Phenotypic and Genetic Study of the Presence of Hair Whorls in Pura Raza Español Horses

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Description of the Traits and Database
2.2. Statistical Analyses
2.3. Genetic Model
3. Results
3.1. Statistical Analysis
3.2. Genetic Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abdel-Azeem, N.M.; Emeash, H.H. Relationship of horse temperament with breed, age, sex, and body characteristics: A questionnaire-based study. Beni-Suef Univ. J. Basic Appl. Sci. 2021, 10, 45. [Google Scholar] [CrossRef]
- Finn, J.L.; Haase, B.; Willet, C.E.; van Rooy, D.; Chew, T.; Wade, C.M.; Hamilton, N.A.; Velie, B.D. The relationship between coat colour phenotype and equine behaviour: A pilot study. Appl. Anim. Behav. Sci. 2016, 174, 66–69. [Google Scholar] [CrossRef]
- Bartolomé, E.; Perdomo-González, D.I.; Ripollés-Lobo, M.; Valera, M. Basal Reactivity Evaluated by Infrared Thermography in the “Caballo de Deporte Español” Horse Breed According to Its Coat Color. Animals 2022, 12, 2515. [Google Scholar] [CrossRef]
- Brunberg, E.; Gille, S.; Mikko, S.; Lindgren, G.; Keeling, L.J. Icelandic horses with the Silver coat colour show altered behaviour in a fear reaction test. Appl. Anim. Behav. Sci. 2013, 146, 72–78. [Google Scholar] [CrossRef]
- Grandin, T.; Deesing, M.J.; Struthers, J.J.; Swinker, A.M. Cattle with hair whorl patterns above the eyes are more behaviorally agitated during restraint. Appl. Anim. Behav. Sci. 1995, 46, 117–123. [Google Scholar] [CrossRef]
- King, T.; Marston, L.C.; Bennett, P.C. Breeding dogs for beauty and behaviour: Why scientists need to do more to develop valid and reliable behaviour assessments for dogs kept as companions. Appl. Anim. Behav. Sci. 2012, 137, 1–12. [Google Scholar] [CrossRef]
- Preston, B.T.; Stevenson, I.R.; Lincoln, G.A.; Monfort, S.L.; Pilkington, J.G.; Wilson, K. Testes size, testosterone production and reproductive behaviour in a natural mammalian mating system. J. Anim. Ecol. 2012, 81, 296–305. [Google Scholar] [CrossRef]
- Topczewska, J.; Krupa, W.; Sokołowicz, Z.; Lechowska, J. Does Experience Make Hucul Horses More Resistant to Stress? A Pilot Study. Animals 2021, 11, 3345. [Google Scholar] [CrossRef]
- Górecka, A.; Golonka, M.; Chruszczewski, M.; Jezierski, T. A note on behaviour and heart rate in horses differing in facial hair whorl. Appl. Anim. Behav. Sci. 2007, 105, 244–248. [Google Scholar] [CrossRef]
- Lanier, J.L.; Grandin, T.; Green, R.; Avery, D.; McGee, K. A note on hair whorl position and cattle temperament in the auction ring. Appl. Anim. Behav. Sci. 2001, 73, 93–101. [Google Scholar] [CrossRef]
- Murphy, J.; Arkins, S. Facial hair whorls (trichoglyphs) and the incidence of motor laterality in the horse. Behav. Processes 2008, 79, 7–12. [Google Scholar] [CrossRef]
- Shivley, C.; Grandin, T.; Deesing, M. Behavioral Laterality and Facial Hair Whorls in Horses. J. Equine Vet. Sci. 2016, 44, 62–66. [Google Scholar] [CrossRef]
- Smith, D.W.; Gong, B.T. Scalp hair patterning as a clue to early fetal brain development. J. Pediatr. 1973, 83, 374–380. [Google Scholar] [CrossRef]
- Furdon, S.A.; Clark, D.A. Scalp hair characteristics in the newborn infant. Adv. Neonatal Care 2003, 3, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Broucek, J.; Kisac, P.; Mihina, S.; Hanus, A.; Uhrincat, M.; Tancin, V. Hair whorls of Holstein Friesian heifers and affects on growth and behaviour. Arch. Anim. Breed. 2007, 50, 374–380. [Google Scholar] [CrossRef]
- Leliveld, L.M.C.; Langbein, J.; Puppe, B. The emergence of emotional lateralization: Evidence in non-human vertebrates and implications for farm animals. Appl. Anim. Behav. Sci. 2013, 145, 1–14. [Google Scholar]
- Tomkins, L.M.; Williams, K.A.; Thomson, P.C.; McGreevy, P.D. Lateralization in the domestic dog (Canis familiaris): Relationships between structural, motor, and sensory laterality. J. Vet. Behav. 2012, 7, 70–79. [Google Scholar] [CrossRef]
- Divided Brains. Available online: https://www.cambridge.org/core/books/divided-brains/442F89DBA8F4BA76CD2F896E960A1E36 (accessed on 29 May 2023).
- Frasnelli, E.; Vallortigara, G.; Rogers, L.J. Left-right asymmetries of behaviour and nervous system in invertebrates. Neurosci. Biobehav. Rev. 2012, 36, 1273–1291. [Google Scholar] [CrossRef]
- Rogers, L.J.; Vallortigara, G.; Andrew, R.J. Divided Brains: The Biology and Behaviour of Brain Asymmetries; Cambridge University Press: Cambridge, UK, 2011; pp. 1–229. [Google Scholar]
- Rogers, L.J.; Andrew, R. (Eds.) Comparative Vertebrate Lateralization; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Hudayl, I. Gala de Caballeros Blason de Paladines; Editora Nacional: Sao Paulo, Brazil, 1977; ISBN 9788427604087. [Google Scholar]
- Aguilar, P. De Tratado de La Caballería de La Gineta [Manuscrito]/Compuesto Y Ordenado por El Capitán Pedro de Aguilar, Vecino de Málaga, Natural de La Ciudad de Antequera; Iuan René: Málaga, Spain, 1572. [Google Scholar]
- Suë, E. Arabian Godolphin; Derrydale Press: New York, NY, USA, 2003. [Google Scholar]
- Beatriz, A.; Torres, A. Avaliação da Reatividade de Equinos Diferenciados Pela Posição de Redemoinho Facial. Ph.D. Thesis, Universidade Federal de Santa Catarina, Florianópolis, Brazil, 2018. [Google Scholar]
- Lima, D.F.P.d.A.; da Cruz, V.A.R.; Pereira, G.L.; Curi, R.A.; Costa, R.B.; de Camargo, G.M.F. Genomic regions associated with the position and number of hair whorls in horses. Animals 2021, 11, 2925. [Google Scholar] [CrossRef]
- Cruz, V.A.R.; Lima, D.F.P.d.A.; Diaz, I.D.P.S.; Curi, R.A.; Pereira, G.L.; Costa, R.B.; de Camargo, G.M.F. Genetic parameters for hair whorl traits in horses. Livest. Sci. 2021, 252, 104679. [Google Scholar] [CrossRef]
- Górecka, A.; Słoniewski, K.; Golonka, M.; Jaworski, Z.; Jezierski, T. Heritability of hair whorl position on the forehead in Konik horses. J. Anim. Breed. Genet. 2006, 123, 396–398. [Google Scholar] [CrossRef]
- Yokomori, T.; Tozaki, T.; Mita, H.; Miyake, T.; Kakoi, H.; Kobayashi, Y.; Kusano, K.; Itou, T. Heritability estimates of the position and number of facial hair whorls in Thoroughbred horses. BMC Res. Notes 2019, 12, 346. [Google Scholar] [CrossRef] [PubMed]
- Mcbane, S.; Cooper, H. Caballos del Mundo; Hispano Europea Editorial S.A.: Barcelona, Spain, 1991; ISBN 9788425508752. [Google Scholar]
- Valera Córdoba, M. La Mejora Genética Equina en La Era de Las Ómicas: Una Historia Compartida: Lección Inaugural Leída en La Solemne Apertura del Curso Académico 2017–2018 en La Universidad de Sevilla; Editorial Universidad de Sevilla: Sevilla, Spain, 2017. [Google Scholar]
- Internationale, E. Fédération Equestre Internationale Identification of Horses with the Narrative and the Diagram; Fédération Equestre Internationale: Lausanne, Switzerland, 2007; pp. 1–19. [Google Scholar]
- Poyato-Bonilla, J.; Sánchez-Guerrero, M.J.; Santos, R.D.; Valera, M. Population study of the Pura Raza Español Horse regarding its coat colour. Ann. Anim. Sci. 2018, 18, 723–739. [Google Scholar] [CrossRef]
- Sánchez-Guerrero, M.J.; Molina, A.; Gómez, M.D.; Peña, F.; Valera, M. Relationship between morphology and performance: Signature of mass-selection in Pura Raza Español horse. Livest. Sci. 2016, 185, 148–155. [Google Scholar] [CrossRef]
- Ripollés-Lobo, M.; Perdomo-González, D.I.; Azor, P.J.; Valera, M. Evaluation of potential effects and genetic parameters in conformational limb defects in Pura Raza Española horses. Ital. J. Anim. Sci. 2023, 22, 407–417. [Google Scholar] [CrossRef]
- Stat Soft Statistica software v. 11.0. Available online: https://www.statistics.com/en/(accessed on 12 September 2023).
- Misztal, I.; Lourenco, D.; Legarra, A. Current status of genomic evaluation. J. Anim. Sci. 2020, 98, skaa101. [Google Scholar] [CrossRef]
- Plummer, M.; Best, N.; Cowles, K.; Vines, K. {CODA}: Convergence Diagnosis and Output Analysis for {MCMC}. R News 2006, 6, 7–11. [Google Scholar]
- Gutiérrez, J.P.; Goyache, F. A note on ENDOG: A computer program for analysing pedigree information. J. Anim. Breed. Genet. 2005, 122, 172–176. [Google Scholar] [CrossRef]
- Sobrino, I.M.; Ramón García, M. Identificación equina II. ExtremaduraPRE 2016, 25, 67–77. [Google Scholar]
- Arita, H.T. El regreso del caballo: Lo macro y lo micro en la evolución. Ciencias 2010, 97, 46–55. [Google Scholar]
- Olmos, G.; Turner, S.P. The relationships between temperament during routine handling tasks, weight gain and facial hair whorl position in frequently handled beef cattle. Appl. Anim. Behav. Sci. 2008, 115, 25–36. [Google Scholar] [CrossRef]
- Randle, H.D. Facial hair whorl position and temperament in cattle. Appl. Anim. Behav. Sci. 1998, 56, 139–147. [Google Scholar] [CrossRef]
- Samlaska, C.P.; James, W.D.; Sperling, L.C. Scalp whorls. J. Am. Acad. Dermatol. 1989, 21, 553–556. [Google Scholar] [CrossRef]
- Lloyd, A.S.; Martin, J.E.; Bornett-Gauci, H.L.I.; Wilkinson, R.G. Horse personality: Variation between breeds. Appl. Anim. Behav. Sci. 2008, 112, 369–383. [Google Scholar] [CrossRef]
- Burattini, B.; Fenner, K.; Anzulewicz, A.; Romness, N.; McKenzie, J.; Wilson, B.; McGreevy, P. Age-related changes in the behaviour of domestic horses as reported by owners. Animals 2020, 10, 2321. [Google Scholar] [CrossRef] [PubMed]
- Negro, S.; Sánchez-Guerrero, M.J.; Bartolomé, E.; Solé, M.; Gómez, M.D.; Membrillo, A.; Molina, A.; Valera, M. Evidence for the effect of serotoninergic and dopaminergic gene variants on stress levels in horses participating in dressage and harness racing. Anim. Prod. Sci. 2019, 59, 2206–2211. [Google Scholar] [CrossRef]
- Sánchez, M.J.; Bartolomé, E.; Valera, M. Genetic study of stress assessed with infrared thermography during dressage competitions in the Pura Raza Español horse. Appl. Anim. Behav. Sci. 2016, 174, 58–65. [Google Scholar] [CrossRef]
- Jacobs, L.N.; Staiger, E.A.; Albright, J.D.; Brooks, S.A. The MC1R and ASIP coat color loci may impact behavior in the horse. J. Hered. 2016, 107, 214–219. [Google Scholar] [CrossRef]
- Pérez-Guisado, J.; Lopez-Rodríguez, R.; Muñoz-Serrano, A. Heritability of dominant-aggressive behaviour in English Cocker Spaniels. Appl. Anim. Behav. Sci. 2006, 100, 219–227. [Google Scholar] [CrossRef]
- Reissmann, M.; Ludwig, A. Pleiotropic effects of coat colour-associated mutations in humans, mice and other mammals. Semin. Cell Dev. Biol. 2013, 24, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Martín, M.; Rodríguez-García, A.; Pérez-Losada, J.; Sagrera, A.; Read, A.P.; Sánchez-García, I. SLUG (SNAI2) deletions in patients with Waardenburg disease. Hum. Mol. Genet. 2002, 11, 3231–3236. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Losada, J.; Sánchez-Martin, M.; Rodríguez-García, A.; Sánchez, M.L.; Orfao, A.; Flores, T.; Sánchez-García, I. Zinc-finger transcription factor Slug contributes to the function of the stem cell factor c-kit signaling pathway. Blood 2002, 100, 1274–1286. [Google Scholar] [CrossRef] [PubMed]
- Brend, T.; Holley, S.A. Balancing segmentation and laterality during vertebrate development. Semin. Cell Dev. Biol. 2009, 20, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Pourquié, O. Segmentation of the vertebrate spine: From clock to scoliosis. Cell 2011, 145, 650. [Google Scholar] [CrossRef]
- Mintz, B. Gene control of mammalian differentiation. Annu. Rev. Genet. 1974, 8, 411–470. [Google Scholar] [CrossRef]
- de Zarate, J.C.O.; de Zarate, C.O.O. Hair whorl and handedness. Brain Cogn. 1991, 16, 228–230. [Google Scholar] [CrossRef]
- Sánchez, M.J.; Azor, P.J.; Molina, A.; Parkin, T.; Rivero, J.L.L.; Valera, M. Prevalence, risk factors and genetic parameters of cresty neck in Pura Raza Español horses. Equine Vet. J. 2017, 49, 196–200. [Google Scholar] [CrossRef]
- Ripolles, M.; Sánchez-Guerrero, M.J.; Perdomo-González, D.I.; Azor, P.; Valera, M. Survey of risk factors and genetic characterization of ewe neck in a world population of pura raza español horses. Animals 2020, 10, 1789. [Google Scholar] [CrossRef]
- Sánchez-Guerrero, M.J.; Solé, M.; Azor, P.J.; Sölkner, J.; Valera, M. Genetic and environmental risk factors for vitiligo and melanoma in Pura Raza Español horses. Equine Vet. J. 2019, 51, 606–611. [Google Scholar] [CrossRef]
- Sánchez-Guerrero, M.J.; Ramos, J.; Valdés, M.; Rivero, J.L.L.; Valera, M. Prevalence, environmental risk factors and heritability of body condition in Pura Raza Español horses. Livest. Sci. 2019, 230, 103851. [Google Scholar] [CrossRef]
- Poyato-Bonilla, J.; Perdomo-González, D.I.; Sánchez-Guerrero, M.J.; Varona, L.; Molina, A.; Casellas, J.; Valera, M. Genetic inbreeding depression load for morphological traits and defects in the Pura Raza Española horse. Genet. Sel. Evol. 2020, 52, 62. [Google Scholar] [CrossRef] [PubMed]
- Gómez, M.D.; Sánchez, M.J.; Bartolomé, E.; Cervantes, I.; Poyato-Bonilla, J.; Demyda-Peyrás, S.; Valera, M. Phenotypic and genetic analysis of reproductive traits in horse populations with different breeding purposes. Animal 2020, 14, 1351–1361. [Google Scholar] [CrossRef] [PubMed]
- Sánchez Guerrero, M.J.; Cervantes, I.; Valera, M.; Gutiérrez, J.P. Modelling genetic evaluation for dressage in pura raza español horses with focus on the rider effect. J. Anim. Breed. Genet. 2014, 131, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Wickens, C.; Brooks, S.A. Genetics of Equine Behavioral Traits. Vet. Clin. North Am.-Equine Pract. 2020, 36, 411–424. [Google Scholar] [CrossRef]
- Kim, N.Y.; Son, J.K.; Cho, I.C.; Shin, S.M.; Park, S.H.; Seong, P.N.; Woo, J.H.; Park, N.G.; Park, H.B. Estimation of genetic parameters for temperament in Jeju crossbred horses. Asian-Australasian J. Anim. Sci. 2018, 31, 1098–1102. [Google Scholar] [CrossRef]
- Hemmann, K.; Raekallio, M.; Vainio, O.; Juga, J. Crib-biting and its heritability in Finnhorses. Appl. Anim. Behav. Sci. 2014, 156, 37–43. [Google Scholar] [CrossRef]
- Lamac, N.; Solkner, J.; Meszaros, G. Analysis of excessive Homozygous regions in Rhodesian ridgeback dogs. Agric. Conspec. Sci. 2017, 82, 277–280. [Google Scholar]
- Salmon Hillbertz, N.H.C.; Isaksson, M.; Karlsson, E.K.; Hellmén, E.; Pielberg, G.R.; Savolainen, P.; Wade, C.M.; Von Euler, H.; Gustafson, U.; Hedhammar, Å.; et al. Duplication of FGF3, FGF4, FGF19 and ORAOV1 causes hair ridge and predisposition to dermoid sinus in Ridgeback dogs. Nat. Genet. 2007, 39, 1318–1320. [Google Scholar] [CrossRef]


{kind=link}
| Location | Position | Number | ||
|---|---|---|---|---|
| Head | 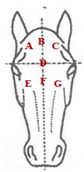 | Circular whorls | Linear whorls | |
| A: top right | Class 1: 1 | Class 1: 1 | ||
| B: top center | Class 2: 2 | Class 2: 2 | ||
| C: top left | Class 3: 3 | Class 3: ≥3 | ||
| D: between the eyes | Class 4: 4 | |||
| E: lower right | Class 5: ≥5 | |||
| F: lower center | ||||
| G: lower left | ||||
| Body and Neck |  | Class 1: 1–2 | ||
| Class 2: 3–4 | ||||
| A: right | Class 3: 5–6 | |||
| B: center | Class 4: 7–8 | |||
| C: left | Class 5: 9–10 | |||
| Class 6: >10 | ||||
| Limbs |  | A: right forelimb | Class 1: 1 | |
| B: left forelimb | ||||
| C: right hindlimb | Class 2: 2 | |||
| D: left hindlimb | Class 3: ≥ 3 | |||
| Location | Without | Top Right | Top Center | Top Left | Between Eyes | Low Right | Low Center | Low Left | Symmetry | |
|---|---|---|---|---|---|---|---|---|---|---|
| Head | Circular Whorls | 1828 (4.21%) | 993 (2.28%) | 10,785 (24.85%) | 1728 (3.98%) | 23,396 (53.91%) | 21,430 (49.38%) | 8131 (18.74%) | 22,308 (47.03%) | 20,972 (87.25%) |
| Linear Whorls | 39,124 (90.15%) | 40 (0.09%) | 618 (1.42%) | 78 (0.18%) | 1156 (2.66%) | 1501 (3.46%) | 1569 (3.62%) | 1546 (3.56%) | 1422 (87.56%) | |
| Body–neck | Without | Right | Center | Left | Symmetry | |||||
| Circular Whorls | 2019 (4.65%) | 38,712 (89.20%) | 37,335 (86.03%) | 38,757 (89.30%) | 38,252 (98.70%) | |||||
| Linear Whorls | 16,671 (38.41%) | 18,910 (43.57%) | 24,325 (56.05%) | 18,906 (43.56%) | 18,571 (98.21%) | |||||
| Limbs | Without | Right forelimb | Left forelimb | Right hindlimb | Left hindlimb | Symmetry | ||||
| Circular Whorls | 38,777 (83.35%) | 666 (1.53%) | 696 (1.60%) | 3723 (8.58%) | 3699 (8.52%) | 3885 (87.92%) | ||||
| Linear Whorls | 39,898 (91.94%) | 250 (0.58%) | 271 (0.62%) | 3128 (7.21%) | 3100 (7.14%) | 3056 (89.91%) | ||||
| Without | 1 | 2 | 3 | 4 | ≥5 | |||
|---|---|---|---|---|---|---|---|---|
| Head | Circular Whorls | 1828 (4.21%) | 14,710 (33.89%) | 5915 (13.62%) | 13,645 (31.44%) | 4218 (9.72%) | 3086 (7.11%) | |
| Linear Whorls | Without | 1 | 2 | ≥3 | ||||
| 39,124 (90.15%) | 2044 (4.71%) | 1398 (3.22%) | 319 (0.73%) | |||||
| Body–neck | Without | 1–2 | 3–4 | 5–6 | 7–8 | 9–10 | >10 | |
| Circular Whorls | 2019 (4.65%) | 3287 (7.57%) | 6907 (15.92%) | 8751 (20.16%) | 8120 (18.71%) | 6002 (13.83%) | 8312 (19.15%) | |
| Linear Whorls | 16,671 (38.41%) | 7390 (17.03%) | 5386 (12.41%) | 6736 (15.52%) | 4397 (10.13%) | 1780 (4.10%) | 1038 (2.39%) | |
| Limbs | Without | 1 | 2 | ≥3 | ||||
| Circular Whorls | 38,777 (83.35%) | 692 (1.59%) | 3705 (8.54%) | 224 (0.52%) | ||||
| Linear Whorls | 39,898 (91.94%) | 419 (0.97%) | 2942 (6.78%) | 139 (0.32%) | ||||
| Location | Position | σu | σe | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | Median | HPD 95% | Mean | Median | HPD 95% | h2 | |||
| Circular Whorls | Head | Top right | 3.958 | 3.919 | 3.609–4.378 | 0.976 | 0.982 | 0.926–1.019 | 0.802 |
| Top center | 5.187 | 5.161 | 4.064–6.282 | 0.933 | 0.950 | 0.827–1.010 | 0.848 | ||
| Top left | 0.377 | 0.378 | 0.301–0.457 | 1.000 | 1.000 | 0.977–1.025 | 0.274 | ||
| Between eyes | 0.246 | 0.244 | 0.200–0.295 | 1.017 | 0.997 | 0.666–1.383 | 0.195 | ||
| Low right | 0.295 | 0.244 | 0.085–1.118 | 1.002 | 0.999 | 0.913–1.116 | 0.227 | ||
| Low center | 3.362 | 3.775 | 0.642–5.161 | 0.998 | 0.998 | 0.904–1.099 | 0.771 | ||
| Low left | 0.276 | 0.206 | 0.128–0.905 | 1.001 | 1.001 | 0.904–1.079 | 0.216 | ||
| Body-neck | Right | 8.011 | 7.930 | 5.731–10.240 | 0.985 | 0.986 | 0.958–1.013 | 0.890 | |
| Center | 7.793 | 7.674 | 5.733–10.110 | 0.986 | 0.986 | 0.959–1.012 | 0.880 | ||
| Left | 4.860 | 4.853 | 3.515–6.399 | 0.983 | 0.983 | 0.959–1.007 | 0.832 | ||
| Limbs | Right front | 2.010 | 2.007 | 1.803–2.214 | 0.523 | 0.523 | 0.504–0.540 | 0.793 | |
| Left front | 1.922 | 1.919 | 1.731–2.131 | 0.520 | 0.520 | 0.501–0.537 | 0.787 | ||
| Right back | 6.670 | 6.637 | 5.776–7.858 | 0.760 | 0.760 | 0.735–0.791 | 0.898 | ||
| Left back | 6.692 | 6.640 | 5.667–7.812 | 0.760 | 0.760 | 0.731–0.786 | 0.898 | ||
| Linear Whorls | Head | Top right | 0.200 | 0.204 | 0.165–0.228 | 0.189 | 0.189 | 0.180–0.198 | 0.515 |
| Top center | 0.866 | 0.864 | 0.765–0.975 | 0.758 | 0.757 | 0.711–0.805 | 0.533 | ||
| Top left | 0.402 | 0.407 | 0.337–0.457 | 0.270 | 0.270 | 0.258–0.287 | 0.598 | ||
| Between eyes | 1.841 | 1.833 | 1.558–2.195 | 0.720 | 0.721 | 0.668–0.782 | 0.718 | ||
| Low right | 3.991 | 4.094 | 2.419–5.175 | 0.748 | 0.742 | 0.453–1.055 | 0.842 | ||
| Low center | 2.487 | 2.476 | 2.157–2.860 | 0.767 | 0.768 | 0.709–0.821 | 0.764 | ||
| Low left | 3.834 | 3.828 | 3.452–4.213 | 0.804 | 0.804 | 0.752–0.849 | 0.827 | ||
| Body–neck | Right | 0.136 | 0.136 | 0.131–0.140 | 0.079 | 0.077 | 0.075–0.081 | 0.636 | |
| Center | 0.136 | 0.136 | 0.131–0.140 | 0.078 | 0.079 | 0.075–0.807 | 0.636 | ||
| Left | 0.102 | 0.102 | 0.097–0.107 | 0.128 | 0.129 | 0.125–0.132 | 0.442 | ||
| Limbs | Right forelimb | 0.842 | 0.843 | 0.739–0.953 | 0.406 | 0.406 | 0.385–0.429 | 0.675 | |
| Left forelimb | 0.883 | 0.883 | 0.796–0.973 | 0.396 | 0.396 | 0.379–0.415 | 0.690 | ||
| Right hindlimb | 4.876 | 4.876 | 4.391–5.499 | 0.983 | 0.983 | 0.960–1.005 | 0.832 | ||
| Left hindlimb | 4.798 | 4.822 | 3.983–5.442 | 0.981 | 0.981 | 0.958–1.004 | 0.830 | ||
| σu | σe | h2 | ||||||
|---|---|---|---|---|---|---|---|---|
| Location | Particularity | Mean | Median | HPD 95% | Mean | Median | HPD 95% | |
| Head | Circular whorls | 0.646 | 0.646 | 0.604–0.683 | 1.013 | 1.012 | 0.982–1.044 | 0.389 |
| Linear whorls | 0.058 | 0.058 | 0.053–0.062 | 0.164 | 0.164 | 0.160–0.169 | 0.260 | |
| Body–neck | Circular whorls | 1.459 | 1.459 | 1.389–1.519 | 1.415 | 1.415 | 1.367–1.462 | 0.508 |
| Linear whorls | 1.500 | 1.500 | 1.445–1.555 | 1.092 | 1.092 | 1.054–1.131 | 0.579 | |
| Limbs | Circular whorls | 0.115 | 0.115 | 0.107–0.124 | 0.246 | 0.246 | 0.239–0.253 | 0.318 |
| Linear whorls | 0.126 | 0.126 | 0.120–0.133 | 0.158 | 0.158 | 0.153–0.163 | 0.444 | |
| Location and Position | Head | Body-Neck | Limbs | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Top Right | Top Center | Top Left | Between Eyes | Low Right | Low Centre | Low Left | Right | Centre | Left | Right | Left | Right | Left | ||
| Forelimbs | Hindlimbs | ||||||||||||||
| Head | Top right | −0.081 | −0.184 | 0.377 | 0.141 | 0.300 | 0.148 | 0.047 | 0.058 | 0.047 | 0.032 | 0.108 | 0.104 | 0.108 | |
| Top center | 0.005 | 0.437 | 0.123 | 0.280 | 0.720 | 0.286 | 0.250 | 0.238 | 0.251 | −0.037 | −0.054 | 0.156 | 0.152 | ||
| Top left | −0.771 | 0.044 | 0.202 | 0.103 | 0.226 | 0.113 | 0.169 | 0.165 | 0.221 | −0.272 | −0.206 | 0.085 | 0.055 | ||
| Between eyes | 0.040 | −0.440 | 0.081 | 0.266 | 0.306 | 0.290 | 0.380 | 0.379 | 0.382 | 0.095 | 0.129 | 0.152 | 0.147 | ||
| Low right | −0.426 | 0.164 | 0.578 | −0.229 | 0.348 | 0.998 | 0.147 | 0.147 | 0.151 | 0.290 | 0.335 | 0.444 | 0.445 | ||
| Low center | −0.048 | −0.040 | −0.095 | −0.446 | −0.051 | 0.351 | 0.233 | 0.229 | 0.149 | 0.141 | 0.110 | 0.184 | 0.177 | ||
| Low left | −0.078 | 0.101 | 0.270 | −0.195 | 0.807 | 0.053 | 0.146 | 0.146 | 0.146 | 0.279 | 0.323 | 0.443 | 0.443 | ||
| Body-neck | Right | 0.028 | 0.046 | 0.074 | −0.021 | 0.998 | −0.120 | 0.998 | 0.999 | 0.773 | 0.060 | 0.038 | 0.096 | 0.096 | |
| Center | 0.028 | −0.017 | 0.062 | 0.220 | 0.239 | −0.125 | 0.252 | 0.973 | 0.769 | 0.054 | 0.034 | 0.088 | 0.087 | ||
| Left | 0.018 | 0.053 | −0.005 | 0.109 | 0.998 | −0.121 | 0.998 | 0.792 | 0.900 | 0.084 | 0.034 | 0.104 | 0.101 | ||
| Limbs | Right forelimb | −0.172 | 0.007 | −0.081 | 0.01 | 0.027 | 0.066 | 0.033 | 0.056 | 0.048 | 0.058 | 0.932 | 0.375 | 0.372 | |
| Left forelimb | −0.252 | 0.001 | 0.164 | 0.031 | 0.026 | 0.054 | 0.024 | 0.068 | 0.065 | 0.062 | 0.926 | 0.417 | 0.419 | ||
| Right hindlimb | 0.028 | 0.035 | −0.024 | 0.030 | 0.038 | −0.065 | 0.060 | 0.176 | 0.998 | 0.190 | 0.082 | 0.071 | 0.998 | ||
| Left hindlimb | 0.016 | 0.032 | −0.027 | 0.034 | 0.038 | −0.055 | 0.048 | 0.998 | 0.998 | 0.189 | 0.096 | 0.061 | 0.828 | ||
| Location | Head | Body–Neck | Limbs | |||
|---|---|---|---|---|---|---|
| Linear Whorls | Circular Whorls | Linear Whorls | Circular Whorls | Linear Whorls | ||
| Head | Circular whorls | 0.213 | 0.012 | −0.004 | 0.138 | 0.054 |
| Linear whorls | 0.153 | 0.335 | 0.272 | 0.448 | ||
| Body–neck | Circular whorls | 0.429 | 0.349 | 0.130 | ||
| Linear whorls | 0.183 | 0.125 | ||||
| Limbs | Circular whorls | 0.618 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Encina, A.; Ligero, M.; Sánchez-Guerrero, M.J.; Rodríguez-Sainz de los Terreros, A.; Bartolomé, E.; Valera, M. Phenotypic and Genetic Study of the Presence of Hair Whorls in Pura Raza Español Horses. Animals 2023, 13, 2943. https://doi.org/10.3390/ani13182943
Encina A, Ligero M, Sánchez-Guerrero MJ, Rodríguez-Sainz de los Terreros A, Bartolomé E, Valera M. Phenotypic and Genetic Study of the Presence of Hair Whorls in Pura Raza Español Horses. Animals. 2023; 13(18):2943. https://doi.org/10.3390/ani13182943
Chicago/Turabian StyleEncina, Ana, Manuel Ligero, María José Sánchez-Guerrero, Arancha Rodríguez-Sainz de los Terreros, Ester Bartolomé, and Mercedes Valera. 2023. "Phenotypic and Genetic Study of the Presence of Hair Whorls in Pura Raza Español Horses" Animals 13, no. 18: 2943. https://doi.org/10.3390/ani13182943
APA StyleEncina, A., Ligero, M., Sánchez-Guerrero, M. J., Rodríguez-Sainz de los Terreros, A., Bartolomé, E., & Valera, M. (2023). Phenotypic and Genetic Study of the Presence of Hair Whorls in Pura Raza Español Horses. Animals, 13(18), 2943. https://doi.org/10.3390/ani13182943