Simple Summary
With the advance of modern civilization, there has been intensive contact among large parts of the world. This has not only encouraged increased travel and trade, but has also led to the anthropogenic introduction of animal and plant species into new areas of settlement. Such neozoa (invasive species) encounter different and sometimes completely new environmental conditions, which requires the adaptation of the neozoa to their new habitat. Under certain circumstances, the neozoa may even have significant survival advantages, for example, if the prey–hunter constellation shifts in their favor. The North American raccoon (Procyon lotor) was introduced into Europe by intentional and accidental releases in the 20th century and spread throughout most of Europe in the following decades. The first release was in Germany, in Central Europe, which is still the distribution hotspot of the European raccoon. Over time, raccoons were released into the wild for hunting and subsequently spread. There have also been repeated unintentional releases of raccoons by breeders, fur farms, and private pet owners. These releases have accelerated the spread of the raccoon in Europe. The influence of the raccoon on the native fauna and flora has been insufficiently studied so far, but it seems to be only marginal according to current knowledge.
Abstract
The raccoon (Procyon lotor) is a North American half-bear that is present in much of Europe and Asia as a result of both accidental and planned releases. In Europe, raccoons were introduced primarily as a source of fur for the fur industry. In the 1930s, raccoons were released into the wild in Central Europe. At the same time, animals from fur farms and private holdings continued to enter the wild. In the following decades, the raccoon spread over large parts of Europe. In addition to the invasive spread of the Central European initial population, individual releases of raccoons occurred frequently, mainly in Southern Europe. The high adaptability of the raccoon favors its expansion into new habitats. It has a high reproductive rate, is very mobile, and encounters few predators in Europe. Raccoons have recently become a topic of interest when large raccoon populations have colonized suburban and urban areas. Despite the proximity of raccoons and humans, however, there have been hardly any conflicts to date, unlike in North America. A significant negative impact on the native fauna has been suspected but not proven. Raccoons have been identified as vectors of zoonotic diseases. Nevertheless, monitoring of the increasing numbers of raccoons in Europe seems advisable.
1. Introduction
The raccoon (Procyon lotor) is a member of the half-bear predator group (Procyonidae) native to North America that now occurs as a neozoon through artificial dispersal in other parts of the world [1,2,3]. In addition to the raccoon (Procyon), the Procyonidae include coatis (Nasua), ringtails (Bassariscus), and olingos (Bassaricyon). The raccoons include the North American raccoon (Procyon lotor), the most common representative of this group, as well as the crab-eating raccoon (Procyon cancrivorus) and the Cozumel raccoon (Procyon pygmaeus) [4]. Hereinafter, only the species Procyon lotor is discussed and is referred to as ‘raccoon’ in the text.
Adult raccoons reach a body length of up to 60 cm and a body weight of up to 12 kg. The body markings are gray to black, rarely red or brown, and occur independently of the geographical location of the habitat [5]. The body volume and body weight of the animals increases towards the northern ranges. This is in accordance with Bergmann’s animal geographic rule, where the average body size of animals increases towards the poles [4].
Raccoons are nocturnal omnivores and generalists. They occur in deciduous and mixed forests, mostly near water bodies. The population is usually larger near agricultural land or settlement areas [6]. Raccoons have also settled in suburban and urban habitats for decades [2,6,7,8]. Their diet includes plants and invertebrates, and the composition of the diet depends on the habitat and season [9]. In anthropogenically influenced habitats, a large proportion of the diet is provided by human sources [10,11]. The distribution of food resources influences the distribution of raccoons. In urban habitats, the increased supply of food and retreat opportunities leads to reduced territory sizes with higher population densities. The area of the territory is up to 702 ha in forests, up to 300 ha in rural habitats, and up to 79 ha in urban areas [11,12,13]. The population density is up to eight individuals/km2 in forest areas and five individuals/km2 in rural areas and increases significantly in urban areas to up to 333 individuals/km2 [8,14].
Raccoons have varying but comparatively high reproductive rates, depending on the habitat and the resources that are available in it. The mating season begins at the beginning of the year and can last until August. If a female does not become pregnant early in the year or lost her offspring, she may become pregnant again in the second half of the year [4,15]. After a gestation period of approximately 60 days, an average of five pups are born [4,15]. Raccoons living in the wild reach an age of only a few years (average, 3.1 years), but captive animals can live up to 17 years [16,17].
2. The Raccoons as a Neozoon
As a result of both unintentional and planned releases of raccoons outside their natural range, they are now found in parts of Europe and Asia, including Japan. As livestock for the fur industry, raccoons were kept in European fur farms as early as the early 20th century. Since 1934, here have been both planned and accidental releases of raccoons in Germany [18,19,20,21,22,23,24,25,26]. The following decades saw the spread of raccoons throughout major parts of Europe.
In the 1970s, raccoons entered Japan as pets. They were released by some pet owners and breeders and spread rapidly to almost the entire area of Japan [1].
Some characteristics of raccoons favor their invasive colonization of new habitats. Raccoons are highly adaptable and, as omnivores, can utilize a very broad spectrum of food, including anthropogenic sources. This feature is illustrated by their colonization of suburban and urban habitats. Raccoons are very good at defending themselves and have few predators, especially in newly colonized areas. Furthermore, they have a high reproductive rate [2,8]. Finally, raccoons have complex, sometimes sex-specific social behavior, which can also promote population density and habitat colonization. Raccoon pups can survive on their own from about four months of age. Female cubs often remain with their mothers, while male offspring migrate and colonize new territories [27]. Mothers and their female offspring usually form family groups, and many female raccoons remain near their mothers throughout their lives. The animals live in a common territory and have recurrent contact at sleeping and feeding sites. Young males, on the other hand, often form so-called coalitions in distant areas. A few unrelated males live in the same area and sometimes meet at sleeping and feeding sites, but without living as closely together as the female groups. This helps male raccoons defend themselves against strange males or other predators. However, in times of deprivation, such as severe winters, or during the mating season, contact between the female and male groups occurs regularly. In addition to female groups and male coalitions, solitary raccoons also occur. This is rather rare and seems to occur mainly in land areas with low raccoon population density and high resource availability. [27,28,29,30]. Population density and the intensity of social contacts depend primarily on the available resources. If sufficient food is available, there can be considerable overlapping of the territories of two raccoon groups and the number of contacts of the animals increases accordingly [18].
Raccoons prefer deciduous forests near water as habitats. Wetlands and riparian areas provide abundant food, and as omnivores, raccoons can use nearly any available trophic resource [3,31]. The availability of burrows in the ground or in plant cover for wintering and nesting is also important. [13,32,33]. Although raccoons are highly adaptable, there are environmental factors that prevent their expansion. Raccoons do not occur in deserts and boreal forests, and they cannot colonize habitats where the ground is covered with snow for 30% of the year or longer [34,35]. No raccoons have been sighted in high alpine mountains. In parts of the former Soviet Union, however, raccoons have been recorded in subalpine and alpine habitats up to 3500 m altitude [35]. The limiting factor in these extreme habitats is the permanent availability of food.
Young male raccoons spread out and can range over many hundreds of kilometers. This migration serves to expand the habitat of the species, promotes genetic variation, and prevents local overpopulation and resource depletion [36,37,38]. It is not only the extreme habitats mentioned above that pose insurmountable obstacles. Although raccoons often expand along rivers, very broad rivers and other geographic barriers may temporarily impede expansion [3,39]. However, there is no evidence that landscape barriers have caused a permanent blockade to expansion. There is even a report of a raccoon swimming across the Rhine River in southern Germany [40]. In addition, in recent decades there have been repeated releases of individual raccoons in Europe, often by private individuals, so that natural barriers are of little importance in suitable habitats [3,34,41,42].
3. The Distribution of Raccoons in Europe
Raccoons released in Central Germany in 1934 and in Northern Germany in the 1940s have long been considered the source of Central European raccoon populations [18,19,20,21,22,23,24,25,26]. There is now evidence that there was also a third release center in Eastern Germany [26]. Today, many decades later, these original populations still form the distribution focus of raccoons in Central Europe. It is difficult to determine the exact number of individuals in a population. However, official German hunting statistics suggest an extremely pronounced increase in the German raccoon population. While 3000 raccoons were killed in 1995, 100,000 animals killed were recorded two decades later [33]. Another analysis of hunting statistics from Germany determined the abundance index (the number of raccoons killed per hunting license), which takes into account the hunting effort. The data also demonstrate the very strong and almost exponential increase of the Central European raccoon population. While in 1995/96 there were about 100 killed animals per hunting license, in 2015/16 there were about 3400 killed animals per hunting license [34].
The descent of the Central European raccoon populations from the original release centers in Germany can be demonstrated today by population genetic analyses. In the North American habitat of origin, 76 different haplotypes were detected in 311 animals examined. In contrast, analyses of 193 individuals from Central Europe showed a comparatively low genetic diversity within the population, with only five different haplotypes [43,44]. Population genetic analyses have also supported the hypothesis that a third founding population was released in Eastern Germany [43,45]. A similar situation is found in Southern Europe. Spanish raccoon populations can also be traced back to a few individuals, according to population genetic studies [2,3,41].
The distribution of raccoons in Europe began with the release of raccoons in Germany in the 1930s and 1940s. At the same time, there were also individual official and private releases in other European countries, such as in the Soviet Union [35]. Selective individual releases of raccoons continue to have a role in the distribution of the species in Europe [3] and occur in parallel with the distribution of existing raccoon populations. Here, a distinction is made between unintentional releases (e.g., from fur farms and wildlife gardens) and planned releases (e.g., for hunting or by private livestock owners) [34].
Raccoons first appeared in Germany’s neighboring countries (Denmark, Poland, Czechia, Slovakia, Austria, Switzerland, France, Belgium, Netherlands, and Luxembourg). Subsequently, they spread further to the north (United Kingdom, Ireland, Norway, and Sweden), east (Lithuania, Ukraine, Russia, and Belarus), and south and southeast of Europe (Italy, Spain, Greece, Hungary, Slovenia, and Serbia) [3,31,34,35,42,46,47,48,49,50,51,52,53] (Figure 1).
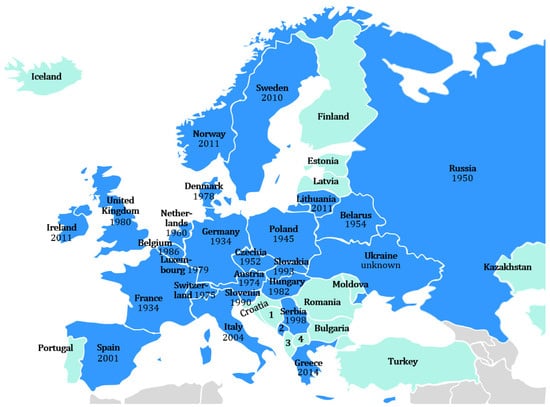
Figure 1.
European distribution of raccoons (blue: raccoons detected, green: no raccoons detected, gray: non-European country, not considered). A state is defined as a distribution area of raccoons, regardless of the abundance and distribution of the raccoons within the state territory. The year indicates the first detection of raccoons in a national territory. Abbreviations: 1: Bosnia and Herzegovina, 2: Montenegro (1998), data collected before the independence of Montenegro and therefore no separation between Serbia and Montenegro, 3: Albania, 4: Northern Macedonia [3,31,34,35,42,46,47,48,49,50,51,52,53].
In Central Europe, the original populations have increased and spread widely during the past nearly 100 years (Figure 2), mainly in Germany, Denmark, the Netherlands, Belgium, Luxembourg, and France. As a result, there are large areas with raccoon populations that are in contact with each other depending on geographic conditions. In Southern Europe (Southern France, Spain, and Italy), in contrast, there have been repeated individual releases [3,34,41,42], as well as in France very early in the last century. Due to this patchy release, raccoons already occur very widely in Southern Europe, although the main distribution area is far away. Thus, the animals often live in geographically separated populations. A similar situation can be found in Eastern and Southeastern Europe. Here, too, only isolated, geographically separated raccoon populations are found. In the literature, there are single records of animals that have not been deposited in the Global Biodiversity Information Facility (GBIF) database. It is therefore possible that some of the areas shown have not been permanently inhabited by the raccoon [34].
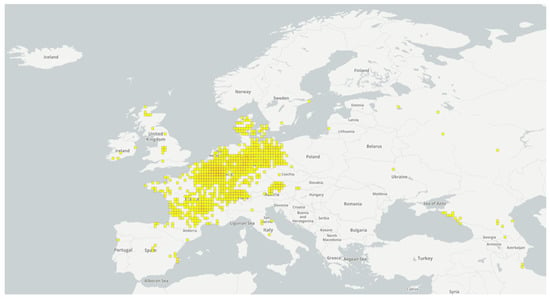
Figure 2.
Occurrence of the raccoon in Europe. Detection of animals from 2000 to 2022 (yellow dots). Analysis and visualization of d“Procyon lotor (Linnaeus, 1758)” using the Global Biodiversity Information Facility (GBIF) in November 2022 (https://www.gbif.org/species/5218786; accessed on 18 November 2022). Basis of records: observation, machine observation, human observation, material sample, material citation, and preserved specimen.
A special situation is found in the raccoon populations on the territory of the former Soviet Union. In the 20th century, raccoons were released into the wild on a large scale in a number of Soviet republics with the support of the government and with scientific assistance [35]. The animals originated from zoological gardens of the Soviet Union or, in a few cases, from raccoon farms in Western Europe. In addition, raccoons living in the wild were caught in live traps and released in other areas. Raccoons were released in Central Asia (Kyrgyzstan) as early as 1936. Further releases followed in the Far East (Asian part of Russia 1937), Eurasia (Azerbaijan 1941), Southern Russia (1950), and Central Asia (Uzbekistan 1953). In some cases, raccoons were released several times in one area or in closely located areas. From 1936 to 1958, a total of 1253 individuals were released throughout the former Soviet Union [35,54,55].
Currently, raccoons are frequently found in the European part of Russia, but there are no systematic studies on this issue. The Mammals of Russia portal (https://rusmam.ru/atlas/map; accessed on 18 November 2022) shows habitats in the more temperate climatic zones of Russia (Figure 3), e.g., at Lake Onega in the north, in the greater Moscow area, and in the east in the area of the city of Kazan. The main raccoon population is found on the southern edge of European Russia. Raccoons live along the Black Sea coast and throughout the Caucasus to the Caspian Sea. Counts from 1964 estimated about 25,000 animals in this region [35]. The high population density may be due to the fact that the Caucasus was a focal point of raccoon settlement in the former Soviet Union during the last century [36].
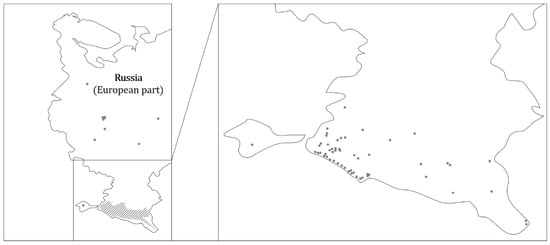
Figure 3.
Occurrence of the raccoon in the European part of Russia (dark grey dots). Raccoons are repeatedly spotted in European Russia (left). However, the main populations are located in the Caucasus and on the Black Sea coast of the extreme southern part of European Russia (right). Distribution data were obtained from the Mammals of Russia portal (https://rusmam.ru/atlas/map; accessed on 18 November 2022)) in November 2022.
Many neozoa have been intentionally or unintentionally introduced into the former Soviet Union. The potential ecological impacts of these neozoa on the environment have been tracked for a long time [56]. In the case of the raccoon, there is evidence that other species, e.g., ground-nesting birds, reptiles, and amphibians, have been predated or displaced regionally and seasonally. This has also been shown in studies in the rest of Europe. Other predators, such as the European mink (Mustela lutreola), may be displaced from their habitats by raccoons [2,57,58,59].
4. The Impact of Raccoons on European Habitats
One interaction of raccoons with their environment that can be particularly well perceived by humans is cohabitation in the same habitat and resulting contacts. In North America, raccoons are considered pests. The animals cause crop damage and can attack domestic and farm animals. Furthermore, damage and soiling of homes, equipment, and other facilities occurs with corresponding economic impacts [60,61,62]. A similar situation could be suspected of Europe, but there are hardly any systematic studies on this question. The city of Kassel (Germany), located in Central Europe, is one of the oldest hotspots of raccoon release in Europe. Accordingly, populations have been established in the city area and its surroundings. The city is believed to have the highest raccoon density of all European cities [63]. In towns, raccoons have a much lower escape distance from humans and other large animals. Together with the extensive supply of food and shelter, this makes the high density of raccoons possible. Nevertheless, wild raccoons living in suburban and urban habitats went unnoticed for a long time. It was not until the 1960s that wild raccoons were discovered in human settlements [63,64]. Even in densely populated urban centers, there are sufficient opportunities for raccoons to survive (parks, gardens, cemeteries). However, raccoon densities decrease from the urban fringe to the urban center, probably due to decreasing resources for survival [18,64]. In contrast to North America, the overall perception of urban raccoons in Europe does not appear to be solely negative. The animals are often even fed by residents in gardens or housing developments [32,63,65]. Over the decades, all attempts to control raccoon populations in urban areas have proven ineffective. Resettlement and various hunting options have ultimately been unsuccessful [18,34,63]. A very similar situation is found in other European habitats, although there is much less data available than for Central Europe [41,42,66].
Neozoa can also have impacts on the ecosystem through interaction with native animal species. Predator–prey interactions and competition for food and sleeping or nesting sites can cause ecological shifts to occur. This has been suggested for the raccoon as a medium-sized predator in Europe [31], but to date there are no robust data on this question [56,66,67,68,69]. There are no large predators in the European distribution range of raccoons that could limit the raccoon population. Comparable native predators are the European mink (Mustela lutreola), polecat (Mustela putorius), European badger (Meles meles), and raccoon dog (Nyctereutes procyonoides), which has also been released as a neozoon in Eastern Europe. Interactions of these predator species with raccoons have not been observed to date. The decrease in the European raccoon dog population is not due to competition with the raccoon, as was first suspected, but has been attributed to infection with canine distemper virus [69,70]. In North America, raccoons prey on ground- and cavity-nesting birds, especially in geographically isolated habitats, such as islands [71,72]. The ecological impact of raccoons in Europe has been poorly studied. The results of studies on the influence of raccoons on European ecosystems are difficult to interpret statistically because of their small number. Currently, the impact of raccoons on the native fauna is considered to be rather low.
5. Raccoons as Vectors of Zoonotic Diseases
One area of concern in interactions between neozoa and the environment is the transmission of pathogens. Because raccoons have strong populations in suburban and urban habitats, this issue affects not only other wildlife, but also livestock and humans [2]. In anthropogenic habitats, infectious diseases are among the most common causes of mortality in raccoons [8,14,73]. In extreme cases, neozoa can not only increase disease transmission, but also introduce new pathogens [74]. In their North American habitat, raccoons are intermediate and final hosts for a number of zoonotic pathogens that regularly lead to infections in humans [75,76,77,78,79]. However, a much lower spectrum of parasites and zoonotic pathogens has been detected in wild raccoons in Central Europe and Germany than in North American raccoons [80,81].
The raccoon roundworm (Baylisascaris procyonis) is the most common parasite of the raccoon. The raccoon is the final host of the nematode and excretes the worm eggs with its feces. These eggs, when ingested by humans, can cause serious and fatal infectious diseases. Particularly in urban habitats, a large number of nematode eggs can be released into the environment due to the high raccoon density. The eggs are extremely resistant to environmental influences and are infectious for a long time. In North America, the rate of infestation of raccoons with the raccoon roundworm is up to 80% and represents a corresponding risk of infection in humans [82,83]. In Europe, only a few cases of infectious diseases transmitted by raccoons have been described. In 1991, for example, one case was documented in which a raccoon keeper in Germany became sick due to a raccoon roundworm infection [84].
In addition, there are several other helminth species that parasitize in raccoons. Besides Baylisascaris procyonis, human pathogenic species are, for example, Trichinella species and Strongyloides procyonis, which has also been found in raccoons in Eastern Europe [31]. There are also raccoon pathogenic species from the protozoan group that can be transmitted to humans. These include members of the species Trypanosoma, Toxoplasma, Cryptosporidium, Neospora, and Sarcocystis, which have been detected sporadically in European raccoons [2,81,85].
Among the most important bacterial zoonotic pathogens in raccoons are two species, Leptospira and Salmonella. Bacteria of these groups are considered hazardous zoonotic pathogens worldwide, affecting a wide range of host animals [86,87]. In North America, a number of other bacterial species have been identified in raccoons, including Francisella, Mycobacterium, Borrelia, and Rickettsia [2,81]. The bacterial infection status of European raccoon populations has rarely been systematically studied. However, the few random analyses available suggest that the broad bacterial spectrum and sometimes high infestation of North American raccoons have not been transferred to the new habitats in Europe [81,88,89].
Viral pathogens in raccoons include the rabies virus. Rabies is a fatal and incurable zoonotic encephalitis that can affect almost all carnivores [90,91]. Often, the various animal pathogenic virus types can infect other animal species. In Europe, canine rabies is considered largely eliminated. However, rabies reservoirs exist in wild animals, for example, in foxes, raccoon dogs, and bats [92,93]. Rabies virus-positive raccoons have been reported sporadically in Central and Eastern Europe [2]. The causative agent of canine distemper, canine distemper virus, is also consistently documented in wildlife, including raccoons. Most canine distemper virus infections in raccoons are described at the end of the mating season and during the migration of the young male raccoons [94,95]. Viral strains isolated from Central and Southern European raccoons have all been assigned to the phylogenetic European group [96,97,98]. Several other viruses, such as parvoviruses, pseudorabies virus, and adenoviruses, have been detected in North American raccoon populations, but not yet in European populations [2].
Overall, the risk of infection with zoonotic pathogens from raccoons in Europe appears to be rather low. This may be due to the lower spectrum of pathogens in European raccoon populations. For risk assessment, however, the current scientific study situation in Europe must also be considered. There are fewer scientific analyses regarding the health and infection status of raccoons compared to research in North America. As is often the case in wildlife biology and wildlife medicine, this is due to the difficulty in accessing individuals in a population. In addition, the animals’ high local mobility and rapid distribution into new habitats makes systematic documentation of infestation rates challenging. Only random samples with usually low numbers of cases are available, from which extrapolations are made to populations with often high numbers of individuals, at least in Central Europe. For almost all of Europe, these surveys are not carried out systematically and over longer periods of time, but represent individual projects of single research institutions. Nevertheless, it seems advisable to monitor the potential risk of infection from wild raccoons. Systematic and, if favorable, Europe-wide programs to screen European raccoon populations for pathogens might help.
6. Outlook
Raccoons have been living in Europe for almost a century. Their numbers and the areas they inhabit within Europe are constantly increasing. In areas lacking in humans and close to nature, raccoons spread slowly. Here, contact with humans has a subordinate role. However, as with other predators (such as the wolf), conflicts can occur. In suburban and urban habitats, humans have become accustomed to the presence of raccoons and other urban wildlife, such as the fox, wild boar, and marten. Potential conflicts due to feeding damage or damage to facilities occur, but do not seem to prevent a predominantly consensual coexistence between humans and animals. Raccoons in densely populated urban areas have changed their behavior. They have reduced shyness and can coexist more closely with their own species. This, together with their adaptation to the wide anthropogenic food supply, enables the enormous population densities in urban habitats.
Management strategies for controlling the population size and dispersal potential of raccoons have failed so far. Hunting and wildlife biology programs, as well as relocation campaigns fail because of the high reproductive rate and the high mobility of the animals. Thus, it seems reasonable to accept raccoons as a neozoon and therefore as part of the European fauna. A feared negative impact on the overall native flora and fauna has not been observed so far, but regional studies show predation or displacement of native species. This ecological influence may depend on seasonal variation in food availability. The danger of zoonotic pathogens seems to have been overestimated. The pathogen spectrum of wild European raccoons is significantly smaller than that of North American raccoons. Accordingly, there have been few cases in Europe of zoonoses attributed to contact with raccoons. It is possible, however, that all of these effects depend on population density. Systematic monitoring of raccoon habitats, their population densities, and pathogen reservoirs therefore seems advisable. This would be all the more effective if carried out on a European level.
7. Conclusions
The raccoon has been spread throughout many parts of Europe by both accidental and planned releases. In this process, the raccoon’s adaptability favored its colonization and survival in new habitats. A high reproductive capacity and mobility, the absence of natural predators, and the use of a wide range of resources are the most striking characteristics of the raccoon. These characteristics even have culminated in the raccoon’s ability to colonize suburban and urban habitats with great success. Conflicts with endogenous fauna and humans are present, but are nowhere near as significant as in the raccoon’s original distribution range in North America. Programs to eradicate raccoons in parts of Europe have regularly failed. Thus, raccoons should be considered part of the European wildlife.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Ikeda, T.; Asano, M.; Matoba, Y.; Abe, G. Present status of invasive alien raccoon and its impact in Japan. Glob. Environ. Res. 2004, 8, 125–131. [Google Scholar]
- Beltran-Beck, B.; Garcia, F.J.; Gortazar, C. Raccoons in Europe: Disease hazards due to the establishment of an invasive species. Eur. J. Wildl. Res. 2012, 58, 5–15. [Google Scholar] [CrossRef]
- García, J.T.; García, F.J.; Alda, F.; González, J.L.; Aramburu, M.J.; Cortés, Y.; Prieto, B.; Pliego, B.; Pérez, M.; Herrera, J.; et al. Recent invasion and status of the raccoon (Procyon lotor) in Spain. Biol. Invasions 2012, 14, 1305–1310. [Google Scholar] [CrossRef]
- Lotze, J.H.; Anderson, S. Procyon lotor. Mamm. Species 1979, 119, 1–8. [Google Scholar] [CrossRef]
- Kays, R. Raccoons and relatives. In Walker’s Mammals of the World, 6th ed.; Nowak, R.M., Walker, E.P., Eds.; The John Hopkins University Press: Baltimore, MD, USA; London, UK, 1999; pp. 694–700. [Google Scholar]
- Pedlar, J.H.; Fahrig, L.; Merriam, H.G. Raccoon habitat use at 2 spatial scales. J. Wildl. Manag. 1997, 61, 105–112. [Google Scholar] [CrossRef]
- Hohmann, U.; Gerhard, R.; Kasper, M. Home range size of adult raccoons (Procyon lotor) in Germany. Z. Saugetierkd. 2000, 65, 124–127. [Google Scholar]
- Rosatte, R.C. Management of raccoons (Procyon lotor) in Ontario, Canada: Do human intervention and disease have significant impact on raccoon populations? Mammalia 2000, 64, 369–390. [Google Scholar] [CrossRef]
- Engelmann, A.; Michler, B.A.; Michler, F.U. Eine Frage der Saison—Aktuelle Ergebnisse zur Nahrungsökologie des Waschbären (Procyon lotor) in der nordostdeutschen Tiefebene. Labus 2012, 36, 47–62. [Google Scholar]
- Bozek, C.K.; Prange, S.; Gehrt, S.D. The influence of anthropogenic resources on multiscale habitat selection by raccoons. Urban Ecosyst. 2007, 10, 413–425. [Google Scholar] [CrossRef]
- Prange, S.; Gehrt, S.D.; Wiggers, E.P. Influences of anthropogenic resources on raccoon (Procyon lotor) movements and spatial distribution. J. Mammal. 2004, 85, 483–490. [Google Scholar] [CrossRef]
- Gehrt, S.D. Ecology and Management of Striped Skunks, Raccoons and Coyotes in Urban Landscapes. In People and Predators: From Conflict to Coexistence, 2nd ed.; ND Press: Washington, DC, USA, 2004; pp. 81–94. [Google Scholar]
- Köhnemann, B.A.; Michler, F.U. Sumpf- und Moorlandschaften der nordostdeutschen Tiefebene—Idealhabitate für Waschbären (Procyon lotor L., 1758) in Mitteleuropa? Beitr. Jagd. Wildforsch. 2009, 34, 511–524. [Google Scholar]
- Riley, S.P.D.; Hadidad, J.; Mansli, D.A. Population density, survival, and rabies in raccoons in an urban national park. Can. J. Zool. 1998, 76, 1153–1164. [Google Scholar] [CrossRef]
- Fritzell, E.K. Reproduction of raccoons (Procyon lotor) in North Dakota. Am. Midl. Nat. 1978, 100, 253–256. [Google Scholar] [CrossRef]
- Johnson, A.S. Biology of the raccoon (Procyon lotor varius Nelson and Goldman) in Alabama. Auburn Univ. Ala. Agric. Exp. Stn. Bull. 1970, 402, 1–148. [Google Scholar]
- Grau, G.A.; Sanderson, G.C.; Rogers, J.P. Age determination of raccoons. J. Wildl. Manag. 1970, 34, 364–372. [Google Scholar] [CrossRef]
- Hohmann, U.; Bartussek, I. Der Waschbär, 2nd ed.; Oertel und Spörer: Reutlingen, Germany, 2005. [Google Scholar]
- Wiegmann, A.F.A. Ueber die Gattung. Procyon. Arch. Nat. 1837, 3, 353–372. [Google Scholar]
- Franz, J.M. Die Einbürgerung von Säugetieren und Vögeln in Europa. Ergebnisse und Aussichten. Entomophaga 1963, 8, 374. [Google Scholar] [CrossRef]
- Leicht, E. Waschbär—Kleiner Feldversuch mit großer Wirkung. AFZ-DerWald 2009, 11, 570–573. [Google Scholar]
- Pagel, T.; Spieß, W. Der Zoologische Garten in Cöln eröffnet am 22. Juli 1860—150 Jahre Wildtierhaltung und -zucht. Zool. Gart. NF 2011, 80, 117–202. [Google Scholar] [CrossRef]
- Müller-Using, D. Die Ausbreitung des Waschbären (Procyon lotor [L.]) in Westdeutschland. Z. Jagdwiss. 1959, 5, 108–109. [Google Scholar] [CrossRef]
- Lutz, W. Occurrence and morphometrics of the raccoon Procyon lotor L. in Germany. Ann. Zool. Fenn. 1995, 32, 15–20. [Google Scholar]
- Frantz, A.C.; Cyriacks, P.; Schley, L. Spatial behaviour of a female raccoon (Procyon lotor) at the edge of the species’ European distribution range. Eur. J. Wildl Res. 2005, 51, 126–130. [Google Scholar] [CrossRef]
- Stubbe, M. Der Waschbär Procyon lotor (L., 1758) in der DDR. Hercynia. NF 1975, 12, 80–91. [Google Scholar]
- Bartussek, I. Die Waschbären Kommen, 1st ed.; Cognitio: Niedenstein, Germany, 2004. [Google Scholar]
- Gehrt, S.D. Raccoon Social Organization in South Texas. Ph.D. Thesis, University of Missouri, Missouri, MO, USA, 1994. [Google Scholar]
- Pitt, J.A.; Lariviere, S.; Messier, F. Social organization and group formation of raccoons at the edge of their distribuition. J. Mamm. 2008, 89, 646–653. [Google Scholar] [CrossRef]
- Muschik, I.; Köhnemann, B.A.; Michler, F.U. Untersuchungen zur Entwicklung des Raum und Sozialverhaltens von Waschbär-Mutterfamilien (Procyon lotor L.). Beitr. Jagd. Wildforsch. 2011, 36, 573–585. [Google Scholar]
- Bartoszewicz, M.; Okarma, H.; Zalwski, A.; Szczesna, J. Ecology of the raccoon (Procyon lotor) from western Poland. Ann. Zool. Fenn. 2008, 45, 291–298. [Google Scholar] [CrossRef]
- Michler, F.U.; Hohmann, U.; Stubbe, M. Aktionsräume, Tagesschlafplätze und Sozialsystem des Waschbären (Procyon lotor Linné 1758) im urbanen Lebensraum der Großstadt Kassel (Nordhessen). Beitr. Jagd. Wildforsch. 2004, 29, 57–273. [Google Scholar]
- Fischer, M.L.; Hochkirch, A.; Heddergott, M.; Schulze, C.; Anheyer-Behmenburg, H.E.; Lang, J.; Michler, F.U.; Hohmann, U.; Ansorge, H.; Hoffmann, L.; et al. Historical invasion records can be misleading: Genetic evidence for multiple introductions of invasive raccoons (Procyon lotor) in Germany. PLoS ONE 2015, 10, e0125441. [Google Scholar] [CrossRef]
- Salgado, I. Is the raccoon (Procyon lotor) out of control in Europe? Biodivers. Conserv. 2018, 27, 2243–2256. [Google Scholar] [CrossRef]
- Aliev, F.F.; Sanderson, G.C. Distribution and status of the raccoon in the Soviet Union. J. Wildl. Manag. 1966, 30, 497–502. [Google Scholar] [CrossRef]
- Michler, F.U.; Köhnemann, B.A. Tierische Spitzenleistung—Abwanderungsverhalten von Waschbären (Procyon lotor L., 1758) in Norddeutschland. Labus 2010, 31, 52–59. [Google Scholar]
- Lynch, G.M. Long-range movement of a raccoon in Manitoba. J. Mammal. 1967, 48, 659–660. [Google Scholar] [CrossRef]
- Priewert, A.R. Record of an extensive movement by a raccoon. J. Mammal. 1961, 42, 113. [Google Scholar] [CrossRef]
- Helbig, D. Untersuchungen zum Waschbären (Procyon lotor LINNE, 1758) im Raum Bernburg. Nat. Land Sachs. Anhalt. 2011, 48, 3–19. [Google Scholar]
- Cerutti, H. Der Immigrant. Neue Zürcher Zeitung. 2006, 1, 4. [Google Scholar]
- Alda, F.; Ruiz-López, M.J.; García, F.J.; Gompper, M.E.; Eggert, L.S.; Garcia, J.T. Genetic evidence for multiple introduction events of raccoons (Procyon lotor) in Spain. Biol. Invasions 2013, 15, 687–698. [Google Scholar] [CrossRef]
- Mori, E.; Mazza, G.; Menchetti, M.; Panzeri, M.; Gager, Y.; Bertolino, S.; Di Febbraro, M. The masked invader strikes again: The conquest of Italy by the Northern raccoon. Hystrix 2015, 26, 47–51. [Google Scholar]
- Frantz, A.C.; Heddergott, M.; Lang, J.; Schulze, C.; Ansorge, H.; Runge, M.; Braune, S.; Michler, F.U.; Wittstatt, U.; Hoffmann, L.; et al. Limited mitochondrial DNA diversity is indicative of a small number of founders of the German raccoon (Procyon lotor) population. Eur. J. Wildl. Res. 2013, 59, 665–674. [Google Scholar] [CrossRef]
- Cullingham, C.I.; Kyle, C.J.; Pond, B.A.; White, B.N. Genetic structure of raccoons in eastern North America based on mtDNA: Implications for subspecies designation and rabies disease dynamics. Can. J. Zool. 2008, 86, 947–958. [Google Scholar] [CrossRef]
- Gramlich, S.; Schulz, H.; Köhnemann, B.A.; Michler, F.U. Mater semper certa?—Molekularbiologische Analyse einer Waschbärenpopulation (Procyon lotor Linné, 1758) im Müritz-Nationalpark. Beitr. Jagd. Wildforsch. 2011, 36, 521–530. [Google Scholar]
- Broekhuizen, S.; Müskens, G.J.D.M.; Niewold, F.J.J.; Thissen, J.B.M. Heimkehrer und Neubürger unter den Säugetieren der Niederlande im 19. und 20. Jahrhundert. Beitr. Jagd. Wildforsch. 2001, 26, 155–170. [Google Scholar]
- Schlei, L.; Schank, C.; Schaul, M.; Sinner, C. Neubürger und Heimkehrer unter den Wildtieren Luxenburgs. Beitr. Jagd. Wildforsch. 2001, 26, 141–154. [Google Scholar]
- Léger, F. Répartition en France de trois petits carnivores introduits. Beitr. Jagd. Wildforsch. 2001, 26, 137–139. [Google Scholar]
- Heltai, M.; Szemethy, L.; Lanszky, J.; Csanyi, S. Returning and new mammal predators in Hungary: The status and distribution of the golden jackal (Canis aureus), raccoon dog (Nyctereutes procyonides) and raccoon (Procyon lotor) in 1997-2000. Beitr. Jagd. Wildforsch. 2000, 26, 95–102. [Google Scholar]
- Spitenberger, F.; Bauer, K.; Sackl, P.; Sieber, J. Heimkehrer und Neubürger der österreichischen Säugertierfauna. Beitr. Jagd. Wildforsch. 2001, 26, 127–136. [Google Scholar]
- Canova, L.; Rossi, S. First records of the northern raccoon Procyon lotor in Italy. Hystrix. It. J. Mamm. 2008, 19, 179–182. [Google Scholar]
- Zimina, R.P. The main features of the Caucasian natural landscapes and their conservation, USSR. Artic. Alp. Res. 1978, 10, 479–488. [Google Scholar] [CrossRef]
- Stille, M.; Gasteratos, I.; Stille, B. Alien and invasive terrestrial vertebrate species on Corfu, Ionian Islands, Greece. J. Vertebr. Biol. 2021, 70, 20126. [Google Scholar] [CrossRef]
- Aliev, F.F. Acclimatization and the economic acclimatization of the raccoon in Azerbaijan. Acad. Sci. Azerbaijan SSR Rept. 1955, 8, 571–578. [Google Scholar]
- Redford, P. Raccoon in the U.S.S.R. J. Mammal. 1962, 43, 541–542. [Google Scholar] [CrossRef]
- Khlyap, L.A.; Bobrov, V.V.; Warshavsky, A.A. Biological invasions on Russian territory: Mammals. Russ. J. Biol. Invasions 2010, 1, 127–140. [Google Scholar] [CrossRef]
- Kauhala, K. Introduced carnivores in Europe with special reference to central and northern Europe. Wildl. Biol. 1996, 2, 197–204. [Google Scholar] [CrossRef]
- Kudaktin, A.N.; Romas, A.V. Raccon in the Russian Black Sea Coast. Biota Environ. 2019, 2, 88–103. [Google Scholar]
- Langgemach, T.; Bellebaum, J. Prädation und der Schutz bodenbrütender Vogelarten in Deutschland. Vogelwelt 2005, 126, 259–298. [Google Scholar]
- Gehrt, S.D.; Fritzell, E.K. Resource distribution, female home range dispersion and male spatial interactions: Group structure in a solitary carnivore. Anim. Behav. 1998, 55, 1211–1227. [Google Scholar] [CrossRef]
- Miller, C.A.; Campbell, L.K.; Yeagle, J.A. Attitudes of Homeowners in the Greater Chicago Metropolitan Region Toward Nuisance Wildlife; Program Report SR-00-02; Illinois Natural History Survey: Champaign, IL, USA, 2001. [Google Scholar]
- Beasly, J.C.; Rhodes, O.E. Relationship between raccoon abundance and crop damage. Hum. Wildl. Confl. 2006, 2, 248–259. [Google Scholar]
- Michler, F.U. Untersuchungen zur Raumnutzung des Waschbären (Procyon lotor, L. 1758) im urbanen Lebensraum am Beispiel der Stadt Kassel (Nordhessen). Diploma Thesis, Halle-Wittenberg University, Halle, Germany, 2003. [Google Scholar]
- Hohmann, U.; Voigt, S.; Andreas, U. Quo vadis raccoon? New visitors in our backyards—On the urbanization of an allochthone carnivore in Germany. Nat. Verhalt. 2001, 2, 143–148. [Google Scholar]
- Jernelöv, A. Raccoons in Europe (Germany). In The Long-Term Fate of Invasive Species: Aliens Forever or Integrated Immigrants with time? 1st ed.; Jernelöv, A., Ed.; Springer: Basel, Switzerland, 2017; pp. 217–230. [Google Scholar]
- Boscherini, A.; Mazza, G.; Menchetti, M.; Laurenzi, A.; Mori, E. Time is running out! Rapid range expansion of the invasive northern raccoon in central Italy. Mammalia 2020, 84, 98–101. [Google Scholar] [CrossRef]
- Lutz, W. The introduced raccoon Procyon lotor population in Germany. Wildl. Biol. 1996, 2, 228. [Google Scholar] [CrossRef]
- Michler, F.U.; Michler, B.A. Ökologische, ökonomische und epidemiologische Bedeutung des Waschbären (Procyon lotor) in Deutschland—Eine aktuelle Übersicht. Beitr. Jagd. Wildforsch. 2012, 37, 385–397. [Google Scholar]
- Mulder, J.L. A review of the ecology of the raccoon d.dog (Nyctereutes procyonoides) in Europe. Lutra 2012, 55, 101–127. [Google Scholar]
- Mulder, J.L. The raccoon dog (Nyctereutes procyonoides) in the Netherlands-its present status and a risk assessment. Lutra 2013, 56, 23–43. [Google Scholar]
- Davis, M.B.; Simons, T.R.; Groom, M.J.; Weaver, J.L.; Cordes, J.R. The breeding status of the American oystercatcher on the east coast of North America and breeding success in North Carolina. Waterbirds 2001, 24, 195–202. [Google Scholar] [CrossRef]
- Erwin, R.M.; Truitt, B.R.; Jiménez, J.E. Ground-nesting waterbirds and mammalian carnivores on the Virginia barrier island region: Running out of options. J. Coast. Res. 2001, 17, 292–296. [Google Scholar]
- Hadidian, J.; Prange, S.; Rosatte, R.; Riley, S.P.D.; Gehrt, S.D. Raccoons (Procyon lotor). In Urban Carnivores. Ecology, Conflict, and Conservation, 1st ed.; Clark, H.O., Ed.; The John Hopkins University Press: Baltimore, MD, USA; London, UK, 2010; pp. 35–47. [Google Scholar]
- Manchester, S.J.; Bullock, J.M. The impacts of non-native species on UK biodiversityand the effectiveness of control. J. Appl Ecol 2000, 37, 845–864. [Google Scholar] [CrossRef]
- Galton, M.M. The epidemiology of leptospirosis in the United States. Public Health Rep. 1896, 74, 141–148. [Google Scholar] [CrossRef]
- Orihel, T.C.; Eberhard, M.L. Zoonotic filariasis. Clin. Microbiol Rev. 1998, 11, 366–381. [Google Scholar] [CrossRef]
- Fried, B.; Abruzzi, A. Food-borne trematode infections of humans in the United States of America. Parasitol. Res. 2010, 106, 1263–1280. [Google Scholar] [CrossRef]
- Mackenstedt, U.; Jenkins, D.; Romig, T. The role of wildlife in the transmission of parasitic zoonoses in peri-urban and urban areas. Int. J. Parasitol. Parasites Wildl. 2015, 4, 71–79. [Google Scholar] [CrossRef]
- Weinstein, S.B. Baylisascaris procyonis demography and egg production in a California raccoon population. J. Parasitol. 2016, 102, 622–628. [Google Scholar] [CrossRef]
- Schwarz, S.; Sutor, A.; Mattis, R.; Conraths, F.J. Der Waschbärspulwurm (Baylisascaris procyonis)—Kein Zoonoserisiko für Brandenburg? Berl. Münchener Tierärztl. Wochenschr. 2015, 128, 34–38. [Google Scholar]
- Stope, M. Wild life raccoons in Germany as a reservoir for zoonotic agents. Eur. J. Wildl Res. 2019, 65, 94. [Google Scholar] [CrossRef]
- Sorvillo, F.; Ash, L.R.; Berlin, O.G.W.; Morse, S.A. Baylisascaris procyonis: An emerging helminthic zoonosis. Emerg. Infect. Dis. 2002, 8, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Kazakos, K.R. Baylisascaris procyonis and related species. In Parasitic Diseases of Wild Mammals, 2nd ed.; Samuel, W.M., Pybus, M.J., Kocan, A.A., Eds.; Iowa State University: Ames, IA, USA, 2001; pp. 301–341. [Google Scholar]
- Küchle, M.; Knorr, H.L.; Medenblik-Frysch, S.; Weber, A.; Bauer, C.; Naumann, G.O. Diffuse unilateral subacute neuroretinitis syndrome in a German most likely caused by the raccoon roundworm, Baylisascaris procyonis. Graefes Arch. Clin. Exp. Ophthalmol. 1993, 231, 48–51. [Google Scholar] [CrossRef]
- Stolte, M.; Odening, K.; Walter, G.; Bockhardt, I. The raccoon as intermediate host of three Sarcocystis species in Europe. Comp. Parasitol. 1996, 63, 145–149. [Google Scholar]
- Bharti, A.R.; Nally, J.E.; Ricaldi, J.N.; Matthias, M.A.; Diaz, M.M.; Lovett, M.A.; Levett, P.N.; Gilman, R.H.; Willig, M.R.; Gotuzzo, E.; et al. Leptospirosis: A zoonotic disease of global importance. Lancet Infect. Dis. 2003, 3, 757–771. [Google Scholar] [CrossRef]
- Morse, E.V.; Midla, D.A.; Kazakos, K.R. Raccoons (Procyon lotor L.) as carriers of Salmonella. J. Environ. Sci. Health A 1983, 18, 541–560. [Google Scholar]
- Hildebrand, J.; Bunkowska-Gawlik, K.; Adamczyk, M.; Gajda, E.; Merta, D.; Popiolek, M.; Perec-Matysiak, A. The occurrence of Anaplasmataceae in European populations of invasive carnivores. Ticks Tick Borne Dis. 2018, 9, 934–937. [Google Scholar] [CrossRef]
- Cybulska, A.; Skopek, R.; Kornacka, A.; Popiolek, M.; Pirog, A.; Laskowski, Z.; Moskwa, B. First detection of Trichinella pseudospiralis infection in raccoon (Procyon lotor) in Central Europe. Vet. Parasitol. 2018, 254, 114–119. [Google Scholar] [CrossRef]
- Bourhy, H.; Reynes, J.M.; Dunham, E.J.; Dacheux, L.; Larrous, F.; Huong, V.T.Q.; Xu, G.; Yan, J.; Miranda, M.E.G.; Holmes, E.C. The origin and phylogeography of dog rabies virus. J. Gen. Virol. 2008, 89, 2673–2681. [Google Scholar] [CrossRef]
- Faber, M.; Dietzschold, B.; Li, J. Immunogenicity and safety of recombinant rabies viruses used for oral vaccination of stray dogs and wildlife. Zoonoses Public Health 2009, 56, 262–269. [Google Scholar] [CrossRef]
- Holmala, K.; Kauhala, K. Habitat use of medium-sized carnivores in southeast Finland-key habitats for rabies spread? Ann. Zool. Fenn. 2009, 46, 233–246. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Botvinkin, A.D.; McElhinney, L.M.; Smith, J.S.; Orciari, L.A.; Hughes, G.J.; Fooks, A.R.; Rupprecht, C.E. Molecular epidemiology of terrestrial rabies in the former Soviet Union. J. Wildl. Dis. 2004, 40, 617–631. [Google Scholar] [CrossRef]
- Nakano, H.; Kameo, Y.; Sato, H.; Mochizuki, M.; Yokoyama, M.; Uni, S.; Shibasaki, T.; Maeda, K. Detection of antibody to canine distemper virus in wild raccoons (Procyon lotor) in Japan. J. Vet. Med. Sci. 2009, 71, 1661–1663. [Google Scholar] [CrossRef]
- Roscoe, D.E. Epizootiology of canine distemper in New Jersey raccoons. J. Wildl. Dis. 1993, 29, 390–395. [Google Scholar] [CrossRef]
- Nikolin, V.M.; Wibbelt, G.; Michler, F.U.; Wolf, P.; East, M.L. Susceptibility of carnivore hosts to strains of canine distemper virus from distinct genetic lineages. Vet. Microbiol. 2012, 156, 45–53. [Google Scholar] [CrossRef]
- Benetka, V.; Leschnik, M.; Affenzeller, N.; Mostl, K. Phylogenetic analysis of Austrian canine distemper virus strains from clinical samples from dogs and wild carnivores. Vet. Rec. 2011, 168, 377. [Google Scholar] [CrossRef]
- Michler, F.U.; Köhnemann, B.A.; Roth, M.; Speck, S.; Fickel, J.; Wibbelt, G. Todesursache sendermarkierter Waschbären (Procyon lotor L., 1758) im Müritz-Nationalpark (Mecklenburg-Vorpommern). Beiträge Jagd. Wildforsch. 2009, 34, 339–355. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).