1. Introduction
Rutin is a collection of natural compounds that possess a distinctive polyphenolic structure of flavonoids [
1]. It exhibits an abundance of phenolic hydroxyl groups, particularly catechol functional groups, which facilitate easy absorption in the small intestine or binding with endogenous proteins within the intestinal tract. Consequently, it could reach intestinal tissue and be transported to other tissues via the bloodstream. Rutin has demonstrated various biological activities including anti-inflammatory effects [
2], antioxidant properties [
3], antiviral capabilities [
4], as well as vasoactive and neuroprotective functions [
5]. These remarkable attributes hold potential for applications aimed at reducing oxidative stress, alleviating inflammation, promoting growth, and enhancing immunity in organisms. In addition, its safety, low toxicity, and lack of biological residues make it a promising candidate for the development of functional feed additives in aquaculture. For example, supplementation with 0.8 and 1.0 g/kg of rutin in the diet can improve the growth performance and muscle quality of grass carp (
Ctenopharyngodon idella) [
6]. The administration of 1.5 g/kg of rutin in the diet of silver catfish (
Rhamdia quelen) resulted in increased antioxidant enzyme activities in the brain, liver, kidney, and gills while also promoting growth [
7]. Furthermore, dietary supplementation with rutin can improve the liver antioxidant response of Nile tilapia and reduce liver and muscle fiber damage induced by T-2 toxin, thereby improving the health status of Nile tilapia (
Oreochromis niloticus) [
8].
Yellow catfish (
Pelteobagrus fulvidraco) is a widely cultured fish species in China. Its production reached about 0.58 million tons, which ranked tenth in total freshwater fish output [
9]. Currently, fish cultivation predominantly relies on high-density ponds. However, this intensive culture pattern inevitably leads to metabolic disorders in fish and deterioration of water quality. With the increasing market demands and expansion of large-scale farming, yellow catfish have been found to exhibit a range of issues including growth retardation, compromised antioxidant capacity, and weakened disease resistance [
10]. Consequently, ensuring the health and enhancing the growth performance of yellow catfish remains a significant concern.
Therefore, we hypothesized that rutin could serve as a functional additive in the yellow catfish diet to enhance their growth performance and health status. To test this hypothesis, we conducted a feeding experiment employing a single-factor gradient test to investigate the impact of dietary rutin on the growth performance, body composition, serum biochemical indexes, liver enzyme activities, antioxidant-related genes, intestinal morphology, and microbiome composition of yellow catfish. The obtained findings could provide valuable insights into comprehending the overall effects of dietary rutin on yellow catfish. This gathered information establishes a solid foundation for the practical utilization of rutin in feed manufacturing processes.
2. Materials and Methods
2.1. Ethics Statement
Yellow catfish are commonly farmed fish and are not endangered. The welfare of experimental fish involved in this study was approved by the Animal care and use committee of the Yangtze River Fisheries Research Institute, Chinese Academy of Fisheries Sciences. The approval code is YFI2022JM03.
2.2. Experimental Diets
The test rutin used in the experiment was ultra violet pure, and its purity was >97% (Shanghai yuanye Bio-Technology Co., Ltd., Shanghai, China). Based on the previous literature [
11,
12], dietary rutin supplementation levels were set at 0, 100 mg/kg, and 500 mg/kg, named R0, R100, and R500, respectively.
Table 1 displayed the formulation and proximate compositions of the experimental diets. The experimental diet production procedures have been described in detail in a published paper from the same laboratory [
13]. To briefly introduce the process, all the dry components were pulverized and sifted through a 60-mesh sieve, then measured based on the formulation (
Table 1). After 10 min of mixing, the dry ingredients were slowly added with fish oil and soybean oil to continue stirring until there were no obvious oily particles, and 45% distilled water were added to continue stirring for 5 min. The strips were extruded by a small meat grinder through a 1 mm sieve and dried by a dryer. The strips were simply broken by a crusher and then passed through a 20-mesh screen. The small pellets were preserved in a refrigerator set at −20 °C for future use.
2.3. Experimental Fish and Feeding Management
The experimental yellow catfish were provided by a local fish breeding facility. The experimental fish were acclimated for 4 weeks in an indoor recirculating aquiculture system (RAS) at Yangtze River Fisheries Research Institute, Chinese Academy of Fishery Sciences. During the domestication period, the fish were hand-fed three times a day with apparent satiation. Prior to the official feeding trial, the fish were fasted for 24 h. Afterwards, 270 yellow catfish with initial weight (10.27 ± 0.62 g, n = 30) were chosen and randomly divided into 9 tanks (R = 0.82 m, water depth = 0.75 m). Three tanks were randomly divided into groups and fed one of the test diets. The fish were fed three times a day (8:30, 12:30 and 16:30), and the feeding rate was 3–5% of body weight. The fish were weighed once every two weeks, and the feeding rate was adjusted according to the change of body weight. The feeding trial went on for 56 days. Feed intake and mortality of test fish were recorded daily. The filter sand tank of the RAS was backwashed with about 10% fresh water before feeding every morning and afternoon. The water temperature was kept at 28 °C. During the culture experiment, water quality parameters were measured once a week. The main water quality parameters were dissolved oxygen > 5 mg/L, pH: 6.5–7.5, total ammonia < 0.2 mg/L, nitrite < 0.05 mg/L.
2.4. Sample Collection and Processing
At the end of the feeding trial, the fish were fasted for 24 h, then counted and weighed in buckets, and the weight gain rate and specific growth rate were calculated. Six fish per tank were anesthetized with 75 mg/L MS-222(Sigma, St. Louis, MO, USA), and their body length and weight were measured. Blood was drawn through the tail artery and allowed to stand for 4 h, then centrifuged at 960× g at 4 °C for 10 min to isolate the supernatant. The viscera and liver were dissected on ice and weighed to calculate the viscerosomatic index (VSI), hepatosomatic index (HSI), and condition factor (CF). Part of the liver and midgut of 3 fish per tank were placed in frozen tubes, frozen in liquid nitrogen, and stored in the refrigerator at −80 °C for the determination of liver metabolomics, antioxidant indexes, and intestinal microorganisms. An additional portion of the liver and the midgut of 6 fish per tank was used for the preparation of HE tissues sections for histological observation.
2.5. Proximate Analysis
Approximate analyses of whole fish and experimental diets were performed according to the method of the Association of Official Agricultural Chemists (AOAC, 2000 [
14]). In brief, crude protein was determined by the Kjeldahl method; crude lipid content was determined by Soxhlet ether extraction. Moisture was determined by freeze-drying. Ash content was checked for 24 h at 550 °C using a muffle furnace. The gross energy was determined using an isothermal automatic calorimeter.
2.6. Serum Biochemical Parameters Detection
The contents of serum total protein (TP) (Sysmex, 290618), albumin (ALB) (Sysmex, 290615), triglyceride (TG) (Sysmex, 80945, 80946), total cholesterol (TCHO) (Sysmex, 290723, 290724), glucose (GLU) (Sysmex, 290713, 290714), and the activities of alkaline phosphatase (ALP) (Sysmex, 290701, 290702), aspartate aminotransferase (AST) (Sysmex, 290705, 290706) and alanine aminotransferase (ALT) (Sysmex, 290703, 290704) were detected by an automatic biochemical analyzer (Chemistry-800, Kobe, Japan). Diagnostic reagents were obtained from Sysmex Wuxi Co., Ltd. (Wuxi, China) according to standard protocols.
2.7. Analysis of SOD and MDA in Liver and Intestine
The tissue was thoroughly homogenized with 10 volumes (w/v) of ice-cold saline solution; it was centrifuged at 960× g for 10 min at 4 °C to separate the supernatant. Superoxide dismutase (SOD, A001-3-1) and malondialdehyde (MDA, A003-1-1) were measured by biochemical kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
2.8. Histopathology Evaluation
The fixed liver and intestinal samples were trimmed, dehydrated, embedded in paraffin, sectioned with a thickness of 5 μm, stained with HE, and sealed by dehydration. Images were examined using a light microscope (Leica DM2500, Leica, Solms, Germany). Five fields were randomly selected from each intestinal section to measure the number, height, width, diameter of mucosal fold, and muscular layer thickness using Image pro plus 6.0 software (IPP 6.0, Media Cybernetics Inc., Bethesda, MD, USA).
2.9. Gut Microbiota Analysis
The midgut samples from the R0, R100, and R500 groups were collected for DNA extraction. After the measurement of DNA integrity and purity, about 2 ng/μL DNA for each sample was used for 16S rRNA sequencing. The primer sets including 338F (5′-ACTCCTACGGGAGGCAGCA-3′)/806R (5′-GGACTACHVGGGTWTCTAAT-3′) were utilized to amplify variable V3–V4 regions of the 16s rRNA. The sequencing was accomplished by the staff of Shanghai Meiji Biotechnology Co., Ltd. (Shanghai, China) with the Illumina Miseq platform. Bioinformatics analysis was described in detail following our recently published literature [
15]. Representative sequences were assigned to operational taxonomic units (OTUs) with 97% similarity, and alpha diversity analysis and taxonomic composition analysis at the phylum or genus level were performed based on the clustering results.
2.10. Quantitative Real-Time PCR (qRT-PCR)
Total RNA was extracted from liver samples of groups R0, R100, and R500 using Trizol reagent (Takara Biotechnology, Tokyo, Japan), according to the manufacturer’s instructions. RNA integrity was verified by 2% agarose gel electrophoresis, and the concentration and purity were determined using a Nanodrop spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Two μg of RNA from each sample was reverse transcribed to first-strand cDNA using the primer-scriptTM RT kit (Takara Biotechnology).
qRT-PCR was performed using an ABI 7500 Real-Time PCR system with a reaction volume of 20 µL, 10 µL SYBR Premix Ex Taq II (Takara Biotechnology), 8.2 µL double-distilled water, 1 µL cDNA template, and 0.4µL primer (10 µM) included. All reactions were performed three times. Superoxide dismutase 1 (
Cu/Zn-SOD), superoxide dismutase 2 (
Mn-SOD), catalase (
CAT), glutaredoxin
3 (
GPx), alpha-induced protein 8-like 1 (
TNFα),
IL-10 (interleukin 10), lysozyme g-like (
LYZ), and ribosomal protein L13a (
Rpl13a) were selected as target genes for detection. These primer sequences are listed in
Table 2. The reference gene
β-actin was used for gene normalization. Relative mRNA expression levels were calculated according to the 2
−ΔΔCt method [
16].
2.11. Statistical Analysis
SPSS20.0 (IBM Corp., Armonk, NY, USA) was used for one-way analysis of variance (one-way ANOVA) of the experimental data, and Duncan’s multiple comparison analysis was used to analyze the significance of the difference between groups. All data results were expressed as mean ± standard deviation (mean ± SD, n = 3), and p < 0.05 indicated significant difference.
4. Discussion
In this study, the inclusion of 100 mg/kg rutin significantly enhanced growth performance and reduced the feed conversion ratio in yellow catfish. However, supplementation with 500 mg/kg rutin did not exhibit a significant effect on growth. Previous studies have demonstrated that flavonoid extracts and plant-derived compounds can act as growth promoters in aquaculture species by modulating antioxidant enzyme activities, immune responses, intestinal morphology, and microbial compositions [
17,
18]. In the case of yellow catfish, the observed improvement in growth performance and feed utilization may be attributed to enhanced antioxidant capacity and improved gut health. These factors effectively reduce oxidative stress and facilitate nutrient absorption within the fish gut. Similar findings have also been reported for other commercially important fish species such as Nile tilapia [
8] and grass carp [
6]. However, a relatively higher concentration of rutin in the diet may contain certain anti-nutritional substances, leading to a decrease in feed intake and subsequently causing growth retardation in fish. Moreover, supplementation of dietary rutin at 100 mg/kg significantly reduced the crude protein content in the whole body, indicating that rutin could potentially function as a regulatory factor involved in nutrient metabolism within this particular species.
SOD is an important indicator for the assessment of oxidative damage in fish. SOD has a function in scavenging the free radicals to protect the body from oxidative stress, and its activity is positively associated with the health conditions in fish species [
19]. Rutin is reported to trigger the increment in SOD activity, remove excess reactive oxygen species (ROS), and therefore, improve the antioxidant system [
20]. Likewise, the addition of rutin in the diet significantly increased the activities of many kinds of antioxidant enzymes in the brain, liver, kidney, and gill of silver catfish [
7]. In the current experiment, the upward trend or significant elevation of SOD activity in both liver and intestine was detected when fish were fed the diets with 100 mg/kg, demonstrating that rutin has ameliorative roles on the antioxidant capacity of yellow catfish, and relieves oxidative-caused injury by counteracting the generation of free radicals. This data may further evidence that rutin could serve as a potent agent in the improvement of antioxidative response in aquatic organisms [
21]. MDA is one of several end products of the lipid peroxidation chain and indirectly reflects the degree of oxidative damage [
22]. In the present study, MDA in the liver and intestine of yellow catfish was found to be inversely correlated with increasing dietary rutin concentrations, which may indicate that rutin is able to prevent peroxidation levels in fish tissues. Similar results have been found in rats [
23].
Moreover, the mRNA expressions of antioxidant-related genes were examined in the liver. Cu/Zn-SOD is the most studied of superoxide dismutase; Cu is responsible for the catalytic activity and Zn is mainly in charge of the maintenance of its structure [
24]. Mn-SOD is located in the soluble matrix of the mitochondria and has crucial roles in eliminating ROS in cells [
25]. The product encoded by the
CAT gene is a major antioxidant enzyme that converts hydrogen peroxide to oxygen and water in an energy efficient way [
26]. GPx represents a key enzyme in the cellular machinery that catalyzes the reduction of hydroperoxides [
27]. The present data showed that dietary rutin at 500 mg/kg significantly induced the transcriptional expression of
Cu/Zn-SOD,
Mn-SOD,
CAT, and
GPx genes, which may further support the enhancement of the antioxidant response in response to dietary rutin treatment. Furthermore, several genes, including
TNFα,
IL-10,
LYZ in liver, were significantly up-regulated as fish were fed the diet with 500 mg/kg rutin, indicating that rutin could exert an anti-inflammatory effect on yellow catfish. A similar pattern of response has been reported on tilapia [
28]. Fish fed the 100 mg/kg rutin diet had the lowest transcriptional expression of Mn-SOD, CAT, and GPx genes, suggesting that appropriate rutin supplementation had protective effects against oxidative stimulation.
Serum biochemical parameters generally reflect the fish physiological and metabolic conditions under nutritional manipulation or environmental stress [
29]. TP could indicate protein metabolism in fish and maintain the balance of intravascular osmotic pressure [
30]. ALB is considered a key modulator of plasma oncotic pressure within the vascular space [
31]. ALP is an important non-specific immune enzyme for evaluating the fish immunity status in aquaculture [
32]. In this study, dietary rutin at a concentration of 500 mg/kg significant elevated the values of serum TP, ALB, and ALP, indicating the beneficial effects of rutin on the enhancement in immune response and metabolic ability of yellow catfish. Previous findings on mammals also confirmed that dietary rutin could effectively increase the levels of serum TP and ALB, and delay the progression of liver fibrosis by down-regulating the expression of transforming growth factor-β1 (TGF-β1) and collagen I [
33]. In addition, the significant reduction of serum GLU was observed in fish fed the rutin-supplemented diets in comparison with that of fish fed the control diet. This may be explained by the fact that flavonoid substances could stimulate the liver to liberate the glucose, induce the activation of insulin receptors or pathways, and thereby facilitate the absorption of glucose in serum [
34]. This observation is in accordance with that of Nazer et al. [
35], who indicated that dietary rutin can significantly decrease the blood glucose concentrations in rainbow trout (
Oncorhynchus mykiss). In addition, the change in blood glucose is correlated with hepatic enzymes activities, which may suggest that rutin has a beneficial influence on liver function. Notably, reduced AST activity observed in fish fed with rutin further supports its potential for improving overall liver function when compared to the control diet.
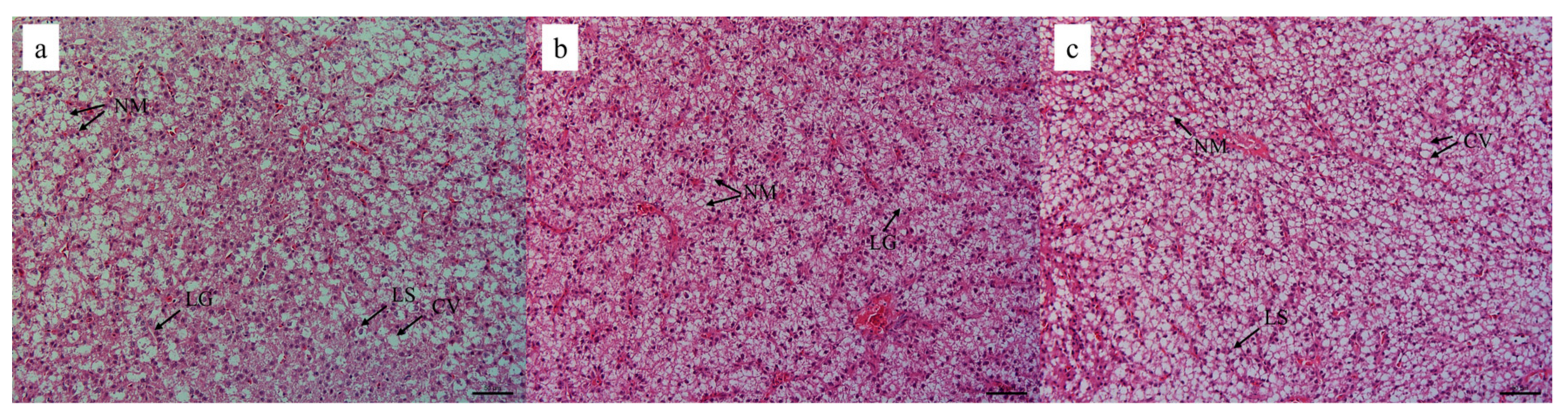
H&E staining is commonly used for histopathology evaluation in fish due to that it can clearly illustrate cellular structures with remarkable signatures [
36]. Excessive rutin intake has been implicated in impairing the health of liver tissue in the GIFT strain of tilapia [
28]. In this research, the liver cellular structures were intact and tightly arranged when fish were fed the 100 mg/kg rutin diet. However, the liver cells treated with 500 mg/kg rutin in the diet showed larger cell volume, serious nuclear migration, hepatocyte vacuolation, and augmented intercellular space. It might be attributed to the occurrence of apoptotic cells or pro-inflammatory cytokines triggered by a high dose of rutin. The intestinal tract is the primary organ involved in fish digestion, nutritional absorption, and intestinal immunity. Some typical indicators such as villus numbers, villus length, muscular thickness and lumen diameter are used for the assessment of intestinal health [
37]. Generally, the strong ability of nutrient absorption is a positive association with the greater absorptive area and increased intestinal villi [
21]. Muscular thickness is closely related to the rhythmic contraction of the gut tract and efficient digestion of chyme [
38]. In this study, the significant increase in values of villus length, muscular thickness, and lumen diameter were observed in fish fed the 100 mg/kg rutin diet, indicating that rutin has beneficial effects on intestinal morphological structures, and thus, effectively prompt its absorptive capability as well as nutrient consumption.
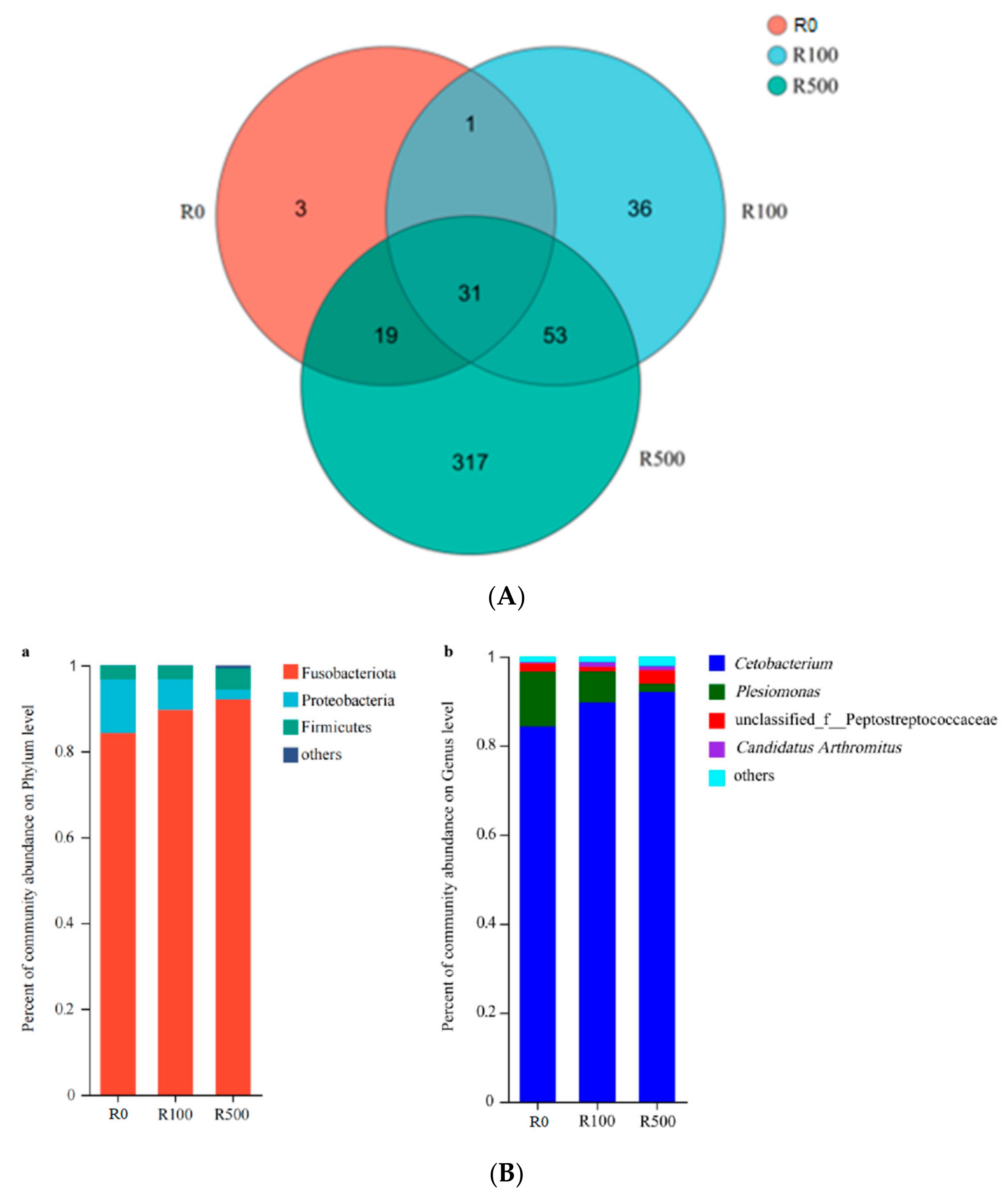
There exists a substantial population of microorganisms within the fish intestine which has established a dynamic and intricate microenvironment over an extended period of evolution [
39]. The gut microbiota is known to uphold homeostasis and influence nutrient absorption, pathogen assimilation, as well as growth and development in fish [
40]. Furthermore, the intestinal flora plays a pivotal role in regulating host immunity and maintaining immune homeostasis [
41]. In the present study, Fusobacteria, Proteobacteria, and Firmicutes were identified as the predominant phyla within the intestinal flora of yellow catfish. These findings align with those reported by Wu et al. [
42]. Notably, Proteobacteria represents the largest phylum encompassing bacteria or pathogens; an elevated abundance of Proteobacteria can disrupt the gut microecological environment in animal hosts while concurrently increasing disease risks [
43]. Firmicutes have been demonstrated to possess the capacity for carbohydrate transformation, cellulose decomposition promotion, and polysaccharide fermentation. They also enhance fish digestion and nutrient absorption while aiding in the maintenance of gut immune system homeostasis in hosts [
44]. In our study, the inclusion of rutin at levels of 100 mg/kg and 500 mg/kg in diets significantly augmented the abundance of Fusobacteria and Firmicutes, but reduced the richness of Proteobacteria. This suggests that appropriate dietary supplementation with rutin may potentially mitigate stress-related diseases. At the genus level,
Cetobacterium, Plesiomonas, and
Peptostreptococcaceae were identified as dominant genera within the gut microbiota of yellow catfish.
Cetobacterium can produce vitamin B
12 in the process of carbohydrates fermentation and play crucial functions in the nutritional metabolism [
45]. Pseudomonas is a common pathogen existing in aquatic water and fish body, of which
Peptostreptococcaceae predominated in gut diseases [
46]. The present investigation showed that the increased abundance of
Cetobacterium and the decreased
Plesiomonas richness were determined on fish fed the diets with 100 mg/kg or 500 mg/kg rutin. These data suggested that dietary rutin has positive effects on the improvement of intestinal structure and microbial composition, which is beneficial to the gut health of yellow catfish.







{kind=link}
{kind=link}
{kind=link}
{kind=link}