
Comparison of Uterine Involution and the Resumption of Ovarian Cyclicity between Lame and Sound Holstein Cows
 ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animals and Housing
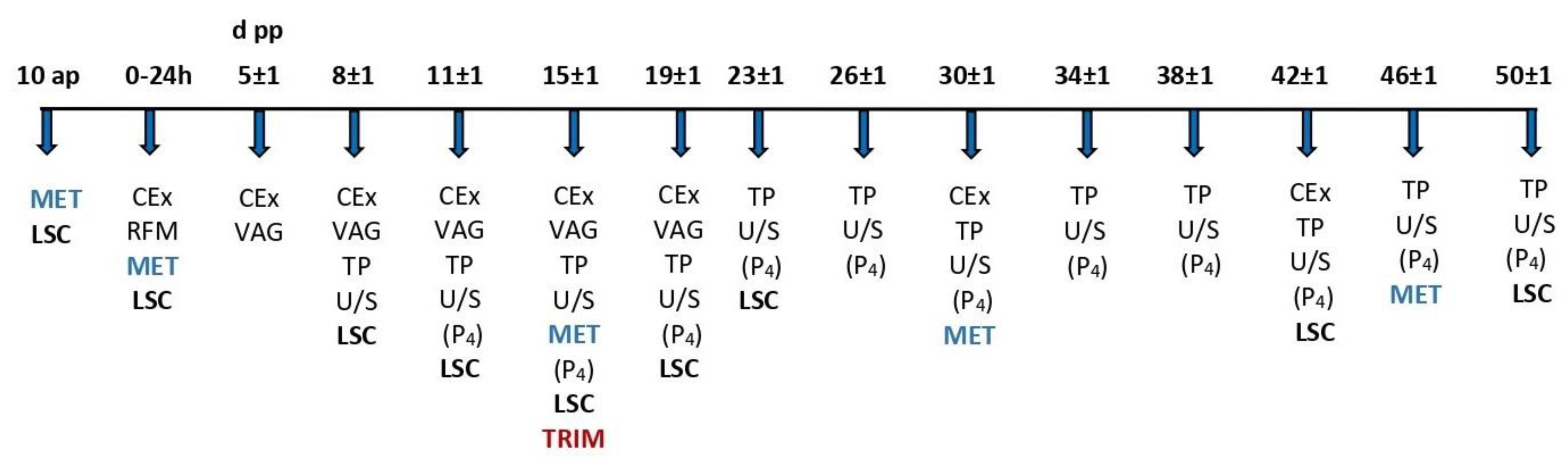
2.3. Inclusion Criteria and Experimental Procedures
- Day 10 ap: Blood sampling for the detection of BHBA and NEFA concentrations and lameness scoring (LSC).
- Calving day and within 24 h: Blood sampling for the detection of BHBA and NEFA concentrations, clinical examination, examination for retained fetal membranes, and lameness scoring.
- Day 5 ± 1 pp: Clinical examination, transrectal palpation, and vaginoscopy for the assessment of the genital system and uterine discharge.
- Day 8 ± 1 to 50 ± 1 pp: Clinical examination, lameness scoring, vaginoscopy, transrectal palpation, and ultrasound examination to record ovarian formations and the involution of the cervix and the formerly gravid uterine horn at the respective time points. In the case of the detection of an ovarian follicle larger than 10 mm, ultrasonography and blood sampling for the detection of progesterone concentration were repeated every 24–48 h until ovulation, atresia and regression, or cyst formation was confirmed. On day 15 ± 1, examination and trimming of all claws were performed on a standing chute, and lesions were recorded. All samplings, measurements, and treatments were performed by the first author.
2.4. Lameness Assessment and Recording
2.5. Genital Examination and Ultrasonography
2.6. Reproductive Definitions and Outcomes
- Cows with normal ovarian activity (cyclic) on day 50 pp: ovulation followed by the emergence of a CL and a new ovulation within an interval of ≤24 days (n = 33/47, 70.2%) [22,32]. In the case of a single ovulation in less than 24 days before the end of the experiment, its duration was extended for proper classification.
- Cows with an ovarian cyst on day 50 pp (n = 8/47, 17.0%).
- Cows with atresia on day 50 pp (n = 3/47, 6.4%).
- Cows with a prolonged luteal phase on day 50 pp (n = 3/47, 6.4%).
- Ovulation within the experimental period (n = 36/47, 76.6%).
- The day of the first ovulation.
- Formation of an ovarian cyst within the experimental period (n = 17/47, 36.2%).
- Prolonged luteal phase within the experimental period (n = 5/47, 10.6%).
2.7. Blood Sampling and Analytic Assays
- At calving: 0.9 mEq/L
- On day 15 pp: 0.6 mEq/L
- Antepartum and after day 15 pp: 0.4 mEq/L
2.8. Statistical Analysis
3. Results
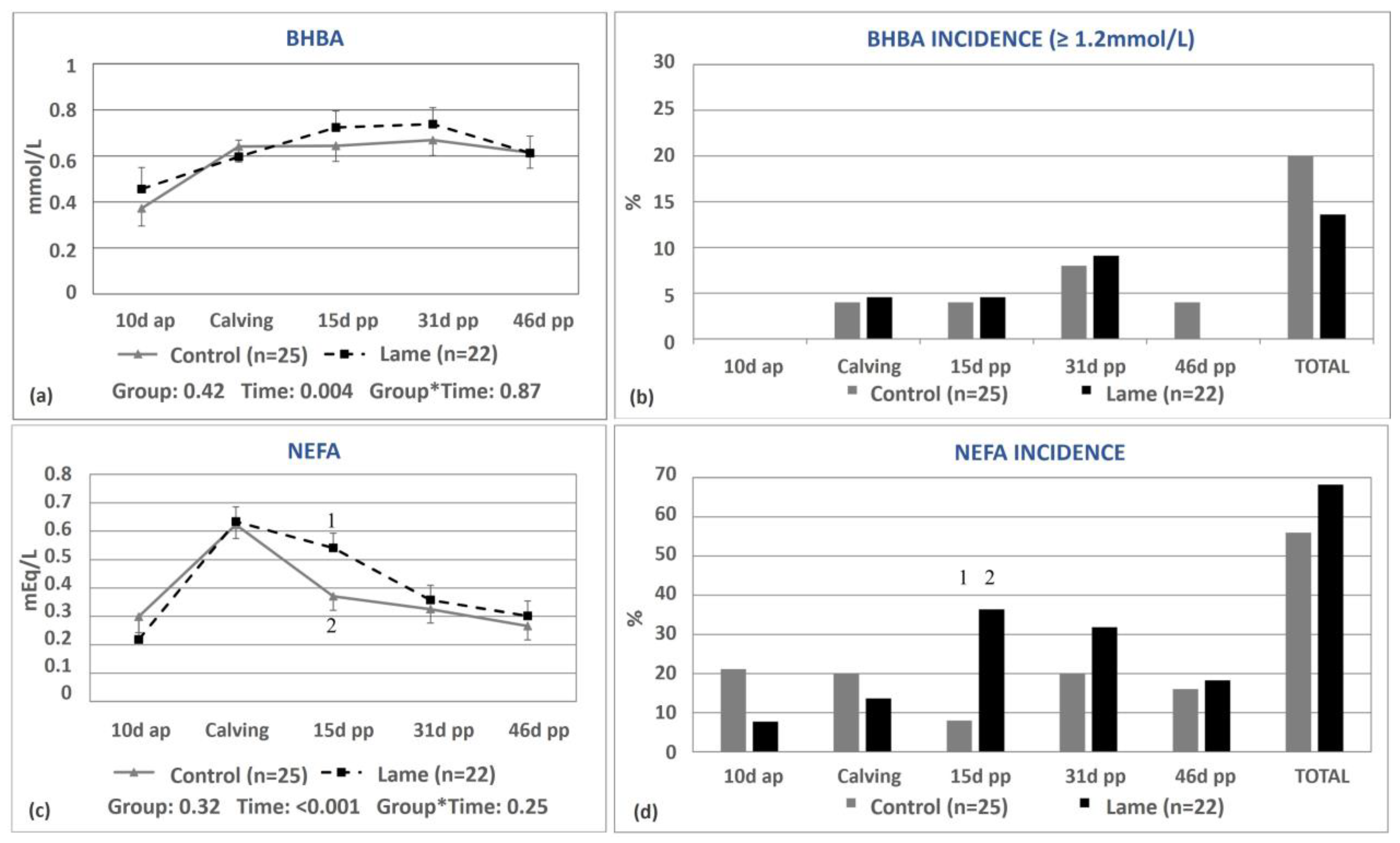
3.1. Energy Status of the Lame Cows
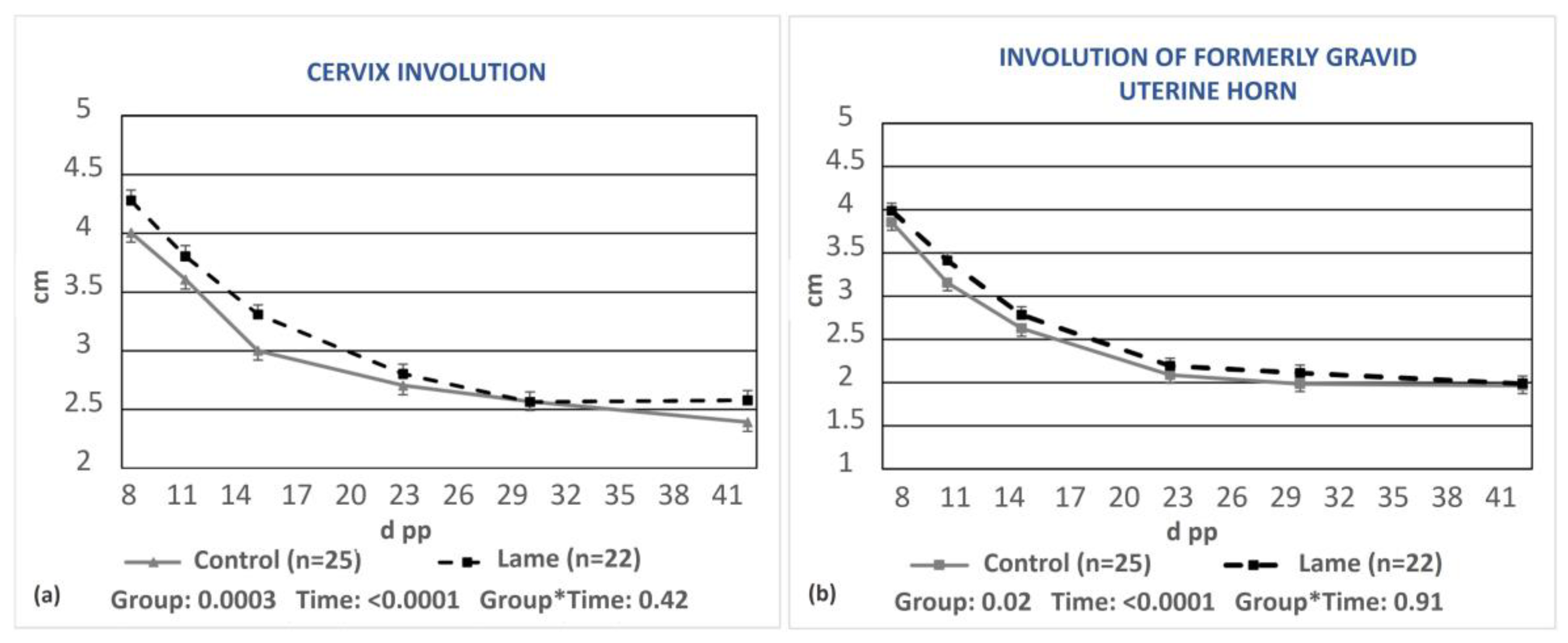
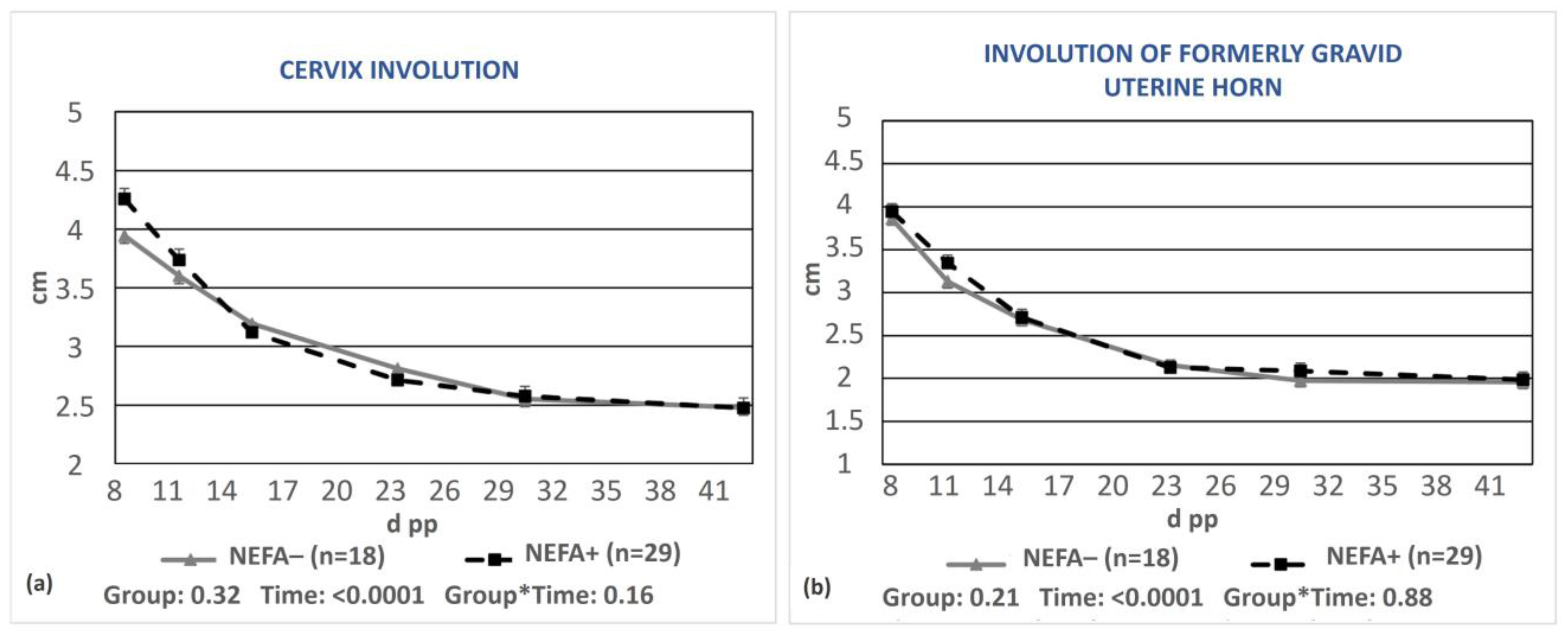
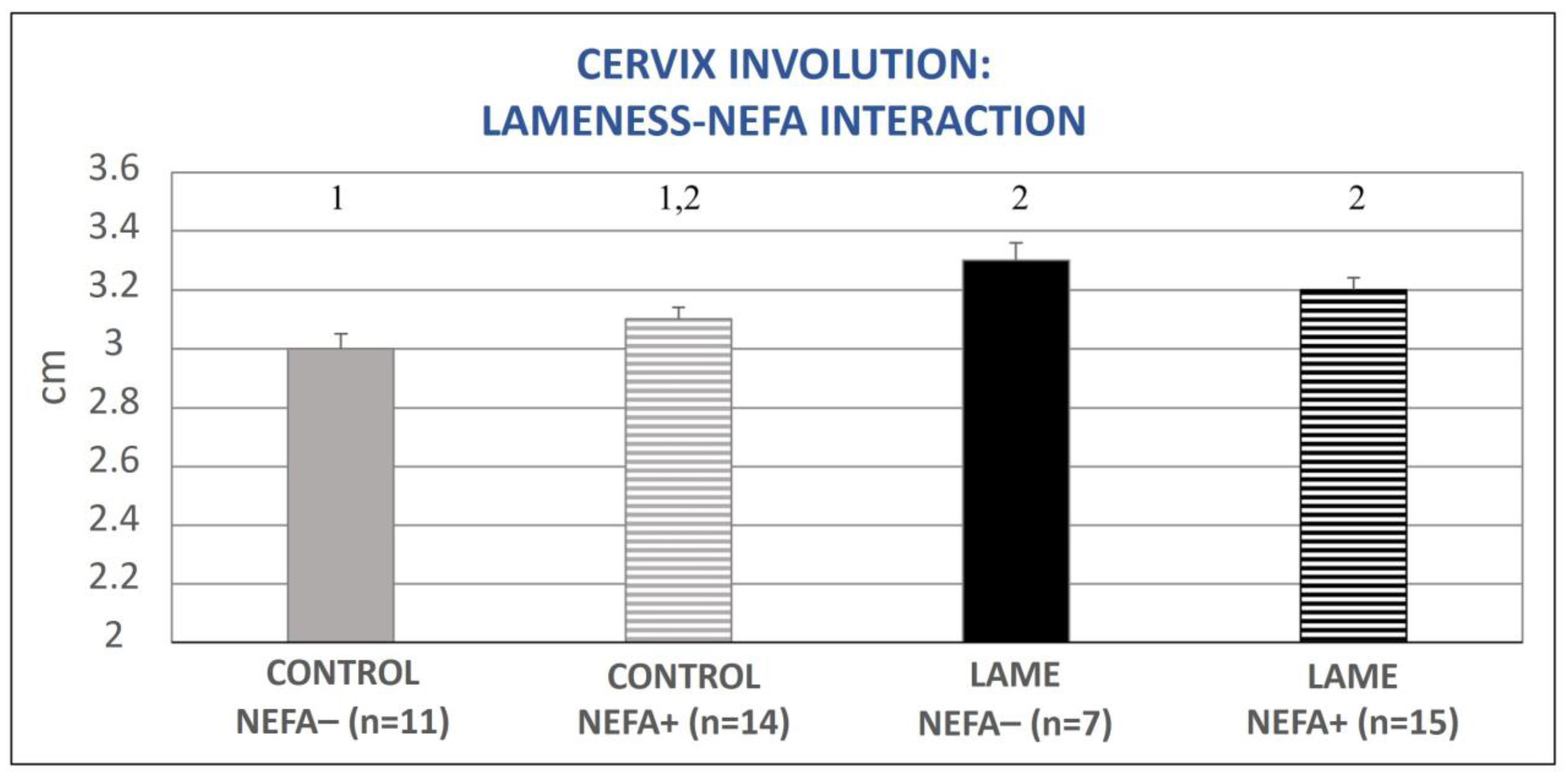
3.2. Uterine Involution in Relation to Lameness and Energy Status
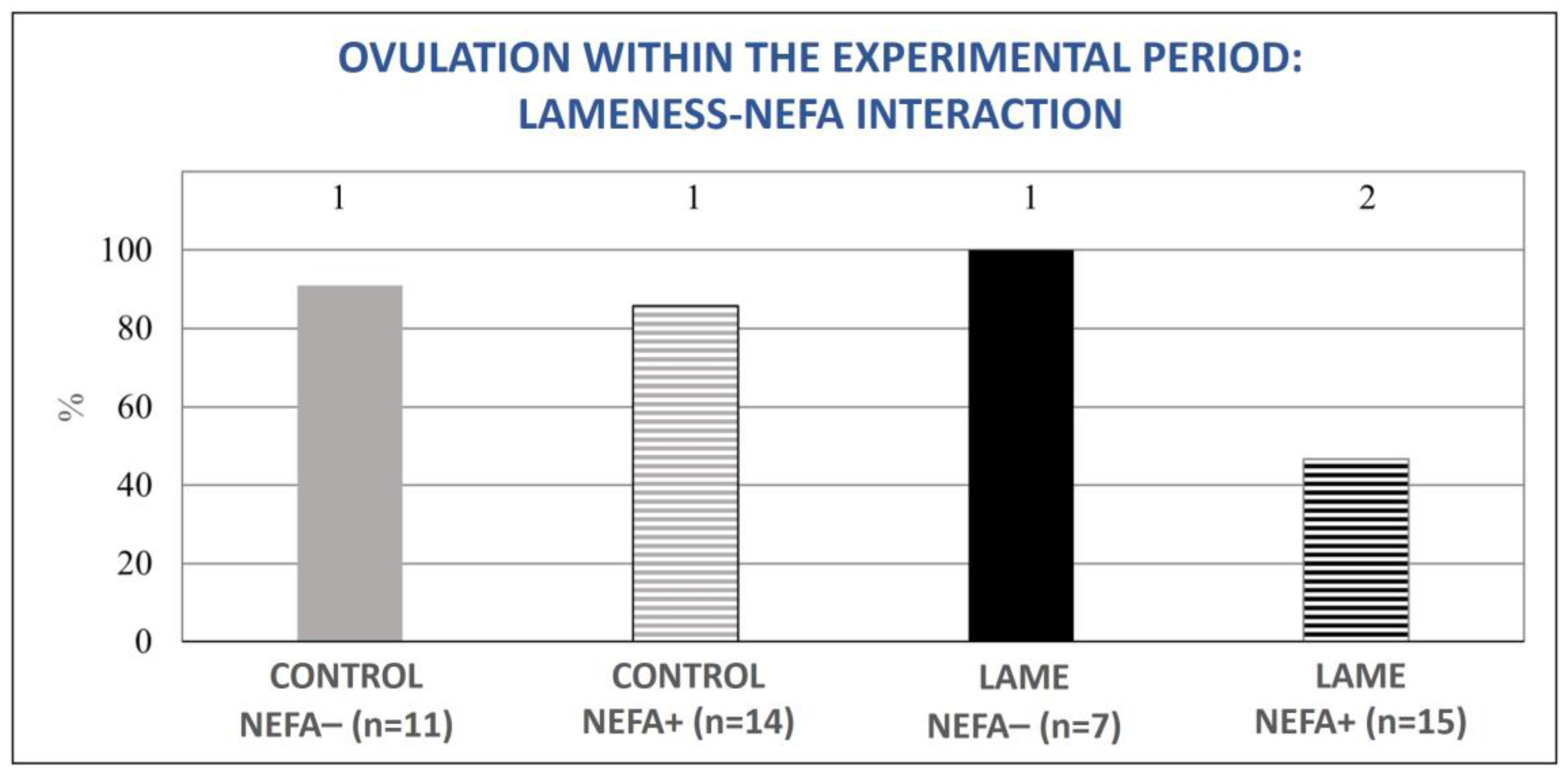
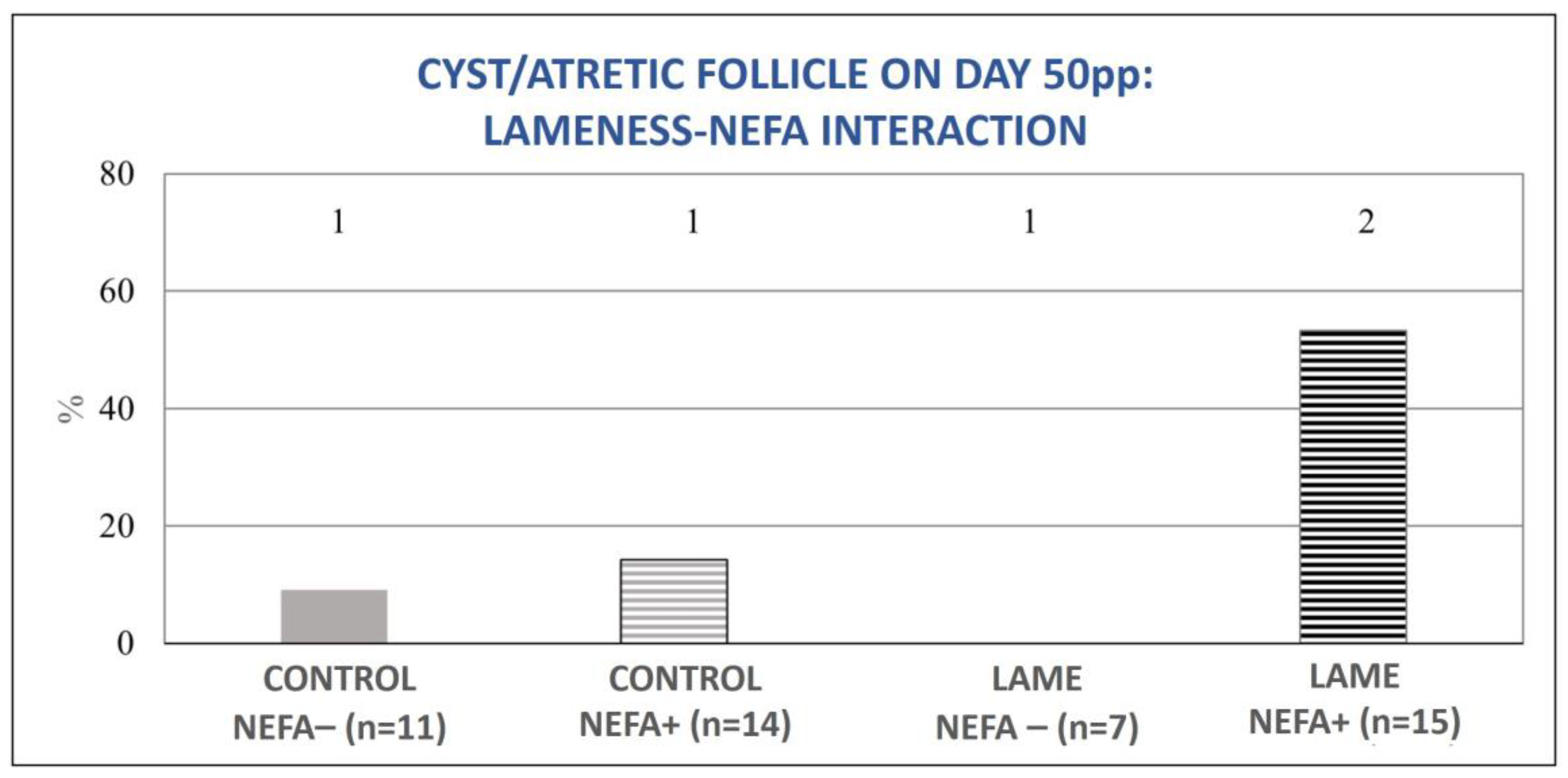
3.3. Resumption of Ovarian Activity in Relation to Lameness and Energy Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fourichon, C.; Seegers, H.; Malher, X. Effect of disease on reproduction in the dairy cow: A meta-analysis. Theriogenology 2000, 53, 1729–1759. [Google Scholar] [CrossRef] [PubMed]
- Machado, V.S.; Caixeta, L.S.; McArt, J.A.A.; Bicalho, R.C. The effect of claw horn disruption lesions and body condition score at dry-off on survivability, reproductive performance, and milk production in the subsequent lactation. J. Dairy Sci. 2010, 93, 4071–4078. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.J.; Kaneko, K.; Walker, S.L.; Jones, D.N.; Routly, J.E.; Smith, R.F.; Dobson, H. Influence of lameness on follicular growth, ovulation, reproductive hormone concentrations and estrus behavior in dairy cows. Theriogenology 2011, 76, 658–668. [Google Scholar] [CrossRef]
- Tsousis, G.; Boscos, C.; Praxitelous, A. The negative impact of lameness on dairy cow reproduction. Reprod. Domest. Anim. 2022, 57, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Okano, A.; Tomizuka, T. Postpartum uterine involution in the cow. Japan Agric. Res. Quart. 1996, 30, 113–121. [Google Scholar]
- Mateus, L.; Lopes da Costa, L.; Bernardo, F.; Robalo Silva, J. Influence of Puerperal Uterine Infection on Uterine Involution and Postpartum Ovarian Activity in Dairy Cows. Reprod. Domest. Anim. 2002, 37, 31–35. [Google Scholar] [CrossRef]
- Kindahl, H.; Odensvik, K.; Aiumlamai, S.; Fredriksson, G. Utero-ovarian relationships during the bovine postpartum period. Anim. Reprod. Sci. 1992, 28, 363–369. [Google Scholar] [CrossRef]
- Lewis, G.S. Uterine Health and Disorders. J. Dairy Sci. 1997, 80, 984–994. [Google Scholar] [CrossRef]
- Sheldon, I.M. The postpartum uterus. Vet. Clin. N. Am. Food Anim. 2004, 20, 569–591. [Google Scholar] [CrossRef]
- Zhang, J.; Deng, L.X.; Zhang, H.L.; Hua, G.H.; Han, L.; Zhu, Y.; Meng, X.J.; Yang, L.G. Effects of parity on uterine involution and resumption of ovarian activities in postpartum Chinese Holstein dairy cows. J. Dairy Sci. 2010, 93, 1979–1986. [Google Scholar] [CrossRef]
- Heppelmann, M.; Weinert, M.; Brömmling, A.; Piechotta, M.; Hoedemaker, M.; Bollwein, H. The effect of puerperal uterine disease on uterine involution in cows assessed by Doppler sonography of the uterine arteries. Anim. Reprod. Sci. 2013, 143, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Yang, H.; Ahmad, M.J.; Yang, Y.; Yang, W.; Riaz, H.; Abulaiti, A.; Zhang, S.; Yang, L.; Hua, G. Postpartum uterine involution and embryonic development pattern in chinese holstein dairy cows. Front. Vet. Sci. 2020, 7, 604729. [Google Scholar] [CrossRef] [PubMed]
- Genís, S.; Arís, A.; Kaur, M.; Cerri, R.L.A. Effect of metritis on endometrium tissue transcriptome during puerperium in Holstein lactating cows. Theriogenology 2018, 122, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Paiano, B.R.; Birgel, B.D.; Birgel, H.E. Uterine involution and reproductive performance in dairy cows with metabolic diseases. Animals 2019, 3, 93. [Google Scholar] [CrossRef] [PubMed]
- Daros, R.R.; Eriksson, H.K.; Weary, D.M.; von Keyserlingk, M.A.G. The relationship between transition period diseases and lameness, feeding time, and body condition during the dry period. J. Dairy Sci. 2020, 103, 649–665. [Google Scholar] [CrossRef]
- Sogstad, A.M.; Osteras, O.; Fjeldaas, T. Bovine claw and limb disorders related to reproductive performance and production diseases. J. Dairy Sci. 2006, 89, 2519–2528. [Google Scholar] [CrossRef] [PubMed]
- Calderon, D.F.; Cook, N.B. The effect of lameness on the resting behavior and metabolic status of dairy cattle during the transition period in a freestall-housed dairy herd. J. Dairy Sci. 2011, 94, 2883–2894. [Google Scholar] [CrossRef]
- Garbarino, E.J.; Hernandez, J.A.; Shearer, J.K.; Risco, C.A.; Thatcher, W.W. Effect of lameness on ovarian activity in postpartum Holstein cows. J. Dairy Sci. 2004, 87, 4123–4131. [Google Scholar] [CrossRef]
- Melendez, P.; Gomez, V.; Bothe, H.; Rodriguez, F.; Velez, J.; Lopez, H.; Bartolome, J.; Archbald, L. Ultrasonographic ovarian dynamic, plasma progesterone, and non-esterified fatty acids in lame postpartum dairy cows. J. Vet. Sci. 2018, 19, 462–467. [Google Scholar] [CrossRef]
- Opsomer, G.; Gröhn, Y.T.; Hertl, J.; Coryn, M.; Deluyker, H.; de Kruif, A. Risk factors for post partum ovarian dysfunction in high producing dairy cows in Belgium: A field study. Theriogenology 2000, 53, 841–857. [Google Scholar] [CrossRef]
- Gillund, P.; Reksen, O.; Gröhn, Y.T.; Karlberg, K. Body condition related to ketosis and reproductive performance in Norwegian dairy cows. J. Dairy Sci. 2001, 84, 1390–1396. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, H.K.; Nakao, T.; Higaki, T.; Suzuki, T.; Akita, M.J.T. Resumption of postpartum ovarian cyclicity in high-producing Holstein cows. Theriogenology 2004, 61, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Walsh, R.B.; Kelton, D.F.; Duffield, T.F.; Leslie, K.E.; Walton, J.S.; LeBlanc, S.J. Prevalence and risk factors for postpartum anovulatory condition in dairy cows. J. Dairy Sci. 2007, 90, 315–324. [Google Scholar] [CrossRef]
- Beam, S.W.; Butler, W.R. Effects of energy balance on follicular development and first ovulation in postpartum. Reprod. Fertil. Suppl. 1999, 54, 411–424. [Google Scholar] [CrossRef]
- Diskin, M.; Mackey, D.; Roche, J.; Sreenan, J. Effects of nutrition and metabolic status on circulating hormones and ovarian follicle development in cattle. Anim. Reprod. Sci. 2003, 78, 345–370. [Google Scholar] [CrossRef] [PubMed]
- Chagas, L.; Bass, J.; Blache, D.; Burke, C.; Kay, J.; Lindsay, D.; Lucy, M.; Martin, G.; Meier, S.; Rhodes, F. Invited review: New perspectives on the roles of nutrition and metabolic priorities in the subfertility of high-producing dairy cows. J. Dairy Sci. 2007, 90, 4022–4032. [Google Scholar] [CrossRef] [PubMed]
- Galindo, F.; Broom, D.M. The effects of lameness on social and individual behavior of dairy cows. J. Appl. Anim. Welf. Sci. 2002, 5, 193–201. [Google Scholar] [CrossRef]
- Bach, A.; Dinarés, M.; Devant, M.; Carré, X. Associations between lameness and production, feeding and milking attendance of Holstein cows milked with an automatic milking system. J. Dairy Res. 2007, 74, 40–46. [Google Scholar] [CrossRef]
- Hoedemaker, M.; Prange, D.; Gundelach, Y. Body condition change ante- and postpartum, health and reproductive performance in German Holstein cows. Reprod. Domest. Anim. 2009, 44, 167–173. [Google Scholar] [CrossRef]
- Randall, L.V.; Green, M.J.; Chagunda, M.G.; Mason, C.; Archer, S.C.; Green, L.E.; Huxley, J.N. Low body condition predisposes cattle to lameness: An 8-year study of one dairy herd. J. Dairy Sci. 2015, 98, 3766–3777. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Dairy Cattle, 7th ed.; The National Academies Press: Washington, DC, USA, 2001; pp. 1–333. [Google Scholar] [CrossRef]
- Tanaka, T.; Arai, M.; Ohtani, S.; Uemura, S.; Kuroiwa, T.; Kim, S.; Kamomae, H. Influence of parity on follicular dynamics and resumption of ovarian cycle in postpartum dairy cows. Anim. Reprod. Sci. 2008, 108, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Van Tassell, C.P.; Wiggans, G.R.; Misztal, I. Implementation of a sire-maternal grandsire model for evaluation of calving ease in the United States. J. Dairy Sci. 2003, 86, 3366–3373. [Google Scholar] [CrossRef] [PubMed]
- Sprecher, D.J.; Hostetler, D.E.; Kaneene, J.B. A lameness scoring system that uses posture and gait to predict dairy cattle reproductive performance. Theriogenology 1997, 47, 1179–1187. [Google Scholar] [CrossRef]
- Greenough, P.R. Bovine Laminitis and Lameness; W.B. Saunders: Edinburgh, UK, 2007; pp. 29–240. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Cronin, J.; Goetze, L.; Donofrio, G.; Schuberth, H.J. Defining postpartum uterine disease and the mechanisms of infection and immunity in the female reproductive tract in cattle. Biol. Reprod. 2009, 81, 1025–1032. [Google Scholar] [CrossRef]
- McDougall, S.; Burke, C.R.; Macmillan, K.L.; Williamson, N.B. Patterns of follicular development during periods of anovulation in pasture-fed dairy cows after calving. Res. Vet. Sci. 1995, 58, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Peter, A.T.; Vos, P.L.A.M.; Ambrose, D.J. Postpartum anestrus in dairy cattle. Theriogenology 2009, 71, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Vanholder, T.; Opsomer, G.; de Kruif, A. Aetiology and pathogenesis of cystic ovarian follicles in dairy cattle: A review. Reprod. Nutr. Dev. 2006, 46, 105–119. [Google Scholar] [CrossRef]
- Leblanc, S. Monitoring Metabolic Health of Dairy Cattle in the Transition Period. J. Reprod. Dev. 2010, 56, S29–S35. [Google Scholar] [CrossRef]
- Ospina, P.A.; Nydam, D.V.; Stokol, T.; Overton, T.R. Evaluation of nonesterified fatty acids and β-hydroxybutyrate in transition dairy cattle in the northeastern United States: Critical thresholds for prediction of clinical diseases. J. Dairy Sci. 2010, 93, 546–554. [Google Scholar] [CrossRef]
- Dubuc, J.; Duffield, T.F.; Leslie, K.E.; Walton, J.S.; LeBlanc, S.J. Risk factors and effects of postpartum anovulation in dairy cows. J. Dairy Sci. 2012, 95, 1845–1854. [Google Scholar] [CrossRef]
- Oltenacu, P.A.; Britt, J.H.; Braun, R.K.; Mellenberger, R.W. Relationships among type of parturition, type of discharge from genital tract, involution of cervix, and subsequent reproductive performance in Holstein cows. J. Dairy Sci. 1983, 66, 612–619. [Google Scholar] [CrossRef]
- Yáñez, U.; Herradón, P.G.; Becerra, J.J.; Peña, A.I.; Quintela, L.A. Relationship between postpartum metabolic status and subclinical endometritis in dairy cattle. Animals 2022, 12, 242. [Google Scholar] [CrossRef] [PubMed]
- Praxitelous, A.; Katsoulos, P.D.; Tsaousioti, A.; Brozos, C.; Theodosiadou, E.K.; Boscos, C.M.; Tsousis, G. Ovarian and energy status in lame dairy cows at puerperium and their responsiveness in protocols for the synchronization of ovulation. Animals 2023, 13, 1537. [Google Scholar] [CrossRef] [PubMed]
- Sood, P.; Nanda, A.S.; Singh, N.; Javed, M.; Prasad, R. Effect of lameness on follicular dynamics in crossbred cows. Vet. Arh. 2009, 79, 131–141. Available online: https://hrcak.srce.hr/37319 (accessed on 20 October 2023).
- Melendez, P.; Bartolome, J.; Archbald, L.F.; Donovan, A. The association between lameness, ovarian cysts and fertility in lactating dairy cows. Theriogenology 2003, 59, 927–937. [Google Scholar] [CrossRef]
- Sun, D.; Li, C.; Gu, C.; Chen, J.; Qu, Y.; Wang, X.; Gao, J.; Wei, S.; Wang, J.; Wu, R. Analysis of mineral elements, metabolism, and inflammation indexes in the plasma of dairy cows suffering from different degrees of lameness. Biol. Trace Elem. Res. 2015, 168, 372–379. [Google Scholar] [CrossRef]
- Mudroň, P. Role of ketosis in lame dairy cows. Bulg. J. Vet. Med. 2019, 22 (Suppl. S1), 70–73. [Google Scholar]
- Kougioumtzis, A. Genetic and Epidemiological Study of Lameness in Dairy Cattle. Ph.D. Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 2014. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | ||
|---|---|---|
| Variable | Control | Lame |
| n | 25 | 22 |
| Ovulation between days 0 and 50 pp (%) | 88.0 1 | 63.6 2 |
| Day of 1st ovulation (mean ± SE) | 27.3 ± 2.8 | 29.3 ± 3.5 |
| Ovarian cyst between days 0 and 50 pp (%) | 36.0 | 36.4 |
| Prolonged luteal phase between days 0 and 50 pp (%) | 12.0 | 4.6 |
| Normal ovarian cyclicity on day 50 pp (%) | 80.0 | 59.1 |
| Ovarian cyst on day 50 pp (%) | 12.0 | 22.7 |
| Atretic follicle on day 50 pp (%) | 0.0 | 13.6 |
| Ovarian cyst or atretic follicle on day 50 pp (%) | 12.0 1 | 36.4 2 |
| Prolonged luteal phase on day 50 pp (%) | 8.0 | 4.6 |
| Group | ||
|---|---|---|
| Variable | BHBA− | BHBA+ |
| n | 39 | 8 |
| Ovulation between days 0 and 50 pp (%) | 79.5 | 62.5 |
| Day of 1st ovulation (mean ± SE) | 28.3 ± 2.4 | 26.8 ± 5.6 |
| Ovarian cyst between days 0 and 50 pp (%) | 33.3 | 50.0 |
| Prolonged luteal phase between days 0 and 50 pp (%) | 7.7 | 12.5 |
| Normal ovarian cyclicity on day 50 pp (%) | 74.4 | 50.0 |
| Ovarian cyst on day 50 pp (%) | 18.0 | 12.5 |
| Atretic follicle on day 50 pp (%) | 2.6 | 25.0 |
| Ovarian cyst or atretic follicle on day 50 pp (%) | 20.5 | 37.5 |
| Prolonged luteal phase on day 50 pp (%) | 5.1 | 12.5 |
| Group | ||
|---|---|---|
| Variable | NEFA− | NEFA+ |
| n | 18 | 29 |
| Ovulation between days 0 and 50 pp (%) | 94.4 1 | 65.5 2 |
| Day of 1st ovulation (mean ± SE) | 28.6 ± 3.3 | 27.5 ± 2.9 |
| Ovarian cyst between days 0 and 50 pp (%) | 27.8 | 41.4 |
| Prolonged luteal phase between days 0 and 50 pp (%) | 11.1 | 6.7 |
| Normal ovarian cyclicity on day 50 pp (%) | 88.9 1 | 58.6 2 |
| Ovarian cyst on day 50 pp (%) | 5.6 | 24.1 |
| Atretic follicle on day 50 pp (%) | 0 | 10.3 |
| Ovarian cyst or atretic follicle on day 50 pp (%) | 5.6 1 | 34.5 2 |
| Prolonged luteal phase on day 50 pp (%) | 5.6 | 6.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Praxitelous, A.; Katsoulos, P.D.; Tsaousioti, A.; Brozos, C.; Schmicke, M.; Boscos, C.M.; Tsousis, G. Comparison of Uterine Involution and the Resumption of Ovarian Cyclicity between Lame and Sound Holstein Cows. Animals 2023, 13, 3645. https://doi.org/10.3390/ani13233645
Praxitelous A, Katsoulos PD, Tsaousioti A, Brozos C, Schmicke M, Boscos CM, Tsousis G. Comparison of Uterine Involution and the Resumption of Ovarian Cyclicity between Lame and Sound Holstein Cows. Animals. 2023; 13(23):3645. https://doi.org/10.3390/ani13233645
Chicago/Turabian StylePraxitelous, Anastasia, Panagiotis D. Katsoulos, Angeliki Tsaousioti, Christos Brozos, Marion Schmicke, Constantin M. Boscos, and Georgios Tsousis. 2023. "Comparison of Uterine Involution and the Resumption of Ovarian Cyclicity between Lame and Sound Holstein Cows" Animals 13, no. 23: 3645. https://doi.org/10.3390/ani13233645
APA StylePraxitelous, A., Katsoulos, P. D., Tsaousioti, A., Brozos, C., Schmicke, M., Boscos, C. M., & Tsousis, G. (2023). Comparison of Uterine Involution and the Resumption of Ovarian Cyclicity between Lame and Sound Holstein Cows. Animals, 13(23), 3645. https://doi.org/10.3390/ani13233645