

First Insights on the Administration of Insect Oil (Black Soldier Fly Larvae) in the Diet of Juvenile Onychostoma macrolepis
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Experimental Diets
2.3. Fish and Feeding Conditions
2.4. Sampling
2.5. Methods of Detection and Analysis
2.5.1. Approximate Composition Detection
2.5.2. Determination of Fatty Acid Composition in Diet
2.5.3. Serum Biochemical Analysis
2.5.4. Antioxidant Activity Assays in the Liver
2.5.5. Intraperitoneal Fat Histology
2.5.6. Gene Expression
2.6. Statistical Analysis
3. Results
3.1. Growth and Biometric Indices
3.2. Approximate Composition of Whole Fish, Muscle and Liver
3.3. Serum Biochemical Indices
3.4. Antioxidant Activities
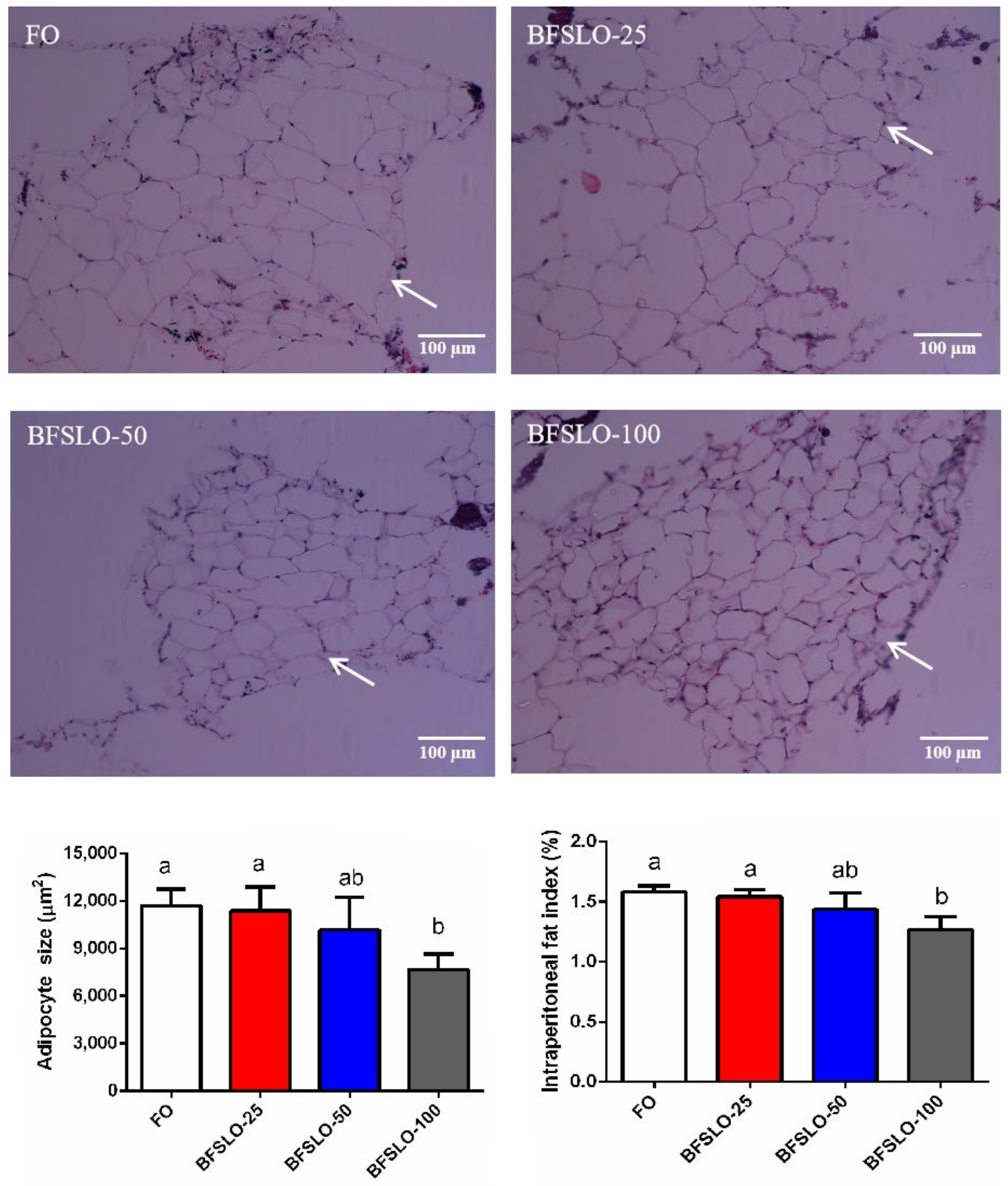
3.5. Intraperitoneal Fat Morphology
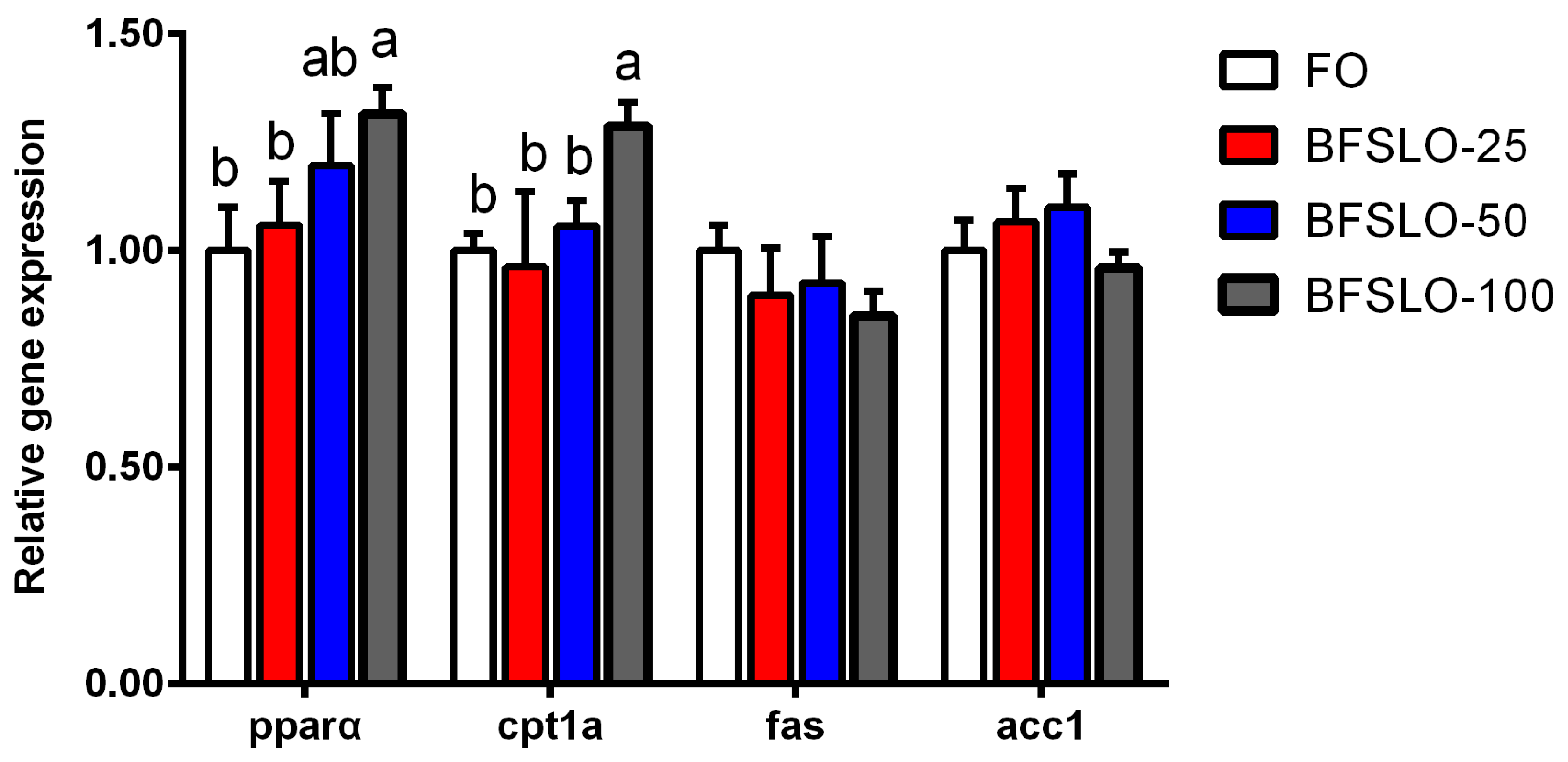
3.6. mRNA Expression of Lipid Metabolism-Related Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Spranghers, T.; Ottoboni, M.; Klootwiik, C.; Ovyn, A.; Deboosfere, S.; De Meulenaer, B.; Michiels, J.; Eeckhout, M.; De Clerq, P.; De Smet, S. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrate. J. Sci. Food Agric. 2017, 97, 2594–2600. [Google Scholar] [CrossRef]
- Benzertiha, A.; Kierończyk, B.; Rawski, M.; Mikolajczak, Z.; Urbański, A.; Nogowski, L.; Józefiak, D. Insect fat in animal nutrition—A review. Ann. Anim. Sci. 2020, 20, 1217–1240. [Google Scholar] [CrossRef]
- Tran, G.; Heuzé, V.; Makkar, H.P.S. Insects in fish diets. Anim. Front. 2015, 5, 37–44. [Google Scholar]
- Mohan, K.; Rajan, D.K.; Muralisankar, T.; Ganesan, A.R.; Sathishkumar, P.; Revathi, N. Use of black soldier fly (Hermetia illucens L.) larvae meal in aquafeeds for a sustainable aquaculture industry: A review of past and future needs. Aquaculture 2022, 553, 738095. [Google Scholar] [CrossRef]
- Gougbedji, A.; Agbohessou, P.; Lalèyè, P.; Francis, F.; Megido, R. Technical basis for the small-scale production of black soldier fly, Hermetia illucens (L. 1758), meal as fish feed in Benin. J. Agric. Food Res. 2021, 4, 100153. [Google Scholar] [CrossRef]
- Dumas, A.; Raggi, T.; Barkhouse, J.; Lewis, E.; Weltzien, E. The oil fraction and partially defatted meal of black soldier fly larvae (Hermetia illucens) affect differently growth performance, feed efficiency, nutrient deposition, blood glucose and lipid digestibility of rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 492, 24–34. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, J.; Song, Z.; Sun, Y. Optimization of extraction of proteins from larvae of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae), using response surface methodology. Acta. Entomol. Sin. 2014, 57, 401–410. [Google Scholar]
- Caimi, C.; Renna, M.; Lussiana, C.; Bonaldo, A.; Gariglio, M.; Meneguz, M.; Dabbou, S.; Schiavone, A.; Gai, F.; Elia, A.C.; et al. First insights on Black Soldier Fly (Hermetia illucens L.) larvae meal dietary administration in Siberian sturgeon (Acipenser baerii Brandt) juveniles. Aquaculture 2020, 515, 734539. [Google Scholar] [CrossRef]
- Kishawy, A.; Mohammed, H.; Zaglool, A.; Attia, M.; Hassan, F.; Roushdy, E.; Ismail, T.; Ibrahim, D. Partial defatted black solider larvae meal as a promising strategy to replace fish meal protein in diet for Nile tilapia (Oreochromis niloticus): Performance, expression of protein and fat transporters, and cytokines related genes and economic efficiency. Aquaculture 2022, 555, 738195. [Google Scholar] [CrossRef]
- Hender, A.; Siddik, M.A.B.; Howieson, J.; Fotedar, R. Black soldier fly, Hermetia illucens as an alternative to fishmeal protein and fish oil: Impact on growth, immune response, mucosal barrier status, and flesh quality of juvenile barramundi, Lates calcarifer (Bloch, 1790). Biology 2021, 10, 505. [Google Scholar] [CrossRef]
- Homska, N.; Kowalska, J.; Bogucka, J.; Ziółkowska, E.; Rawski, M.; Kierończyk, B.; Mazurkiewicz, J. Dietary Fish Meal Replacement with Hermetia illucens and Tenebrio molitor Larval Meals Improves the Growth Performance and Nutriphysiological Status of Ide (Leuciscus idus) Juveniles. Animals 2022, 12, 1227. [Google Scholar] [CrossRef]
- Cardinaletti, G.; Marco, P.D.; Daniso, E.; Messina, M.; Donadelli, V.; Finoia, M.G.; Petochi, T.; Fava, F.; Faccenda, F.; Contò, M.; et al. Growth and Welfare of Rainbow Trout (Oncorhynchus mykiss) in Response to Graded Levels of Insect and Poultry By-Product Meals in Fishmeal-Free Diets. Animals 2022, 12, 1698. [Google Scholar] [CrossRef] [PubMed]
- Gougbedji, A.; Detilleux, J.; Lalèyè, P.A.; Francis, F.; Megido, R.C. Can Insect Meal Replace Fishmeal? A Meta-Analysis of the Effects of Black Soldier Fly on Fish Growth Performances and Nutritional Values. Animals 2022, 12, 1700. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ji, H.; Zhang, B.; Tian, J.; Zhou, J.; Yu, H. Influence of black soldier fly (Hermetia illucens) larvae oil on growth performance, body composition, tissue fatty acid composition and lipid deposition in juvenile Jian carp (Cyprinus carpio var. Jian). Aquaculture 2016, 465, 43–52. [Google Scholar] [CrossRef]
- Schönfeld, P.; Wojtczak, L. Short- and medium-chain fatty acids in energy metabolism: The cellular perspective. J. Lipid Res. 2016, 57, 943–954. [Google Scholar] [CrossRef]
- Xu, X.; Ji, H.; Belghit, I.; Sun, J. Black soldier fly larvae as a better lipid source than yellow mealworm or silkworm oils for juvenile mirror carp (Cyprinus carpio var. specularis). Aquaculture 2020, 527, 735453. [Google Scholar] [CrossRef]
- Marques, V.H.; Moreira, R.G.; Branco, G.S.; Honji, R.M.; Rombenso, A.N.; Viana, M.T.; Araújo, B.C. Different saturated and monounsaturated fatty acids levels in fish oil-free diets to cobia (Rachycentron canadum) juveniles: Effects in growth performance and lipid metabolism. Aquaculture 2021, 541, 736843. [Google Scholar] [CrossRef]
- Xu, X.; Ji, H.; Belghit, I.; Liland, N.; Wu, W.; Li, X. Effects of black soldier fly oil rich in n-3 HUFA on growth performance, metabolism and health response of juvenile mirror carp (Cyprinus carpio var. specularis). Aquaculture 2021, 533, 736144. [Google Scholar] [CrossRef]
- Fawole, F.; Labh, S.; Hossain, M.; Overturf, K.; Small, B.; Welker, T.; Hardy, R.; Kumar, V. Insect (black soldier fly larvae) oil as a potential substitute for fifish or soy oil in the fish meal-based diet of juvenile rainbow trout (Oncorhynchus mykiss). Anim. Nutr. 2021, 7, 1360–1370. [Google Scholar] [CrossRef]
- Maldonado-Othón, C.; Re-Vega, E.; Perez-Velazquez, M.; González-Félix, M. Replacement of fish oil by camelina and black soldier fly larvae oils in diets for juvenile Totoaba macdonaldi and their effect on growth, fatty acid profile, and gene expression of pancreatic lipases. Aquaculture 2022, 552, 737985. [Google Scholar] [CrossRef]
- Gou, N.N.; Ji, H.; Zhong, M.Z.; Chang, Z.G.; Deng, W. Effects of dietary fish oil replacements with three vegetable oils on growth, fatty acid composition, antioxidant capacity, serum parameters and expression of lipid metabolism related genes in juvenile Onychostoma macrolepis. Aquac. Nutr. 2020, 27, 163–175. [Google Scholar] [CrossRef]
- AOAC. Official Methods for Analysis, 19th ed.; Association of Official Analytical Chemists: Rockville, MD, USA, 2012. [Google Scholar]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Osman, O.S.; Selway, J.L.; Kępczyńska, M.A.; Stocker, C.J.; O’Dowd, J.F.; Cawthorne, M.A.; Arch, J.R.; Jassim, S.; Langlands, K. A novel automated image analysis method for accurate adipocyte quantification. Adipocyte 2013, 2, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Gou, N.N.; Ji, H.; Chang, Z.G.; Zhon, M.Z.; Deng, W. Effects of dietary essential fatty acid requirements on growth performance, fatty acid composition, biochemical parameters, antioxidant response and lipid related genes expression in juvenile Onychostoma macrolepis . Aquaculture 2020, 528, 735590. [Google Scholar] [CrossRef]
- Tocher, D.R. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Han, J.; Hamilton, J.A.; Kirkland, J.L.; Corkey, B.E.; Guo, W. Medium chain oil reduces fat mass and down regulates expression of adipogenic genes in rats. Obes. Res. 2003, 11, 734–744. [Google Scholar] [CrossRef]
- Zheng, J.L.; Luo, Z.; Zhu, Q.L.; Tan, X.Y.; Chen, Q.L.; Sun, L.D.; Hu, W. Molecular cloning and expression pattern of 11 genes involved in lipid metabolism in yellow catfish Pelteobagrus fulvidraco . Gene 2013, 531, 53–63. [Google Scholar] [CrossRef]
- Fawole, F.J.; Adeoye, A.A.; Tiamiyu, L.O.; Ajala, K.I.; Obadara, S.O.; Ganiyu, I.O. Substituting fishmeal with Hermetia illucens in the diets of African catfish (Clarias gariepinus): Effects on growth, nutrient utilization, haematophysiological response, and oxidative stress biomarker. Aquaculture 2020, 518, 734849. [Google Scholar] [CrossRef]
- Sankiana, Z.; Khosravia, S.; Kimb, Y.O.; Lee, S.M. Effects of dietary inclusion of yellow mealworm (Tenebrio molitor) meal on growth performance, feed utilization, body composition, plasma biochemical indices, selected immune parameters and antioxidant enzyme activities of mandarin fish (Siniperca scherzeri) juveniles. Aquaculture 2018, 496, 79–87. [Google Scholar]
- Abdel-Tawwab, M.; Khalil, R.H.; Metwally, A.A.; Shakweer, M.S.; Khallaf, M.A.; Abdel-Latif, H.M.R. Effects of black soldier fly (Hermetia illucens L.) larvae meal on growth performance, organs-somatic indices, body composition, and hematobiochemical variables of European sea bass, Dicentrarchus labrax. Aquaculture 2020, 522, 735136. [Google Scholar]
- Moutinho, S.; Pedrosa, R.; Magalhães, R.; Oliva-Teles, A.; Parisi, G.; Peres, H. Black soldier fly (Hermetia illucens) pre-pupae larvae meal in diets for European seabass (Dicentrarchus labrax) juveniles: Effects on liver oxidative status and fillet quality traits during shelf-life. Aquaculture 2021, 533, 736080. [Google Scholar] [CrossRef]
- Yadav, A.K.; Rossi, W., Jr.; Habte-Tsion, H.M.; Kumar, V. Impacts of dietary eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) level and ratio on the growth, fatty acids composition and hepaticantioxidant status of largemouth bass (Micropterus salmoides). Aquaculture 2020, 529, 735683. [Google Scholar] [CrossRef]
- Koruk, M.; Taysi, S.; Savas, M.C.; Yilmaz, O.; Akcay, F.; Karakok, M. Oxidative stress and enzymatic antioxidant status in patients with nonalcoholic steatohepatitis. Ann. Clin. Lab. Sci. 2004, 34, 57–62. [Google Scholar] [PubMed]
- Ratti, S.; Zarantoniello, M.; Chemello, G.; Giammarino, M.; Palermo, F.A.; Cocci, P.; Mosconi, G.; Tignani, M.V.; Pascon, G.; Cardinaletti, G.; et al. Spirulina-enriched Substrate to Rear Black Soldier Fly (Hermetia illucens) Prepupae as Alternative Aquafeed Ingredient for Rainbow Trout (Oncorhynchus mykiss) Diets: Possible Effects on Zootechnical Performances, Gut and Liver Health Status, and Fillet Quality. Animals 2023, 13, 173. [Google Scholar]
- Guerreiro, I.; Castro, C.; Serra, C.R.; Coutinho, F.; Couto, A.; Peres, H.; Pousão-Ferreira, P.; Gasco, L.; Gai, F.; Oliva-Teles, A.; et al. Oxidative Stress Response of Meagre to Dietary Black Soldier Fly Meal. Animals 2022, 12, 3232. [Google Scholar] [CrossRef]
- Melenchón, F.; Mercado, E.; Pula, H.; Cardenete, G.; Barroso, F.G.; Fabrikov, D.; Lourenço, H.M.; Pessoa, M.F.; Lagos, L.; Weththasinghe, P.; et al. Fishmeal Dietary Replacement Up to 50%: A Comparative Study of Two Insect Meals for Rainbow Trout (Oncorhynchus mykiss). Animals 2022, 12, 179. [Google Scholar] [CrossRef]
- Elia, A.; Capucchio, M.; Caldaroni, B.; Magara, G.; Dorr, A.; Biasato, I.; Biasibetti, E.; Righetti, M.; Pastorino, P.; Prearo, M.; et al. Influence of Hermetia illucens meal dietary inclusion on the histological traits, gutmucin composition and the oxidative stress biomarkers in rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 496, 50–57. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Ingredients | FO | BSFLO-25 | BSFLO-50 | BSFLO-100 |
|---|---|---|---|---|
| Casein | 350 | 350 | 350 | 350 |
| Gelatin | 104 | 104 | 104 | 104 |
| Dextrin | 270 | 270 | 270 | 270 |
| Microcrystalline cellulose | 115 | 115 | 115 | 115 |
| Vitamin premix 1 | 10 | 10 | 10 | 10 |
| Mineral misture 2 | 40 | 40 | 40 | 40 |
| Carboxymethylcellulose | 20 | 20 | 20 | 20 |
| BHT | 1 | 1 | 1 | 1 |
| FO 3 | 90 | 67.5 | 45 | 0 |
| BSFLO 4 | 0 | 22.5 | 45 | 90 |
| Total | 1000 | 1000 | 1000 | 1000 |
| Dry matter (%) | 90.09 | 89.52 | 90.03 | 89.71 |
| Crude protein (%) | 37.11 | 37.25 | 36.97 | 37.34 |
| Crude fat (%) | 9.35 | 9.19 | 9.22 | 9.16 |
| Ash (%) | 10.82 | 10.13 | 10.79 | 10.99 |
| NFE 5 (%) | 32.81 | 32.95 | 33.05 | 32.22 |
| GE 6 (MJ/kg) | 18.37 | 18.36 | 18.32 | 18.24 |
| Fatty Acids | FO | BFSLO-25 | BFSLO-50 | BFSLO-100 |
|---|---|---|---|---|
| C12:0 | 0.00 | 6.30 | 12.46 | 25.25 |
| C14:0 | 1.94 | 1.77 | 1.79 | 3.14 |
| C16:0 | 17.65 | 18.80 | 17.63 | 19.61 |
| C18:0 | 5.78 | 5.21 | 4.15 | 4.01 |
| C16:1n-7 | 1.10 | 1.31 | 1.26 | 1.97 |
| C18:1n-9 | 13.98 | 13.86 | 14.19 | 14.05 |
| C18:2n-6 | 4.14 | 6.01 | 8.43 | 10.15 |
| C18:3n-6 | 0.68 | 0.70 | 0.82 | 1.07 |
| C20:3n-6 | 0.50 | 0.20 | 0.70 | 1.00 |
| C20:4n-6 | 0.73 | 1.10 | 1.17 | 1.26 |
| C18:3n-3 | 2.94 | 2.69 | 1.53 | 0.27 |
| C20:3n-3 | 0.28 | 0.24 | 0.18 | 0.00 |
| C20:5n-3 | 21.27 | 17.07 | 12.93 | 0.00 |
| C22:6n-3 | 14.53 | 12.05 | 10.96 | 0.00 |
| ∑SFA 1 | 25.37 | 32.08 | 36.03 | 52.01 |
| ∑MUFA 2 | 15.08 | 15.17 | 15.45 | 16.02 |
| ∑n-6 PUFA | 6.05 | 8.01 | 11.12 | 13.48 |
| ∑n-3 PUFA | 39.02 | 32.05 | 25.60 | 0.27 |
| ∑PUFA 3 | 45.07 | 40.06 | 36.72 | 13.75 |
| n-3/n-6 PUFA | 6.45 | 4.00 | 2.30 | 0.02 |
| Gene | Gene Bank Accession No. | Sequence |
|---|---|---|
| pparα 1 | MG735214.1 | F:TGACATGGAGGTGCTGGAGGAC |
| R:TGCTGCTGTGCTGTTGCTCTG | ||
| cpt1a 2 | MH553647 | F:CTCAGACGGTGTTCAGTGCCATC |
| R:TCCAGCCGTGATAGGACAAGAGG | ||
| fas 3 | MG735211.1 | F:ATCCACAGAGCCACCATCCTACC |
| R:CAAGTCCAGCATCCTCCAAGACAC | ||
| acc1 4 | MG735212.1 | F:AGGTGGTACGGATGGCTGCTC |
| R:GACGGTGCTGGACGCTGTTG | ||
| β-actin5 | JN254630.1 | F:TGACCCACACTGTACCCATC |
| R:CGGACAATTTCACTCTCGGC |
| FO | BFSLO-25 | BFSLO-50 | BFSLO-100 | |
|---|---|---|---|---|
| IBW 1 (g) | 1.75 ± 0.04 | 1.74 ± 0.02 | 1.76 ± 0.03 | 1.77 ± 0.02 |
| FBW 2 (g) | 9.26 a ± 0.20 | 9.07 a ± 0.18 | 8.81 ab ± 0.17 | 8.39 b ± 0.14 |
| WG 3 (%) | 429.57 a ± 14.27 | 422.59 a ± 15.93 | 401.34 ab ± 4.08 | 373.78 b ± 3.30 |
| SGR 4 (%/day) | 2.98 a ± 0.05 | 2.95 a ± 0.06 | 2.88 ab ± 0.02 | 2.78 b ± 0.02 |
| FI 5 (g/fish) | 11.61 a ± 0.15 | 11.28 ab ± 0.12 | 11.01 ab ± 0.26 | 10.68 b ± 0.21 |
| FCR 6 | 1.55 ± 0.06 | 1.54 ± 0.03 | 1.56 ± 0.01 | 1.61 ± 0.02 |
| VSI 7 (%) | 8.86 ± 0.26 | 8.66 ± 0.31 | 8.55 ± 0.21 | 8.19 ± 0.13 |
| HSI 8 (%) | 1.51 ± 0.03 | 1.54 ± 0.03 | 1.55 ± 0.05 | 1.51 ± 0.08 |
| IPFI 9 (%) | 1.58 a ± 0.03 | 1.54 a ± 0.03 | 1.44 ab ± 0.13 | 1.27 b ± 0.11 |
| FO | BFSLO-25 | BFSLO-50 | BFSLO-100 | |
|---|---|---|---|---|
| Whole dody | ||||
| Moisture | 70.27 ± 0.88 | 70.29 ± 0.76 | 70.33 ± 0.96 | 71.2 ± 0.67 |
| Crude protein | 16.44 ± 0.67 | 16.25 ± 0.69 | 16.37 ± 0.84 | 16.13 ± 0.30 |
| Crude lipid | 10.82 a ± 0.19 | 10.41 a ± 0.16 | 10.36 a ± 0.15 | 9.63 b ± 0.13 |
| Ash | 2.31 ± 0.14 | 2.55 ± 0.12 | 2.49 ± 0.15 | 2.85 ± 0.22 |
| Muscle | ||||
| Moisture | 77.18 ± 0.61 | 77.55 ± 0.68 | 77.2 ± 0.53 | 77.94 ± 0.85 |
| Crude protein | 19.15 ± 0.29 | 19.26 ± 0.26 | 19.45 ± 0.21 | 19.36 ± 0.23 |
| Crude lipid | 1.41 ± 0.14 | 1.38 ± 0.08 | 1.25 ± 0.06 | 1.18 ± 0.05 |
| Ash | 1.33 ± 0.08 | 1.29 ± 0.08 | 1.27 ± 0.10 | 1.24 ± 0.08 |
| Liver | ||||
| Moisture | 61.06 ± 1.03 | 61.33 ± 0.84 | 61.45 ± 1.12 | 62.64 ± 0.69 |
| Crude protein | 15.74 ± 0.65 | 15.53 ± 1.00 | 15.2 ± 0.58 | 15.35 ± 0.54 |
| Crude lipid | 21.59 ± 0.81 | 21.85 ± 1.63 | 21.81 ± 1.80 | 20.11 ± 1.68 |
| Ash | 1.08 ± 0.02 | 1.13 ± 0.08 | 1.06 ± 0.03 | 1.05 ± 0.03 |
| FO | BFSLO-25 | BFSLO-50 | BFSLO-100 | |
|---|---|---|---|---|
| ALB 1 (g/L) | 6.14 ± 0.10 | 6.13 ± 0.14 | 6.15 ± 0.21 | 6.21 ± 0.10 |
| GLO 2 (g/L) | 11.76 ± 0.51 | 12.04 ± 0.73 | 10.95 ± 0.41 | 11.25 ± 0.21 |
| TP 3 (g/L) | 17.9 ± 0.55 | 18.17 ± 0.59 | 17.1 ± 0.61 | 17.47 ± 0.28 |
| CHOL 4 (mmol/L) | 1.65 ± 0.07 | 1.75 ± 0.08 | 1.65 ± 0.08 | 1.64 ± 0.10 |
| TG 5 (mmol/L) | 1.73 a ± 0.10 | 1.74 a ± 0.14 | 1.58 ab ± 0.03 | 1.36 b ± 0.03 |
| HDL 6 (mmol/L) | 0.56 ± 0.05 | 0.64 ± 0.06 | 0.63 ± 0.06 | 0.62 ± 0.06 |
| LDL 7 (mmol/L) | 0.43 ± 0.05 | 0.41 ± 0.03 | 0.39 ± 0.02 | 0.36 ± 0.03 |
| AST 8 (U/L) | 169.87 b ± 4.29 | 166.67 b ± 3.84 | 168.27 b ± 6.58 | 188.67 a ± 4.91 |
| ALT 9 (U/L) | 83.00 b ± 4.59 | 81.50 b ± 3.14 | 82.10 b ± 3.86 | 96.07 a ± 2.81 |
| FO | BFSLO-25 | BFSLO-50 | BFSLO-100 | |
|---|---|---|---|---|
| SOD 1 (U/mgprot) | 81.18 b ± 2.33 | 94.96 a ± 2.77 | 94.34 a ± 3.27 | 91.22 a ± 1.29 |
| GPx 2 (U/mgprot) | 20.91 ± 1.46 | 28.98 ± 4.82 | 33.15 ± 4.22 | 26.15 ± 3.77 |
| CAT 3 (U/mgprot) | 17.6 ± 0.92 | 19.3 ± 0.73 | 18.08 ± 1.62 | 19.74 ± 2.15 |
| MDA 4 (nmol/mgprot) | 1.48 a ± 0.11 | 1.15 b ± 0.08 | 1.16 b ± 0.06 | 1.12 b ± 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gou, N.; Wang, K.; Jin, T.; Yang, B. First Insights on the Administration of Insect Oil (Black Soldier Fly Larvae) in the Diet of Juvenile Onychostoma macrolepis. Animals 2023, 13, 518. https://doi.org/10.3390/ani13030518
Gou N, Wang K, Jin T, Yang B. First Insights on the Administration of Insect Oil (Black Soldier Fly Larvae) in the Diet of Juvenile Onychostoma macrolepis. Animals. 2023; 13(3):518. https://doi.org/10.3390/ani13030518
Chicago/Turabian StyleGou, Nina, Kaifeng Wang, Tiezhi Jin, and Bin Yang. 2023. "First Insights on the Administration of Insect Oil (Black Soldier Fly Larvae) in the Diet of Juvenile Onychostoma macrolepis" Animals 13, no. 3: 518. https://doi.org/10.3390/ani13030518
APA StyleGou, N., Wang, K., Jin, T., & Yang, B. (2023). First Insights on the Administration of Insect Oil (Black Soldier Fly Larvae) in the Diet of Juvenile Onychostoma macrolepis. Animals, 13(3), 518. https://doi.org/10.3390/ani13030518