


Inhibition of Cell Apoptosis by Apicomplexan Protozoa–Host Interaction in the Early Stage of Infection

Abstract
:Simple Summary
Abstract
1. Introduction
2. Apoptosis
3. Apicomplexan Protozoa Inhibit Cell Apoptosis in the Early Stage of Infection
4. Virulence Factors Associated with Apicomplexan Protozoa Inhibit Host Cell Apoptosis
4.1. Microneme Proteins
4.2. Rhoptry Proteins
4.3. Dense Granule Proteins
4.4. Heat Shock Proteins
5. Apicomplexan Protozoa Infection Effects on Key Apoptotic Proteins and Pathways in the Host
5.1. Cysteine Protease Family
5.2. Bcl-2 Protein Family
5.3. Cytochrome-C
5.4. NF-κB Signaling Pathway
5.5. Phosphatidylinositol 3-Kinase-Akt Signaling Pathway
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sparvoli, D.; Lebrun, M. Unraveling the Elusive Rhoptry Exocytic Mechanism of Apicomplexa. Trends Parasitol. 2021, 37, 622–637. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.S.; Walia, A.K.; Kanwar, J.R. Protozoa lectins and their role in host-pathogen interactions. Biotechnol. Adv. 2016, 34, 1018–1029. [Google Scholar] [CrossRef]
- Guillermo, L.V.; Pereira, W.F.; De Meis, J.; Ribeiro-Gomes, F.L.; Silva, E.M.; Kroll-Palhares, K.; Takiya, C.M.; Lopes, M.F. Targeting caspases in intracellular protozoan infections. Immunopharmacol. Immunotoxicol. 2009, 31, 159–173. [Google Scholar] [CrossRef]
- Gervais, O.; Renault, T.; Arzul, I. Molecular and cellular characterization of apoptosis in flat oyster a key mechanisms at the heart of host-parasite interactions. Sci. Rep. 2018, 8, 12494. [Google Scholar] [CrossRef] [PubMed]
- Sinai, A.P.; Payne, T.M.; Carmen, J.C.; Hardi, L.; Watson, S.J.; Molestina, R.E. Mechanisms underlying the manipulation of host apoptotic pathways by Toxoplasma gondii. Int. J. Parasitol. 2004, 34, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Jia, Y.; Han, Y.; Wang, W.; Zhu, Y.; Xu, J.; Guan, C.; Ying, J.; Deng, S.; Wang, J.; et al. Eimeria acervulina Microneme Protein 3 Inhibits Apoptosis of the Chicken Duodenal Epithelial Cell by Targeting the Casitas B-Lineage Lymphoma Protein. Front. Vet. Sci. 2021, 8, 636809. [Google Scholar] [CrossRef]
- del Cacho, E.; Gallego, M.; López-Bernad, F.; Quílez, J.; Sánchez-Acedo, C. Expression of anti-apoptotic factors in cells parasitized by second-generation schizonts of Eimeria tenella and Eimeria necatrix. Vet. Parasitol. 2004, 125, 287–300. [Google Scholar] [CrossRef]
- DosReis, G.A.; Barcinski, M.A. Apoptosis and parasitism: From the parasite to the host immune response. Adv. Parasitol. 2001, 49, 133–161. [Google Scholar] [CrossRef]
- Sorice, M. Crosstalk of Autophagy and Apoptosis. Cells 2022, 11, 1479. [Google Scholar] [CrossRef]
- Marek, L. The role of the apoptosome in the activation of procaspase-9. Postep. Hig. Med. Dosw. Online 2013, 67, 54–64. [Google Scholar] [CrossRef]
- Fan, T.J.; Han, L.H.; Cong, R.S.; Liang, J. Caspase family proteases and apoptosis. Acta Biochim. Biophys. Sin. 2005, 37, 719–727. [Google Scholar] [CrossRef]
- Cheng, S.Y.; Leonard, J.L.; Davis, P.J. Molecular aspects of thyroid hormone actions. Endocr. Rev. 2010, 31, 139–170. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Zhao, Q.; Zhu, S.; Han, H.; Dong, H.; Huang, B. Establishment of Eimeria tenella (local isolate) in chicken embryos. Parasite 2012, 19, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Fellenberg, J.; Wang, H.; Carstens, C.; Richter, W. Occurrence and regional distribution of apoptosis in scoliotic discs. Spine 2005, 30, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Lossi, L. The concept of intrinsic versus extrinsic apoptosis. Biochem. J. 2022, 479, 357–384. [Google Scholar] [CrossRef]
- Xu, X.; Lai, Y.; Hua, Z.C. Apoptosis and apoptotic body: Disease message and therapeutic target potentials. Biosci. Rep. 2019, 39, BSR20180992. [Google Scholar] [CrossRef]
- Bosurgi, L.; Rothlin, C.V. Management of cell death in parasitic infections. Semin. Immunopathol. 2021, 43, 481–492. [Google Scholar] [CrossRef]
- Huh, C.G.; Factor, V.M.; Sánchez, A.; Uchida, K.; Conner, E.A.; Thorgeirsson, S.S. Hepatocyte growth factor/c-met signaling pathway is required for efficient liver regeneration and repair. Proc. Natl. Acad. Sci. USA 2004, 101, 4477–4482. [Google Scholar] [CrossRef]
- Leirião, P.; Albuquerque, S.S.; Corso, S.; van Gemert, G.J.; Sauerwein, R.W.; Rodriguez, A.; Giordano, S.; Mota, M.M. HGF/MET signalling protects Plasmodium-infected host cells from apoptosis. Cell Microbiol. 2005, 7, 603–609. [Google Scholar] [CrossRef]
- van de Sand, C.; Horstmann, S.; Schmidt, A.; Sturm, A.; Bolte, S.; Krueger, A.; Lütgehetmann, M.; Pollok, J.M.; Libert, C.; Heussler, V.T. The liver stage of Plasmodium berghei inhibits host cell apoptosis. Mol. Microbiol. 2005, 58, 731–742. [Google Scholar] [CrossRef]
- Mele, R.; Gomez Morales, M.A.; Tosini, F.; Pozio, E. Cryptosporidium parvum at different developmental stages modulates host cell apoptosis in vitro. Infect. Immun. 2004, 72, 6061–6067. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.M.; Levine, S.A.; Splinter, P.L.; Tietz, P.S.; Ganong, A.L.; Jobin, C.; Gores, G.J.; Paya, C.V.; LaRusso, N.F. Cryptosporidium parvum activates nuclear factor kappaB in biliary epithelia preventing epithelial cell apoptosis. Gastroenterology 2001, 120, 1774–1783. [Google Scholar] [CrossRef] [PubMed]
- Tenter, A.M.; Heckeroth, A.R.; Weiss, L.M. Toxoplasma gondii: From animals to humans. Int. J. Parasitol. 2000, 30, 1217–1258. [Google Scholar] [CrossRef]
- Xu, Z.Y.; Zheng, M.X.; Zhang, Y.; Cui, X.Z.; Yang, S.S.; Liu, R.L.; Li, S.; Lv, Q.H.; Zhao, W.L.; Bai, R. The effect of the mitochondrial permeability transition pore on apoptosis in Eimeria tenella host cells. Poult. Sci. 2016, 95, 2405–2413. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.S.; Zhao, Y.J.; Zhang, Y.; Xu, T.; Cui, K.L.; Duan, B.T.; Lv, X.L.; Zhang, L.; Xu, Z.Y.; Bai, R.; et al. Role of EtMIC4 EGF-like in regulating the apoptosis of Eimeria tenella host cells via the EGFR pathway. Poult. Sci. 2022, 101, 102075. [Google Scholar] [CrossRef]
- Carruthers, V.B.; Tomley, F.M. Microneme proteins in apicomplexans. Subcell. Biochem. 2008, 47, 33–45. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, F.; Zhang, D.; Liu, D.; Ding, W.; Springer, T.A.; Song, G. Structural insights into MIC2 recognition by MIC2-associated protein in Toxoplasma gondii. Commun. Biol. 2023, 6, 895. [Google Scholar] [CrossRef]
- Tomley, F.M.; Billington, K.J.; Bumstead, J.M.; Clark, J.D.; Monaghan, P. EtMIC4: A microneme protein from Eimeria tenella that contains tandem arrays of epidermal growth factor-like repeats and thrombospondin type-I repeats. Int. J. Parasitol. 2001, 31, 1303–1310. [Google Scholar] [CrossRef]
- Lu, X.; An, L.; Fan, G.; Zang, L.; Huang, W.; Li, J.; Liu, J.; Ge, W.; Huang, Y.; Xu, J.; et al. EGFR signaling promotes nuclear translocation of plasma membrane protein TSPAN8 to enhance tumor progression via STAT3-mediated transcription. Cell Res. 2022, 32, 359–374. [Google Scholar] [CrossRef]
- Muniz-Feliciano, L.; Van Grol, J.; Portillo, J.A.; Liew, L.; Liu, B.; Carlin, C.R.; Carruthers, V.B.; Matthews, S.; Subauste, C.S. Toxoplasma gondii-induced activation of EGFR prevents autophagy protein-mediated killing of the parasite. PLoS Pathog. 2013, 9, e1003809. [Google Scholar] [CrossRef]
- Meissner, M.; Reiss, M.; Viebig, N.; Carruthers, V.B.; Toursel, C.; Tomavo, S.; Ajioka, J.W.; Soldati, D. A family of transmembrane microneme proteins of Toxoplasma gondii contain EGF-like domains and function as escorters. J. Cell Sci. 2002, 115 Pt 3, 563–574. [Google Scholar] [CrossRef]
- Cova, M.M.; Lamarque, M.H.; Lebrun, M. How Apicomplexa Parasites Secrete and Build Their Invasion Machinery. Annu. Rev. Microbiol. 2022, 76, 619–640. [Google Scholar] [CrossRef]
- Wu, L.; Wu, L.; Xi, C.; Liu, Y.; Jiang, X.; Chen, S.; Cao, J. Toxoplasma Gondii Rhoptry Protein 16 (ROP16) Modifies Apoptosis in Human 293T Cells. J. Nanosci. Nanotechnol. 2020, 20, 24–30. [Google Scholar] [CrossRef]
- Yamamoto, M.; Standley, D.M.; Takashima, S.; Saiga, H.; Okuyama, M.; Kayama, H.; Kubo, E.; Ito, H.; Takaura, M.; Matsuda, T.; et al. A single polymorphic amino acid on Toxoplasma gondii kinase ROP16 determines the direct and strain-specific activation of Stat3. J. Exp. Med. 2009, 206, 2747–2760. [Google Scholar] [CrossRef]
- Butcher, B.A.; Fox, B.A.; Rommereim, L.M.; Kim, S.G.; Maurer, K.J.; Yarovinsky, F.; Herbert, D.R.; Bzik, D.J.; Denkers, E.Y. Toxoplasma gondii rhoptry kinase ROP16 activates STAT3 and STAT6 resulting in cytokine inhibition and arginase-1-dependent growth control. PLoS Pathog. 2011, 7, e1002236. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; An, R.; Chen, L.; Shen, Y.; Chen, Y.; Cheng, L.; Jiang, Z.; Zhang, A.; Yu, L.; Chu, D.; et al. Toxoplasma gondii virulence factor ROP18 inhibits the host NF-κB pathway by promoting p65 degradation. J. Biol. Chem. 2014, 289, 12578–12592. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, S.; Saeij, J.P.J. Toxoplasma Does Not Secrete the GRA16 and GRA24 Effectors Beyond the Parasitophorous Vacuole Membrane of Tissue Cysts. Front. Cell Infect. Microbiol. 2018, 8, 366. [Google Scholar] [CrossRef] [PubMed]
- Ahmadpour, E.; Babaie, F.; Kazemi, T.; Mehrani Moghaddam, S.; Moghimi, A.; Hosseinzadeh, R.; Nissapatorn, V.; Pagheh, A.S. Overview of Apoptosis, Autophagy, and Inflammatory Processes in Toxoplasma gondii Infected Cells. Pathogens 2023, 12, 253. [Google Scholar] [CrossRef]
- Thind, A.C.; Mota, C.M.; Goncalves, A.P.N.; Sha, J.; Wohlschlegel, J.A.; Mineo, T.W.P.; Bradley, P.J. The Toxoplasma gondii effector GRA83 modulates the host’s innate immune response to regulate parasite infection. bioRxiv 2023. [Google Scholar] [CrossRef] [PubMed]
- Matta, S.K.; Olias, P.; Huang, Z.; Wang, Q.; Park, E.; Yokoyama, W.M.; Sibley, L.D. Toxoplasma gondii effector TgIST blocks type I interferon signaling to promote infection. Proc. Natl. Acad. Sci. USA 2019, 116, 17480–17491. [Google Scholar] [CrossRef]
- Ching, X.T.; Lau, Y.L.; Fong, M.Y. Heterologous expression of Toxoplasma gondii dense granule protein 2 and 5. Southeast Asian J. Trop. Med. Public Health 2015, 46, 375–387. [Google Scholar]
- Braun, L.; Brenier-Pinchart, M.P.; Yogavel, M.; Curt-Varesano, A.; Curt-Bertini, R.L.; Hussain, T.; Kieffer-Jaquinod, S.; Coute, Y.; Pelloux, H.; Tardieux, I.; et al. A Toxoplasma dense granule protein, GRA24, modulates the early immune response to infection by promoting a direct and sustained host p38 MAPK activation. J. Exp. Med. 2013, 210, 2071–2086. [Google Scholar] [CrossRef] [PubMed]
- Mota, C.M.; Oliveira, A.C.; Davoli-Ferreira, M.; Silva, M.V.; Santiago, F.M.; Nadipuram, S.M.; Vashisht, A.A.; Wohlschlegel, J.A.; Bradley, P.J.; Silva, J.S.; et al. Neospora caninum Activates p38 MAPK as an Evasion Mechanism against Innate Immunity. Front. Microbiol. 2016, 7, 1456. [Google Scholar] [CrossRef] [PubMed]
- Sangaré, L.O.; Yang, N.; Konstantinou, E.K.; Lu, D.; Mukhopadhyay, D.; Young, L.H.; Saeij, J.P.J. Toxoplasma GRA15 Activates the NF-κB Pathway through Interactions with TNF Receptor-Associated Factors. mBio. 2019, 10, e00808-19. [Google Scholar] [CrossRef] [PubMed]
- Kim, L.; Denkers, E.Y. Toxoplasma gondii triggers Gi-dependent PI 3-kinase signaling required for inhibition of host cell apoptosis. J. Cell Sci. 2006, 119, 2119–2126. [Google Scholar] [CrossRef] [PubMed]
- Shonhai, A.; Maier, A.G.; Przyborski, J.M.; Blatch, G.L. Intracellular protozoan parasites of humans: The role of molecular chaperones in development and pathogenesis. Protein Pept. Lett. 2011, 18, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Hagymasi, A.T.; Dempsey, J.P.; Srivastava, P.K. Heat-Shock Proteins. Curr. Protoc. 2022, 2, e592. [Google Scholar] [CrossRef]
- Lee, J.; Roberts, J.S.; Atanasova, K.R.; Chowdhury, N.; Yilmaz, Ö. A novel kinase function of a nucleoside-diphosphate-kinase homologue in Porphyromonas gingivalis is critical in subversion of host cell apoptosis by targeting heat-shock protein 27. Cell Microbiol. 2018, 20, e12825. [Google Scholar] [CrossRef]
- Mosser, D.D.; Caron, A.W.; Bourget, L.; Meriin, A.B.; Sherman, M.Y.; Morimoto, R.I.; Massie, B. The chaperone function of hsp70 is required for protection against stress-induced apoptosis. Mol. Cell Biol. 2000, 20, 7146–7159. [Google Scholar] [CrossRef]
- Saleh, A.; Srinivasula, S.M.; Balkir, L.; Robbins, P.D.; Alnemri, E.S. Negative regulation of the Apaf-1 apoptosome by Hsp70. Nat. Cell Biol. 2000, 2, 476–483. [Google Scholar] [CrossRef]
- Cho, S.G.; Choi, E.J. Apoptotic signaling pathways: Caspases and stress-activated protein kinases. J. Biochem. Mol. Biol. 2002, 35, 24–27. [Google Scholar] [CrossRef]
- Bruckheimer, E.M.; Cho, S.H.; Sarkiss, M.; Herrmann, J.; McDonnell, T.J. The Bcl-2 gene family and apoptosis. Adv. Biochem. Eng. Biotechnol. 1998, 62, 75–105. [Google Scholar] [CrossRef]
- Cleland, M.M.; Norris, K.L.; Karbowski, M.; Wang, C.; Suen, D.F.; Jiao, S.; George, N.M.; Luo, X.; Li, Z.; Youle, R.J. Bcl-2 family interaction with the mitochondrial morphogenesis machinery. Cell Death Differ. 2011, 18, 235–247. [Google Scholar] [CrossRef]
- Noor, M.R.; Soulimane, T. Structure of caa(3) cytochrome c oxidase--a nature-made enzyme-substrate complex. Biol. Chem. 2013, 394, 579–591. [Google Scholar] [CrossRef]
- Hu, Y.; Benedict, M.A.; Ding, L.; Nunez, G. Role of cytochrome c and dATP/ATP hydrolysis in Apaf-1-mediated caspase-9 activation and apoptosis. EMBO J. 1999, 18, 3586–3595. [Google Scholar] [CrossRef]
- Zhou, M.; Li, Y.; Hu, Q.; Bai, X.C.; Huang, W.; Yan, C.; Scheres, S.H.; Shi, Y. Atomic structure of the apoptosome: Mechanism of cytochrome c- and dATP-mediated activation of Apaf-1. Genes. Dev. 2015, 29, 2349–2361. [Google Scholar] [CrossRef]
- Graumann, K.; Schaumburg, F.; Reubold, T.F.; Hippe, D.; Eschenburg, S.; Lüder, C.G. Toxoplasma gondii inhibits cytochrome c-induced caspase activation in its host cell by interference with holo-apoptosome assembly. Microb. Cell 2015, 2, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Hoesel, B.; Schmid, J.A. The complexity of NF-kappaB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [PubMed]
- Miri, S.; Rasooli, A.; Brar, S.K. Data on changes of NF-κB gene expression in liver and lungs as a biomarker and hepatic injury in CLP-induced septic rats. Data Brief. 2019, 25, 104117. [Google Scholar] [CrossRef] [PubMed]
- Herman, R.K.; Molestina, R.E.; Sinai, A.P.; Howe, D.K. The apicomplexan pathogen Neospora caninum inhibits host cell apoptosis in the absence of discernible NF-kappa B activation. Infect. Immun. 2007, 75, 4255–4262. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.B.; Park, J.H.; Kang, S.S.; Kang, D.H.; Lee, J.H.; Oh, S.J.; Lee, J.Y.; Kim, J.Y.; Tchah, H. Protective Effects of Cyclosporine A Emulsion Versus Cyclosporine A Cationic Emulsion Against Desiccation Stress in Human Corneal Epithelial Cells. Cornea 2020, 39, 508–513. [Google Scholar] [CrossRef]
- Wang, H.; Zhu, Y.; Xu, X.; Wang, X.; Hou, Q.; Xu, Q.; Sun, Z.; Mi, Y.; Hu, C. Ctenopharyngodon idella NF-kappaB subunit p65 modulates the transcription of IkappaBalpha in CIK cells. Fish. Shellfish. Immunol. 2016, 54, 564–572. [Google Scholar] [CrossRef]
- Fruman, D.A.; Chiu, H.; Hopkins, B.D.; Bagrodia, S.; Cantley, L.C.; Abraham, R.T. The PI3K Pathway in Human Disease. Cell 2017, 170, 605–635. [Google Scholar] [CrossRef]
- Xu, J.; Wang, K.; Zhang, Z.; Xue, D.; Li, W.; Pan, Z. The Role of Forkhead Box Family in Bone Metabolism and Diseases. Front. Pharmacol. 2021, 12, 772237. [Google Scholar] [CrossRef]
- Prudêncio, M.; Rodriguez, A.; Mota, M.M. The silent path to thousands of merozoites: The Plasmodium liver stage. Nat. Rev. Microbiol. 2006, 4, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.H.; Jeffers, M.; Bellacosa, A.; Mitsuuchi, Y.; Vande Woude, G.F.; Testa, J.R. Anti-apoptotic signaling by hepatocyte growth factor/Met via the phosphatidylinositol 3-kinase/Akt and mitogen-activated protein kinase pathways. Proc. Natl. Acad. Sci. USA 2001, 98, 247–252. [Google Scholar] [CrossRef]
- Sturm, A.; Amino, R.; van de Sand, C.; Regen, T.; Retzlaff, S.; Rennenberg, A.; Krueger, A.; Pollok, J.M.; Menard, R.; Heussler, V.T. Manipulation of host hepatocytes by the malaria parasite for delivery into liver sinusoids. Science 2006, 313, 1287–1290. [Google Scholar] [CrossRef]


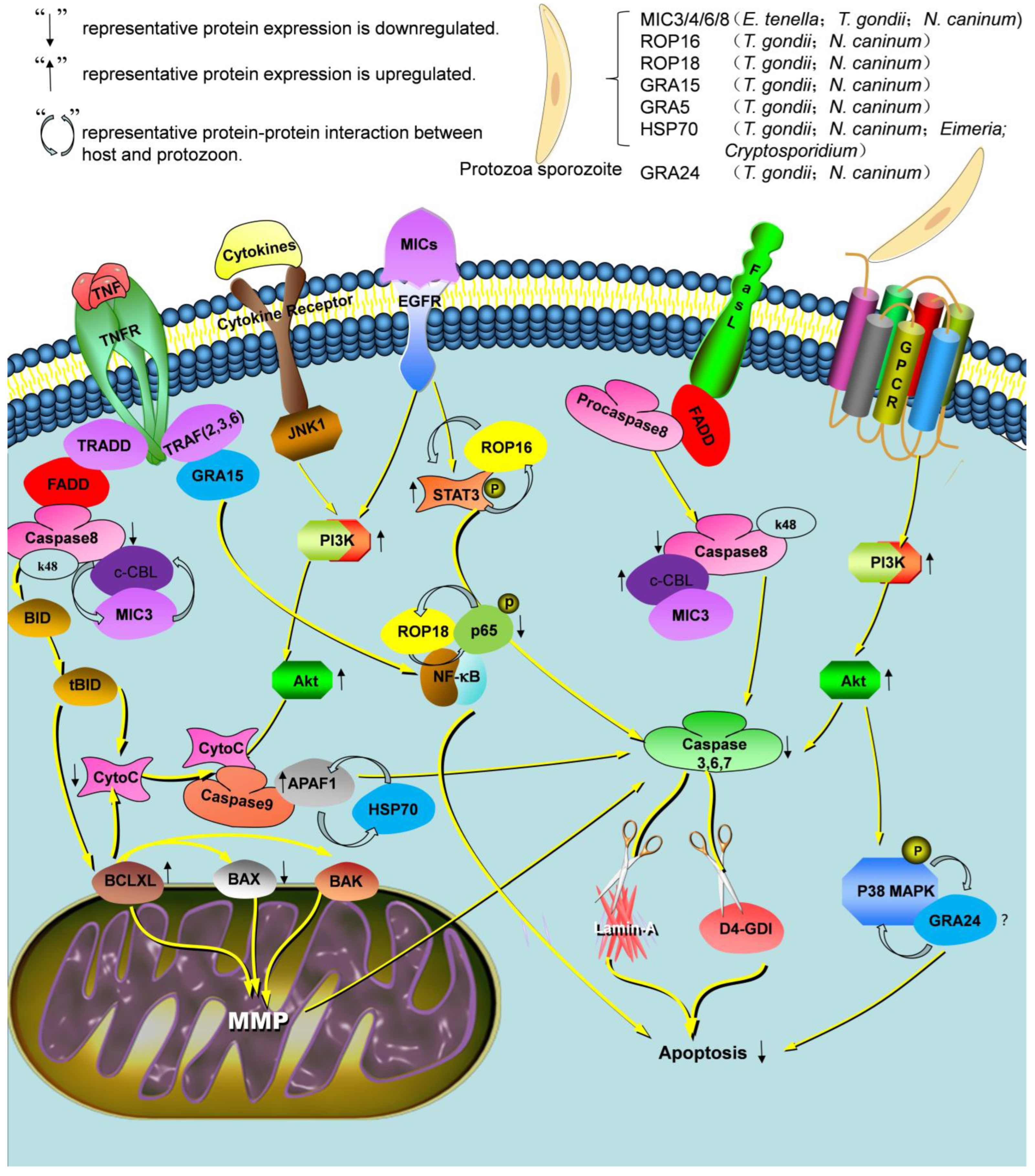{kind=link}
| Signaling Pathway | Apicomplexan Protozoa | References | |
|---|---|---|---|
| MIC6 | EGFR/PI3K/Akt | T. gondii | Muniz-Feliciano L et al. (2013) [30] |
| ROP16 | JKT-STAT3 | T. gondii | Yamamoto M et al. (2009) [33] |
| ROP18 | NF-κB | T. gondii | Du J et al. (2014) [35] |
| GRA15 | NF-κB | T. gondii | Sangaré LO et al. (2019) [43] |
| HSP70 | Cytochrome-C | T. gondii | Mosser D et al. (2000) [48] |
| HSP70 | APAF-1 | T. gondii | Saleh A et al. (2000) [49] |
| MIC3 | EGFR/PI3K/Akt | Eimeria | Muniz-Feliciano L et al. (2013) [30] |
| MIC3 | CBL | Eimeria | Wang P et al. (2021) [6] |
| GRA5 | Unknown | T. gondii | Ching, X.T et al. (2015) [40] |
| MIC8 | EGFR/PI3K/Akt | T. gondii | Meissner M et al. (2002) [31] |
| MIC4 | EGFR/PI3K/Akt | Eimeria | Xuesong Zhang et al. (2022) [25] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lian, L.; Sun, Q.; Huang, X.; Li, W.; Cui, Y.; Pan, Y.; Yang, X.; Wang, P. Inhibition of Cell Apoptosis by Apicomplexan Protozoa–Host Interaction in the Early Stage of Infection. Animals 2023, 13, 3817. https://doi.org/10.3390/ani13243817
Lian L, Sun Q, Huang X, Li W, Cui Y, Pan Y, Yang X, Wang P. Inhibition of Cell Apoptosis by Apicomplexan Protozoa–Host Interaction in the Early Stage of Infection. Animals. 2023; 13(24):3817. https://doi.org/10.3390/ani13243817
Chicago/Turabian StyleLian, Liyin, Qian Sun, Xinyi Huang, Wanjing Li, Yanjun Cui, Yuebo Pan, Xianyu Yang, and Pu Wang. 2023. "Inhibition of Cell Apoptosis by Apicomplexan Protozoa–Host Interaction in the Early Stage of Infection" Animals 13, no. 24: 3817. https://doi.org/10.3390/ani13243817
APA StyleLian, L., Sun, Q., Huang, X., Li, W., Cui, Y., Pan, Y., Yang, X., & Wang, P. (2023). Inhibition of Cell Apoptosis by Apicomplexan Protozoa–Host Interaction in the Early Stage of Infection. Animals, 13(24), 3817. https://doi.org/10.3390/ani13243817