Norway 2016; Norwegian Veterinary Institute: Oslo, Norway, 2020.
- Deak, G.; Gherman, C.M.; Ionică, A.M.; Daskalaki, A.A.; Matei, I.A.; D’Amico, G.; Domşa, C.; Pantchev, N.; Mihalca, A.D.; Cozma, V. Use of a Commercial Serologic Test for Angiostrongylus Vasorum for the Detection of A. chabaudi in Wildcats and A. daskalovi in Badgers. Vet. Parasitol. 2017, 233, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Gomes-de-Sá, S.; Santos-Silva, S.; Moreira, A.d.S.; Barradas, P.F.; Amorim, I.; Cardoso, L.; Mesquita, J.R. Dirofilaria Immitis Antigenemia and Microfilaremia in Iberian Wolves and Red Foxes from Portugal. Parasites Vectors 2022, 15, 119. [Google Scholar] [CrossRef]
- Sobotyk, C.; Nguyen, N.; Negrón, V.; Varner, A.; Saleh, M.N.; Hilton, C.; Tomeček, J.M.; Esteve-Gasent, M.D.; Verocai, G.G. Detection of Dirofilaria Immitis via Integrated Serological and Molecular Analyses in Coyotes from Texas, United States. Int. J. Parasitol. Parasites Wildl. 2022, 18, 20–24. [Google Scholar] [CrossRef]
- Roemer, G.W.; Coonan, T.J.; Garcelon, D.K.; Starbird, C.H.; McCall, J.W. Spatial and Temporal Variation in the Seroprevalence of Canine Heartworm Antigen in the Island Fox. J. Wildl. Dis. 2000, 36, 723–728. [Google Scholar] [CrossRef]
- van der Giessen, J.W.B.; de Vries, A.; Roos, M.; Wielinga, P.; Kortbeek, L.M.; Mank, T.G. Genotyping of Giardia in Dutch Patients and Animals: A Phylogenetic Analysis of Human and Animal Isolates. Int. J. Parasitol. 2006, 36, 849–858. [Google Scholar] [CrossRef]
- Solarczyk, P.; Majewska, A.C.; Moskwa, B.; Cabaj, W.; Dabert, M.; Nowosad, P. Multilocus Genotyping of Giardia Duodenalis Isolates from Red Deer (Cervus elaphus) and Roe Deer (Capreolus capreolus) from Poland. Folia Parasitol. 2012, 59, 237–240. [Google Scholar] [CrossRef]
- Stojecki, K.; Sroka, J.; Cacció, S.M.; Cencek, T.; Dutkiewicz, J.; Kusyk, P. Prevalence and Molecular Typing of Giardia Duodenalis in Wildlife from Eastern Poland. Folia Parasitol. 2015, 62, 42. [Google Scholar] [CrossRef]
- García-Presedo, I.; Pedraza-Díaz, S.; González-Warleta, M.; Mezo, M.; Gómez-Bautista, M.; Ortega-Mora, L.M.; Castro-Hermida, J.A. The First Report of Cryptosporidium bovis, C. ryanae and Giardia Duodenalis Sub-Assemblage A-II in Roe Deer (Capreolus capreolus) in Spain. Vet. Parasitol. 2013, 197, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Procesi, I.G.; Di Filippo, M.M.; De Liberato, C.; Lombardo, A.; Brocherel, G.; Perrucci, S.; Di Cave, D.; Berrilli, F. Giardia Duodenalis in Wildlife: Exploring Genotype Diversity in Italy and across Europe. Pathogens 2022, 11, 105. [Google Scholar] [CrossRef] [PubMed]
- Coppola, F.; Maestrini, M.; Berrilli, F.; Guadano Procesi, I.; Felicioli, A.; Perrucci, S. First Report of Giardia Duodenalis Infection in the Crested Porcupine (Hystrix cristata L., 1758). Int. J. Parasitol. Parasites Wildl. 2020, 11, 108–113. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Surface (kmq) | Population | Population Density | Forest/Total Surface Ratio | |
|---|---|---|---|---|
| ZONE A | 824.4 | 328,873 | 398 | 0.3 |
| ZONE B | 588.3 | 63,313 | 107 | 0.2 |
| ZONE C | 939.6 | 25,763 | 27 | 0.5 |
| Species | Sex | Total | |
|---|---|---|---|
| Males | Females | ||
| Roe deer | 17/23 (74%) | 6/23 (26%) | 23 |
| Fallow deer | 3/4 (75%) | 1/4 (25%) | 4 |
| Foxes | 9/12 (75%) | 3/12 (25%) | 12 |
| Badgers | 4/6 (67%) | 2/6 (33%) | 6 |
| Pine martens | - | 1/1 (100%) | 1 |
| Porcupines | 2/4 (50%) | 2/4 (50%) | 4 |
| Total | 35/50 (70%) | 15/50 (30%) | 50 |
| Species | Zone | Total | ||
|---|---|---|---|---|
| Zone A | Zone B | Zone C | ||
| Roe deer | 9/23 (39%) | 11/23 (48%) | 3/23 (13%) | 23 |
| Fallow deer | 4/4 (100%) | - | - | 4 |
| Foxes | 9/12 (75%) | 3/12 (25%) | - | 12 |
| Badgers | 5/6 (83%) | 1/6 (17%) | - | 6 |
| Pine martens | - | 1/1 (100%) | - | 1 |
| Porcupines | 3/4 (75%) | - | 1/4 (25%) | 4 |
| Total | 30/50 (60%) | 16/50 (32%) | 4/50 (8%) | 50 |
| CAdV 1 | KoV | AstV | BoPV | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Roe deer | -- | 1 | 3 | 0 | ||||||||||
| A | B | C | A | B | C | |||||||||
| 1/9 | 0/11 | 0/3 | 1/9 | 1/11 | 1/3 | |||||||||
| Fallow deer | -- | 0 | 0 | 1 | ||||||||||
| A | B | C | ||||||||||||
| 1/4 | 0/0 | 0/0 | ||||||||||||
| Foxes | 3 | 2 | 5 | -- | ||||||||||
| A | B | C | A | B | C | A | B | C | ||||||
| 1/9 | 2/3 | 0/0 | 2/9 | 0/3 | 0/0 | 3/9 | 2/3 | 0/0 | ||||||
| Badger | 2 | 1 | 0 | -- | ||||||||||
| A | B | C | A | B | C | |||||||||
| 1/5 | 1/1 | 0/0 | 1/5 | 0/1 | 0/0 | |||||||||
| Pine Marten | 0 | 0 | 0 | -- | ||||||||||
| Porcupine | -- | 2 | 0 | -- | ||||||||||
| A | B | C | ||||||||||||
| 1/3 | 0/0 | 0/1 | ||||||||||||
| Total | 5 | 6 | 8 | 1 | ||||||||||
| A | B | C | A | B | C | A | B | C | A | B | C | |||
| 2/14 | 3/5 | 0/0 | 6/30 | 0/16 | 0/4 | 4/30 | 3/16 | 1/4 | 1/13 | 0/11 | 0/3 | |||
| Host Species | Faecal Flotation | Baermann Technique | Commercial Assays | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Giardia | Cryptosporidium | Toxoplasma | Neospora | Leishmania | Dirofilaria | Angiostrongylus | |||
| Roe deer (n = 23) | * GIS (10) Eimeria (5) Capillaria (2) | Dictyocaulus (13) | 1 | 0 | 0 | 7 | ND | ND | ND |
| Fallow deer (n = 4) | Negative | Negative | 0 | 0 | 0 | 3 | ND | ND | ND |
| Red foxes (n = 10) | Ancylostomatidae (8) T canis (5) Capillaria (4) T vulpis (3) Coccidia (2) Hymenolepis (1) | Angiostrongylus (4) | 1 | 1 | 0 | 0 | 0 | 4 | 6 |
| Badgers (n = 5) | Ancylostomatidae (5) Capillaria (1) Coccidia (1) Strongyloides (1) | Crenosoma (4) | 2 | 0 | 0 | 1 | 0 | 1 | 0 |
| Porcupines (n = 4) | Negative | Negative | 1 | 1 | 0 | ND ^ | ND | ND | ND |
| Marten (n = 1) | Negative | Negative | 1 | 0 | 0 | ND | ND | ND | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pacini, M.I.; Mazzei, M.; Sgorbini, M.; D’Alfonso, R.; Papini, R.A. A One-Year Retrospective Analysis of Viral and Parasitological Agents in Wildlife Animals Admitted to a First Aid Hospital. Animals 2023, 13, 931. https://doi.org/10.3390/ani13050931
Pacini MI, Mazzei M, Sgorbini M, D’Alfonso R, Papini RA. A One-Year Retrospective Analysis of Viral and Parasitological Agents in Wildlife Animals Admitted to a First Aid Hospital. Animals. 2023; 13(5):931. https://doi.org/10.3390/ani13050931
Chicago/Turabian StylePacini, Maria Irene, Maurizio Mazzei, Micaela Sgorbini, Rossella D’Alfonso, and Roberto Amerigo Papini. 2023. "A One-Year Retrospective Analysis of Viral and Parasitological Agents in Wildlife Animals Admitted to a First Aid Hospital" Animals 13, no. 5: 931. https://doi.org/10.3390/ani13050931
APA StylePacini, M. I., Mazzei, M., Sgorbini, M., D’Alfonso, R., & Papini, R. A. (2023). A One-Year Retrospective Analysis of Viral and Parasitological Agents in Wildlife Animals Admitted to a First Aid Hospital. Animals, 13(5), 931. https://doi.org/10.3390/ani13050931