
Selected Aspects of Reproductive Behavior of American Mink (Neovison vison) under Artificial Conditions

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Budzyńska, M. Współczesne zagadnienia w badaniach i nauczaniu dobrostanu zwierząt. Wiad. Zootech. 2015, 1, 58–64. [Google Scholar]
- Pellis, S.M.; Pellis, V.C.; Pelletier, A.; Leca, J.B. Is play a behaviour system, and, if so, what kind? Behav. Process. 2019, 160, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Banks, E.M. Behavioural research to answer questions about animal welfare. J. Anim. Sci. 1982, 52, 434–446. [Google Scholar] [CrossRef] [PubMed]
- Tumanowicz, J.; Zachwieja, A.; Kruszynski, W. Po co ten behawior? Prz. Hod. 2009, 77, 22–25. [Google Scholar]
- Burghardt, G.; Bowers, R. From instinct to behaviour systems: An integrated approach to ethological psychology. In APA Handbook of Comparative Psychology; American Psychological Association: Washington, DC, USA, 2017. [Google Scholar]
- Bowers, R. Six clarifications for behaviour systems. Behav. Process. 2020, 170, 103987. [Google Scholar] [CrossRef] [PubMed]
- Yaribeygi, H.; Panahi, Y.; Sahraei, H.; Johnston, T.P.; Sahebkar, A. The impact of stress on body function—A review. EXCLI J. 2017, 16, 1057–1072. [Google Scholar] [PubMed]
- Tilbrook, A.J.; Turner, A.I.; Clarke, I.J. Stress and reproduction: Central mechanisms and sex differences in non-rodent species. Stress 2002, 5, 83–100. [Google Scholar] [CrossRef]
- García, P. Female multiple copulation in the invasive American Mink (Neovison vison). North West. J. Zool. 2010, 6, 138–139. [Google Scholar]
- Amstislavsky, S.; Ternovskaya, Y. Reproduction in mustelids. Anim. Reprod. Sci. 2000, 60, 571–581. [Google Scholar] [CrossRef]
- Ellis, L.C.; Groesbeck, M.D.; Howell, R.E. Pineal gland—Pituitary (alpha-MSH) interrelationships in fur priming and reproductive cycles in mink (Mustela vison). Prog. Clin. Biol. Res. 1982, 9, 197–205. [Google Scholar]
- Ravault, J.P.; Martinet, L.; Bonnefond, C.; Claustrat, B.; Brun, J. Diurnal variations of plasma melatonin concentrations in pregnant or pseudopregnant mink (Mustela vison) maintained under different photoperiods. J. Pineal Res. 1986, 3, 365–373. [Google Scholar] [CrossRef]
- Holcomb, L.C.; Schaible, P.J.; Ringer, R.K. The effects of varied lighting regimes on reproduction in mink. Mich. Agric. Exp. Stn. Quart. Bull. 1962, 44, 666–678. [Google Scholar]
- Murphy, B.D. Precocious Induction of Luteal Activation and Termination of Delayed Implantation in Mink with the Dopamine Antagonist Pimozide. Biol. Reprod. 1983, 29, 658–662. [Google Scholar] [CrossRef] [PubMed]
- Wehrenberg, W.B.; Kurt, K.J.; Hutz, R.J. Effects of equine chorionic gonadotropin on reproductive performance in anoestrous mink. J. Anim. Sci. 1992, 70, 499–502. [Google Scholar] [CrossRef]
- Lagerkvist, G.; Einarsson, E.J.; Forsberg, M.; Gustafsson, H. Profiles of oestradiol-17B and progesterone and follicular development during the reproductive season in mink (Mustela vison). J. Reprod. Fertil. 1992, 94, 11–21. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Douglas, D.A.; Pierson, R.A.; Murph, B.D. Ovarian follicular development in mink (Mustela vison). J. Reprod. Fertil. 1994, 100, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Polejaeva, I.A.; Reed, W.A.; Bunch, T.D.; Ellis, L.C.; White, K.L. Prolactin—Inducted termination of obligate diapause of mink (Mustela vison) blastocystys in vitro and subsequent establishment of embryonic stem—Like cells. J. Reprod. Fert. 1997, 109, 229–236. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sundquvist, C.; Amador, A.G.; Bartke, A. Reproduction and fertility in the mink (Mustela vison). Reproduction 1989, 85, 413–441. [Google Scholar] [CrossRef]
- Boissin-Agasse, L.; Boissin, J.; Ortavant, R. Circadian Photosensitive Phase and Photoperiodic Control of Testis Activity in the Mink (Mustela vison Peale and Beauvois), a Short-Day Mammal. Biol. Reprod. 1982, 26, 110–119. [Google Scholar] [CrossRef]
- Klotchkov, D.V.; Trapezov, O.V.; Kharlamova, A.V. Folliculogenesis, onset of puberty and fecundity of Mink (Mustela vison Scherb) selectively bred for docility or aggressiveness. Theriogenology 1998, 49, 1545–1553. [Google Scholar] [CrossRef]
- Pilbeam, T.E.; Concannon, P.W.; Travis, H.F. The Annual Reproductive Cycle of Mink (Mustela vison). J. Anim. Sci. 1979, 48, 578–584. [Google Scholar] [CrossRef] [PubMed]
- Gulevich, R.G.; Klotchkov, D.V.; Ivanova, L.N.; Osadchuk, L.V. Gonadal function in mink under artificial photoperiods. J. Reprod. Fertil. 1995, 103, 147–152. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fink, R.; Tauson, A.H.; Forsberg, M. Influence of different planes of energy supply prior to the breeding season on blood metabolites in female mink (Mustela vison). Reprod. Nutr. Dev. 1998, 38, 107–116. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Travis, H.F.; Pilbeam, T.E.; Gardner, W.J.; Cole, R.S. Relationship of Vulvar Swelling to Estrus in Mink. J. Anim. Sci. 1978, 46, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Persson, S. The Mink (Mustela vison) as an indicator of environmental reproductive toxicity. Swed. Univ. Agric. Sci. 2007, 50, 1–23. [Google Scholar]
- Hatler, D.F. The Coastal Mink on Vancouver Island, British Columbia. Ph.D. Thesis, University of British Columbia, Vancouver, BC, Canada, 1976. [Google Scholar]
- Birks, J.D.S.; Linn, I.J. Studies on the home range of feral mink (Mustela vison). Symp. Zool. Soc. Lond. 1982, 49, 231–257. [Google Scholar]
- Ireland, M.C. The Behaviour and Ecology of the American Mink Mustela vison (Schreber) in Acoastal Habitat. Ph.D. Thesis, Durham University, Durham, UK, 1990. [Google Scholar]
- Dunstone, N. The Mink; T. & A. D. Poyser Publishing: London, UK, 1993. [Google Scholar]
- Hansson, A. The physiology of reproduction in mink. Acta Zool. 1947, 28, 1–136. [Google Scholar] [CrossRef]
- Enders, R.K. Reproduction in the mink (Mustela vision). Proc. Am. Philos. Soc. 1952, 96, 691–755. [Google Scholar]
- Seremak, B.; Dziadosz, M.; Lasota, B.; Felska-Błaszczyk, L.; Pławski, K.; Masłowska, A.; Mieleńczuk, G. Effect of the quality of handling and care on the reproduction parameters of mink. Acta Sci. Pol. Zootech. 2011, 10, 93–102. [Google Scholar]
- Seremak, B.; Dziadosz, M.; Dworecka-Borczyk, M. Multiple paternity in American Mink: Using males of different color lines allows mating efficiency evaluation. Acta Sci. Pol. Zootech. 2020, 19, 55–62. [Google Scholar] [CrossRef]
- Dallaire, A.J.; Mason, G.J. Juvenile rough-and-tumble play predicts adult sexual behaviour in American mink. Anim. Behav. 2017, 123, 81–89. [Google Scholar] [CrossRef]
- Journal Laws of 2015 Item 266, ACT on the Protection of Animals Used for Scientific or Educational Purposes. Available online: https://isap.sejm.gov.pl/isap.nsf/download.xsp/WDU20150000266/U/D20150266Lj.pdf (accessed on 15 October 2022).
- Brinck, C.; Gerell, R.; Odham, G. Anal pouch secretion in mink Mustela vison. Oikos 1978, 30, 68–75. [Google Scholar] [CrossRef]
- Lodé, T. Conspecific recognition and mating in stone marten Martes foina. Acta Thériol. 1991, 36, 275–283. [Google Scholar] [CrossRef]
- Poole, T.B. Aspects of aggressive behaviour in polecats. Z. Tierpsychol. 1967, 24, 351–369. [Google Scholar] [PubMed]
- Coombes, H.A.; Stockley, P.; Hurst, J.L. Female chemical signalling underlying reproduction in mammals. J. Chem. Ecol. 2018, 44, 851–873. [Google Scholar] [CrossRef] [PubMed]
- Crump, D.R. Anal gland secretion of the ferret (Mustela putorius formafuro). J. Chem. Ecol. 1980, 6, 837–844. [Google Scholar] [CrossRef]
- Gorman, M.L.; Kruuk, H.; Leitch, A. Social functions of the subcaudal scent gland secretion of the European badger Meles meles (Carnivora: Mustelidae). J. Zool. 1984, 203, 549–559. [Google Scholar] [CrossRef]
- Gorman, M.L.; Trowbridge, B.J. The Role of Odor in the Social Lives of Carnivores. In Carnivore Behavior, Ecology, and Evolution; Gittleman, J.L., Ed.; Cornell University Press: Ithaca, NY, USA, 1989; pp. 57–88. [Google Scholar]
- Buesching, C.D.; Waterhouse, J.P.; Macdonald, D.W. Gas chromatographic analysis of the subcaudal gland secretion of the European badger (Meles meles) part I: Chemical differences related to individual-specific parameters. J. Chem. Ecol. 2002, 28, 41–56. [Google Scholar] [CrossRef]
- Elofson, L.; Lagerkvist, G.; Gustafsson, H.; Einarsson, S. Mating systems and reproduction in mink. Acta Agric. Scand. 1989, 39, 23–41. [Google Scholar] [CrossRef]
- Fleming, M. The Adaptive Significance of Multiple Mating in Female Mink (Mustela vison) and Its Effects on the Mating System. Ph.D. Thesis, Washington University, Washington, DC, USA, 1996. [Google Scholar]
- Venge, O. Reproduction in the fox and mink. Anim. Breed. Abstr. 1959, 27, 129–145. [Google Scholar]
- Johansson, I.; Venge, O. Relation of the mating interval to the occurrence of superfetation in the mink. Acta Zool. 1951, 32, 255–258. [Google Scholar] [CrossRef]
- Shackelford, R.M. Superfetation in the ranch mink. Am. Nat. 1952, 86, 311–319. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| First Mating Period (6 March–11 March) | |||||
|---|---|---|---|---|---|
| Male Number | Number of Copulations | Mean Copulation Time | Standard Error SE | Min. | Max. |
| [h:m:s] | [h:m:s] | [h:m:s] | |||
| 1 | 2 | 00:50:45 | 00:19:21 | 00:31:23 | 01:10:07 |
| 2 | 11 | 00:53:12 | 00:06:09 | 00:21:20 | 01:23:26 |
| 3 | 3 | 00:57:25 | 00:19:50 | 00:26:30 | 01:34:24 |
| 4 | 12 | 00:32:46 | 00:03:40 | 00:15:11 | 00:59:19 |
| 5 | 5 | 00:47:18 | 00:08:38 | 00:19:43 | 01:06:50 |
| 6 | 12 | 00:52:13 | 00:08:12 | 00:14:24 | 02:04:42 |
| 7 | 10 | 00:47:00 | 00:07:33 | 00:22:36 | 01:44:25 |
| 8 | 9 | 00:43:09 | 00:06:27 | 00:14:37 | 01:11:00 |
| 9 | 8 | 01:08:32 | 00:07:58 | 00:40:14 | 01:40:10 |
| 10 | 13 | 00:46:22 | 00:04:32 | 00:13:11 | 01:09:28 |
| 11 | 12 | 01:06:57 | 00:06:52 | 00:24:48 | 01:33:44 |
| 12 | 8 | 01:14:31 | 00:09:16 | 00:25:30 | 01:40:57 |
| Total | 105 | 00:52:37 | |||
| Second Mating Period (12 March–17 March) | |||||
|---|---|---|---|---|---|
| Male Number | Number of Copulations | Mean Copulation Time | Standard Error SE | Min. | Max. |
| [h:m:s] | [h:m:s] | [h:m:s] | |||
| 1 | 5 | 01:13:08 | 00:21:44 | 00:24:00 | 02:15:20 |
| 2 | 9 | 01:12:47 | 00:06:04 | 00:37:34 | 01:42:53 |
| 3 | 7 | 01:00:27 | 00:10:24 | 00:17:50 | 01:31:04 |
| 4 | 12 | 00:30:26 | 00:04:17 | 00:13:46 | 00:49:40 |
| 5 | 7 | 01:28:18 | 00:14:21 | 00:34:12 | 02:28:13 |
| 6 | 10 | 00:59:38 | 00:09:08 | 00:22:49 | 01:58:18 |
| 7 | 12 | 01:08:41 | 00:09:07 | 00:20:15 | 02:07:05 |
| 8 | 6 | 01:04:42 | 00:13:50 | 00:21:06 | 01:55:08 |
| 9 | 10 | 01:07:03 | 00:07:53 | 00:41:40 | 01:57:38 |
| 10 | 13 | 00:54:51 | 00:07:18 | 00:13:55 | 01:36:13 |
| 11 | 9 | 01:24:02 | 00:07:35 | 00:48:30 | 02:01:25 |
| 12 | 8 | 01:14:15 | 00:11:32 | 00:42:18 | 02:17:41 |
| Total | 108 | 01:04:32 | |||
| Changes in Mean Copulation Time Based on the Mating Period | ||||
|---|---|---|---|---|
| Male Number | t | df | p | d |
| 1 | −0.59 | 5 | 0.580 | 0.53 |
| 2 | −2.24 | 18 | 0.038 | 1.05 |
| 3 | −0.15 | 8 | 0.884 | 0.11 |
| 4 | 0.41 | 22 | 0.685 | 0.18 |
| 5 | −2.20 | 10 | 0.053 | 1.39 |
| 6 | −0.60 | 20 | 0.553 | 0.27 |
| 7 | −1.78 | 20 | 0.090 | 0.80 |
| 8 | −1.58 | 13 | 0.139 | 0.87 |
| 9 | 0.13 | 16 | 0.897 | 0.07 |
| 10 | −0.99 | 24 | 0.334 | 0.40 |
| 11 | −1.66 | 19 | 0.114 | 0.76 |
| 12 | 0.02 | 14 | 0.986 | 0.01 |
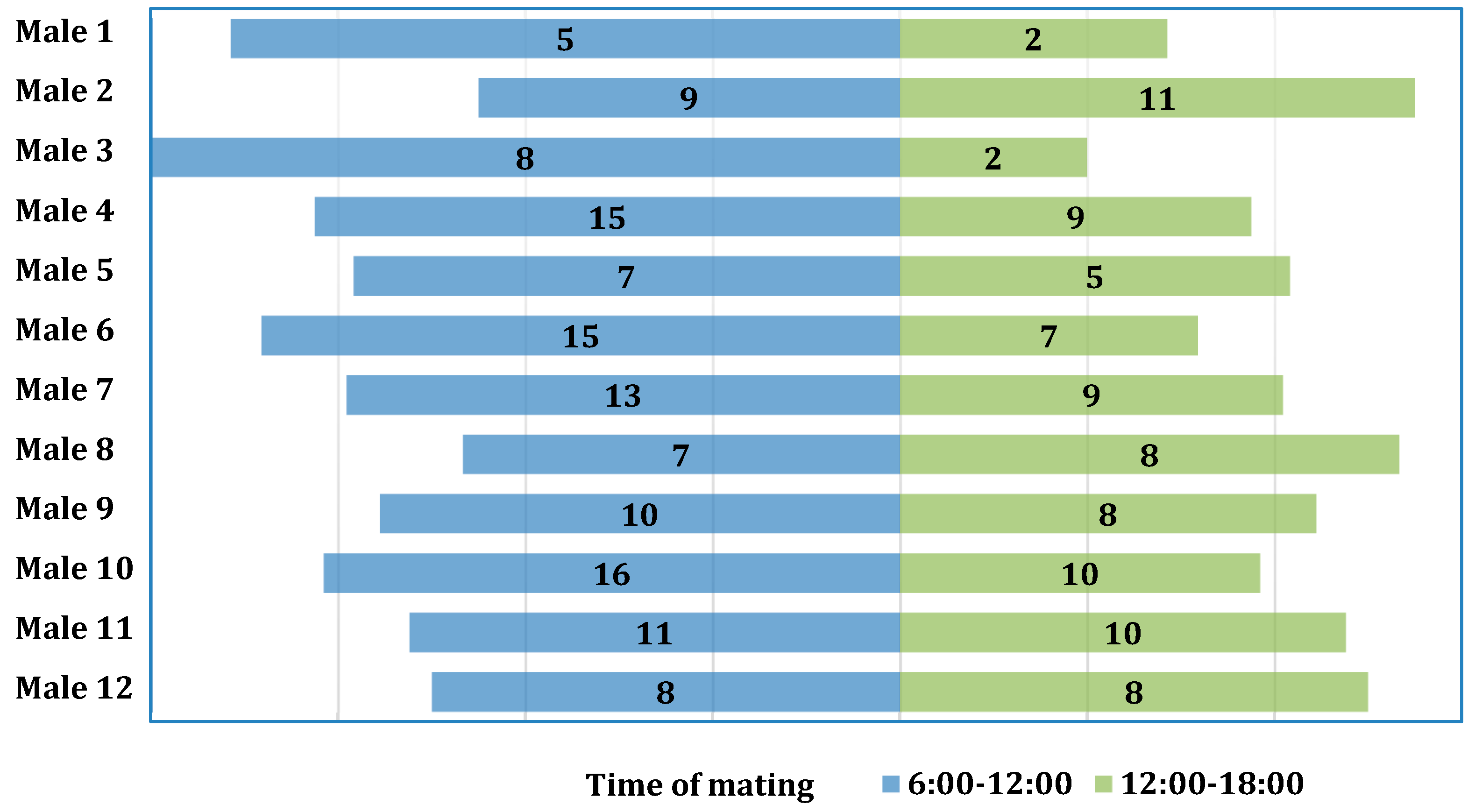
| First Time Interval 6.00–12.00 a.m. | |||||
|---|---|---|---|---|---|
| Male Number | Number of Copulations | Mean Copulation Time | Standard Error SE | Min | Max |
| [h:m:s] | [h:m:s] | [h:m:s] | |||
| 1 | 5 | 01:05:47 | 00:18:29 | 00:31:23 | 02:15:20 |
| 2 | 9 | 00:58:27 | 00:04:30 | 00:38:01 | 01:16:40 |
| 3 | 8 | 01:00:57 | 00:08:42 | 00:26:30 | 01:34:24 |
| 4 | 15 | 00:32:09 | 00:03:13 | 00:13:46 | 00:48:44 |
| 5 | 7 | 01:12:04 | 00:17:03 | 00:19:43 | 02:28:13 |
| 6 | 15 | 00:55:54 | 00:07:46 | 00:23:28 | 02:04:42 |
| 7 | 13 | 00:59:33 | 00:09:10 | 00:22:36 | 02:07:05 |
| 8 | 7 | 00:36:37 | 00:06:18 | 00:21:06 | 01:09:14 |
| 9 | 10 | 01:07:07 | 00:06:01 | 00:40:14 | 01:40:10 |
| 10 | 16 | 00:50:14 | 00:04:55 | 00:13:11 | 01:12:09 |
| 11 | 11 | 01:19:50 | 00:08:39 | 00:24:48 | 02:01:25 |
| 12 | 8 | 01:15:13 | 00:12:46 | 00:25:30 | 02:17:41 |
| Total | 124 | 00:57:41 | -- | ||
| Second Time Interval 12.00 a.m.–06.00 p.m. | |||||
|---|---|---|---|---|---|
| Male Number | Number of Copulations | Mean Copulation Time [h:m:s] | Standard Error SE | Min [h:m:s] | Max [h:m:s] |
| 1 | 2 | 01:09:08 | 00:45:08 | 00:24:00 | 01:54:16 |
| 2 | 11 | 01:04:55 | 00:08:00 | 00:21:20 | 01:42:53 |
| 3 | 2 | 00:53:53 | 00:36:03 | 00:17:50 | 01:29:56 |
| 4 | 9 | 00:30:42 | 00:05:21 | 00:15:11 | 00:59:19 |
| 5 | 5 | 01:10:02 | 00:11:52 | 00:42:56 | 01:50:12 |
| 6 | 7 | 00:54:56 | 00:09:47 | 00:14:24 | 01:18:13 |
| 7 | 9 | 00:57:47 | 00:08:48 | 00:20:15 | 01:42:34 |
| 8 | 8 | 01:05:02 | 00:10:07 | 00:14:37 | 01:55:08 |
| 9 | 8 | 01:08:27 | 00:10:16 | 00:41:40 | 01:57:38 |
| 10 | 10 | 00:51:12 | 00:08:18 | 00:13:55 | 01:36:13 |
| 11 | 10 | 01:08:10 | 00:05:43 | 00:39:46 | 01:33:44 |
| 12 | 8 | 01:13:32 | 00:07:28 | 00:45:30 | 01:45:06 |
| Total | 89 | 01:00:01 | -- | -- | -- |
| Change in Mean Copulation Time with Regard to Time Interval | ||||
|---|---|---|---|---|
| Male Number | t | df | p | d |
| 1 | −0.09 | 5 | 0.935 | 0.08 |
| 2 | −0.66 | 18 | 0.517 | 0.31 |
| 3 | 0.31 | 8 | 0.767 | 0.22 |
| 4 | 0.25 | 22 | 0.807 | 0.11 |
| 5 | 0.09 | 10 | 0.931 | 0.06 |
| 6 | 0.07 | 20 | 0.943 | 0.03 |
| 7 | 0.13 | 20 | 0.896 | 0.06 |
| 8 | −2.30 | 13 | 0.039 | 1.28 |
| 9 | −0.12 | 16 | 0.908 | 0.06 |
| 10 | −0.11 | 24 | 0.917 | 0.04 |
| 11 | 1.10 | 19 | 0.285 | 0.51 |
| 12 | 0.11 | 14 | 0.911 | 0.06 |
| Parameter | B | SE | β | t | p |
|---|---|---|---|---|---|
| 40.70 | 6.05 | 6.73 | 0.000 | ||
| Day | 11.42 | 3.51 | 0.21 | 3.13 | 0.002 |
| Day | Number of Copulations | Number of Males Mating | Mean Copulation Time | Standard Error SE | Min | Max |
|---|---|---|---|---|---|---|
| [h:m:s] | [h:m:s] | [h:m:s] | ||||
| 6 March | 19 | 11 | 00:44:31 | 00:04:03 | 00:21:20 | 01:31:20 |
| 7 March | 17 | 10 | 00:51:56 | 00:05:59 | 00:14:37 | 01:34:24 |
| 8 March | 25 | 11 | 00:45:09 | 00:03:41 | 00:13:11 | 01:09:37 |
| 9 March | 17 | 11 | 00:57:50 | 00:07:13 | 00:15:11 | 02:04:42 |
| 10 March | 9 | 8 | 00:52:53 | 00:09:12 | 00:18:39 | 01:25:02 |
| 11 March | 18 | 10 | 01:07:05 | 00:05:36 | 00:19:43 | 01:44:25 |
| 12 March | 19 | 11 | 00:58:24 | 00:07:59 | 00:15:46 | 02:28:13 |
| 13 March | 24 | 12 | 00:56:30 | 00:05:14 | 00:13:46 | 02:07:05 |
| 14 March | 21 | 12 | 01:06:31 | 00:06:51 | 00:17:30 | 01:57:38 |
| 15 March | 14 | 9 | 01:17:49 | 00:08:12 | 00:21:06 | 02:17:41 |
| 16 March | 17 | 9 | 00:59:39 | 00:07:11 | 00:17:28 | 02:01:25 |
| 17 March | 13 | 7 | 01:17:13 | 00:08:48 | 00:24:36 | 02:15:20 |
| Male | Number of Copulations | R Value | p Value |
|---|---|---|---|
| 1 | 7 | 0.43 | 0.337 |
| 2 | 20 | 0.48 | 0.032 |
| 3 | 10 | 0.01 | 0.987 |
| 4 | 23 | 0.01 | 0.974 |
| 5 | 12 | 0.60 | 0.039 |
| 6 | 22 | 0.26 | 0.236 |
| 7 | 22 | 0.05 | 0.844 |
| 8 | 15 | 0.35 | 0.201 |
| 9 | 18 | 0.29 | 0.247 |
| 10 | 27 | 0.21 | 0.289 |
| 11 | 21 | 0.60 | 0.004 |
| 12 | 18 | 0.13 | 0.616 |
| Male | Number of Copulations | Mean Time before Copulation | Mean Copulation Time | Spearman’s Correlation |
|---|---|---|---|---|
| [h:m:s] | [h:m:s] | R Value | ||
| 1 | 7 | 00:14:24 | 01:06:44 | 0.00 |
| 2 | 20 | 00:21:46 | 01:02:01 | −0.40 |
| 3 | 10 | 00:23:52 | 00:59:33 | −0.18 |
| 4 | 23 | 00:10:13 | 00:31:37 | −0.20 |
| 5 | 12 | 00:33:12 | 01:11:14 | −0.67 * |
| 6 | 22 | 00:43:52 | 00:55:36 | 0.04 |
| 7 | 22 | 00:24:25 | 00:58:50 | −0.41 |
| 8 | 15 | 00:04:50 | 00:51:47 | −0.55 * |
| 9 | 18 | 00:02:41 | 01:07:43 | −0.48 * |
| 10 | 27 | 00:11:27 | 00:50:37 | 0.05 |
| 11 | 21 | 00:06:34 | 01:14:17 | 0.41 |
| 12 | 18 | 00:27:50 | 01:14:23 | −0.53 |
| Total | 17.92 | 00:18:45 | 01:00:23 | −0.27 ** |
| Spearman Correlation, r Value | |
|---|---|
| Types of Behavior | Copulation Time |
| Rest | −0.14 |
| Race | −0.10 |
| Sniffing the female | −0.24 ** |
| Rubbing | −0.65 * |
| Observation | −0.06 |
| Care | 0.02 |
| Game | −0.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seremak, B.; Pilarczyk, B.; Wojciechowska, A.; Tomza-Marciniak, A. Selected Aspects of Reproductive Behavior of American Mink (Neovison vison) under Artificial Conditions. Animals 2023, 13, 3346. https://doi.org/10.3390/ani13213346
Seremak B, Pilarczyk B, Wojciechowska A, Tomza-Marciniak A. Selected Aspects of Reproductive Behavior of American Mink (Neovison vison) under Artificial Conditions. Animals. 2023; 13(21):3346. https://doi.org/10.3390/ani13213346
Chicago/Turabian StyleSeremak, Beata, Bogumiła Pilarczyk, Aleksandra Wojciechowska, and Agnieszka Tomza-Marciniak. 2023. "Selected Aspects of Reproductive Behavior of American Mink (Neovison vison) under Artificial Conditions" Animals 13, no. 21: 3346. https://doi.org/10.3390/ani13213346
APA StyleSeremak, B., Pilarczyk, B., Wojciechowska, A., & Tomza-Marciniak, A. (2023). Selected Aspects of Reproductive Behavior of American Mink (Neovison vison) under Artificial Conditions. Animals, 13(21), 3346. https://doi.org/10.3390/ani13213346