
Long-Term Dietary Fish Meal Substitution with the Black Soldier Fly Larval Meal Modifies the Caecal Microbiota and Microbial Pathway in Laying Hens

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets, Animals, and Sampling
2.2. Quantification of Free Amino Acid in Liver
2.3. Quantification of Fatty Acid Concentration in Liver
2.4. Measurement of Plasma Biochemical Parameter
2.5. Disaccharidase Activity Test in Intestinal Mucosa
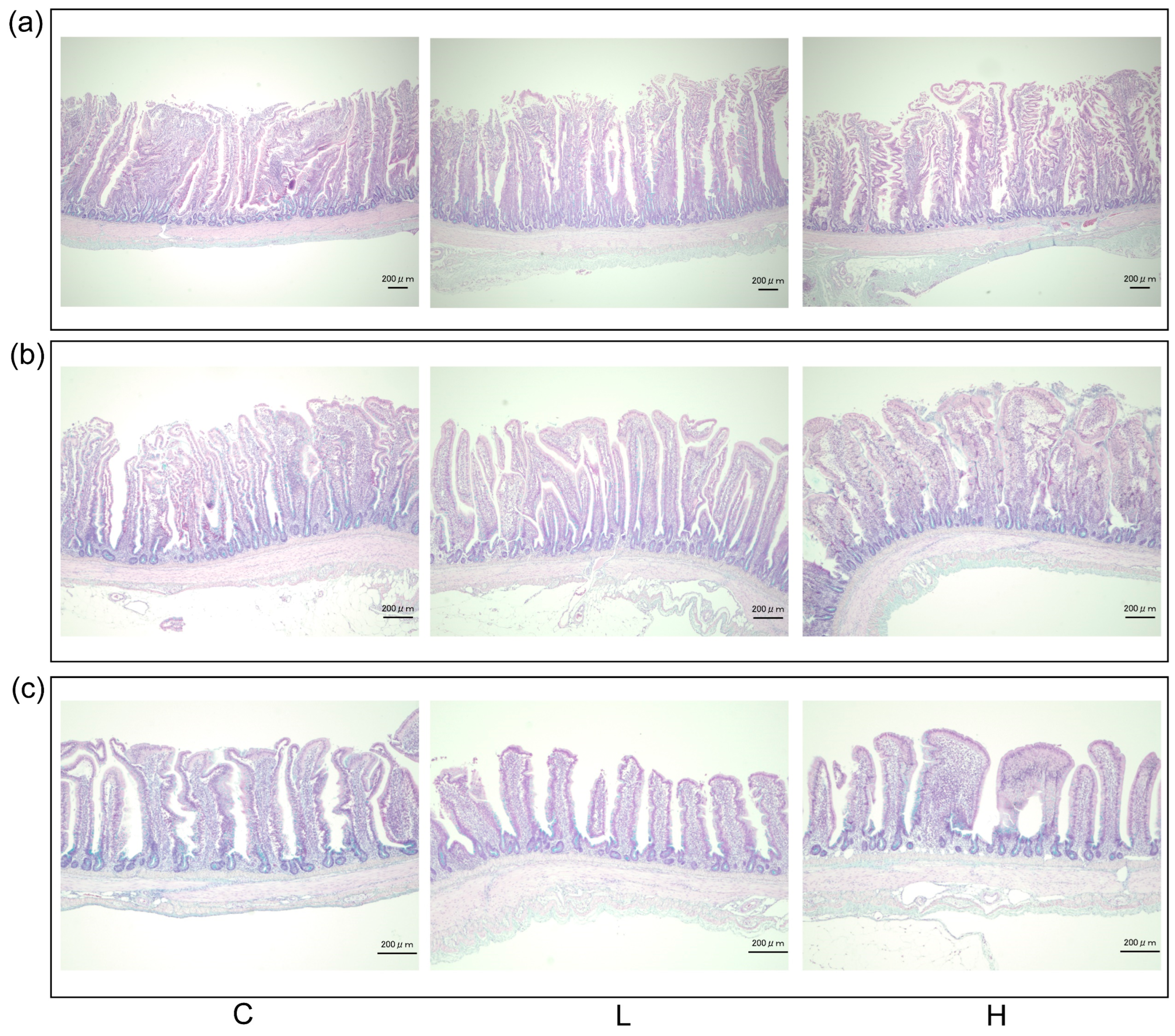
2.6. Intestinal Morphological Analysis
2.7. 16S rRNA Gene Amplicon Sequencing and Taxonomic Analysis
2.8. Determination of Short-Chain Fatty Acid in Caecal Content
2.9. Prediction of Microbial Functional Abundance and Metabolic Pathway Analysis
2.10. Statistical Analysis
3. Results
3.1. Free Amino Acids in the Liver
3.2. Fatty Acid Concentration in the Liver
3.3. Plasma Biochemical Parameters
3.4. Disaccharidase Activities in Intestinal Mucosa
3.5. Intestinal Morphology
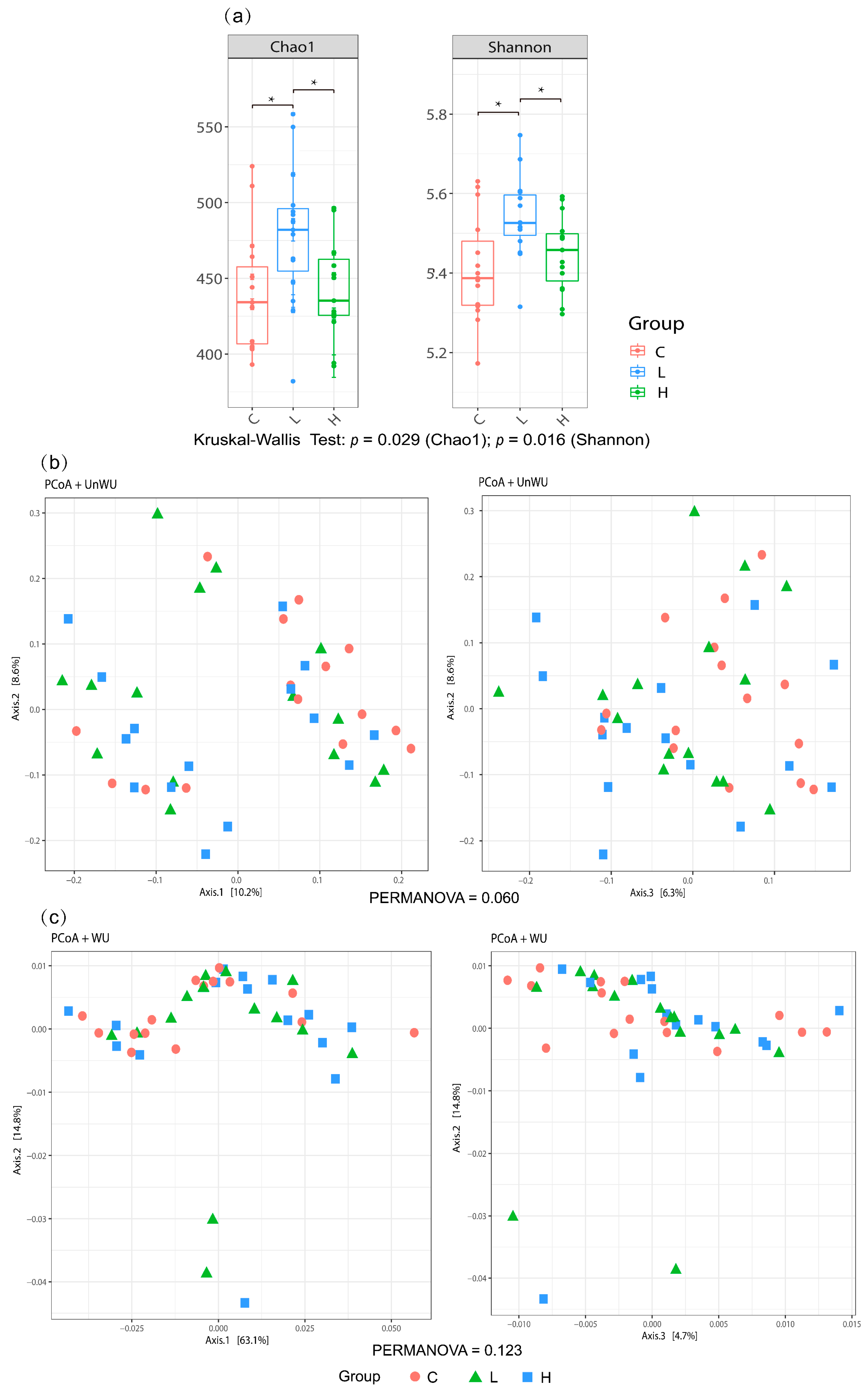
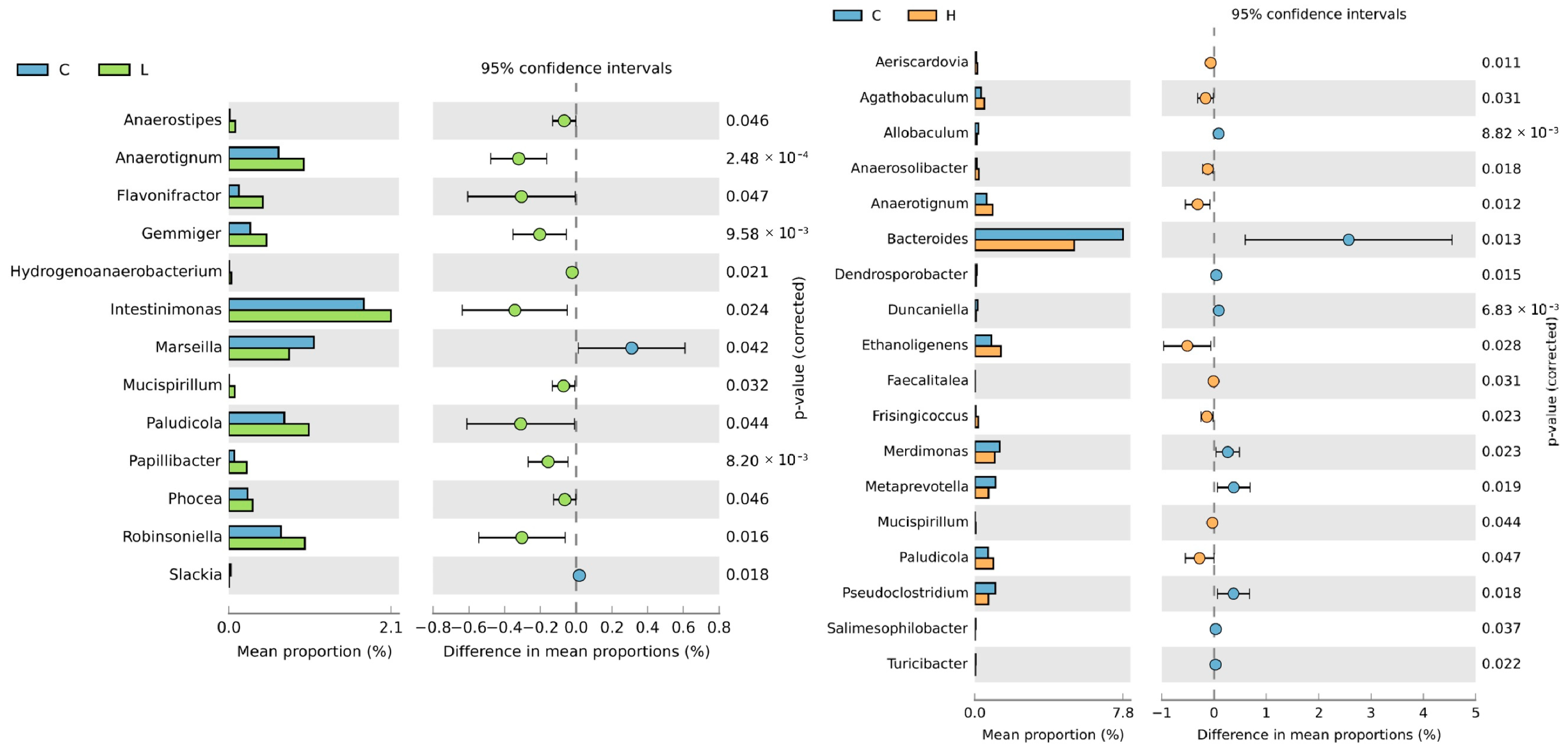
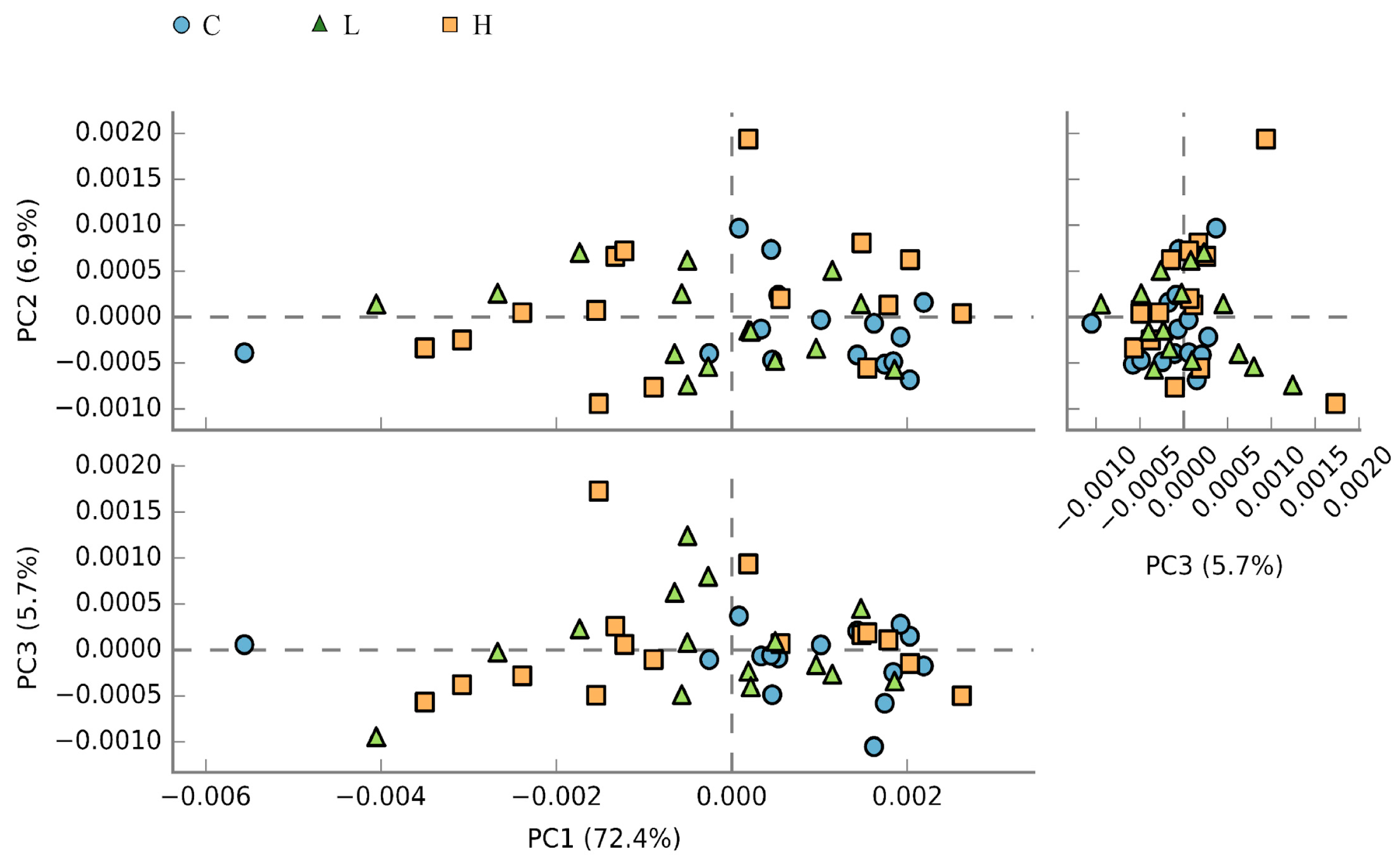
3.6. Caecal Microbiota
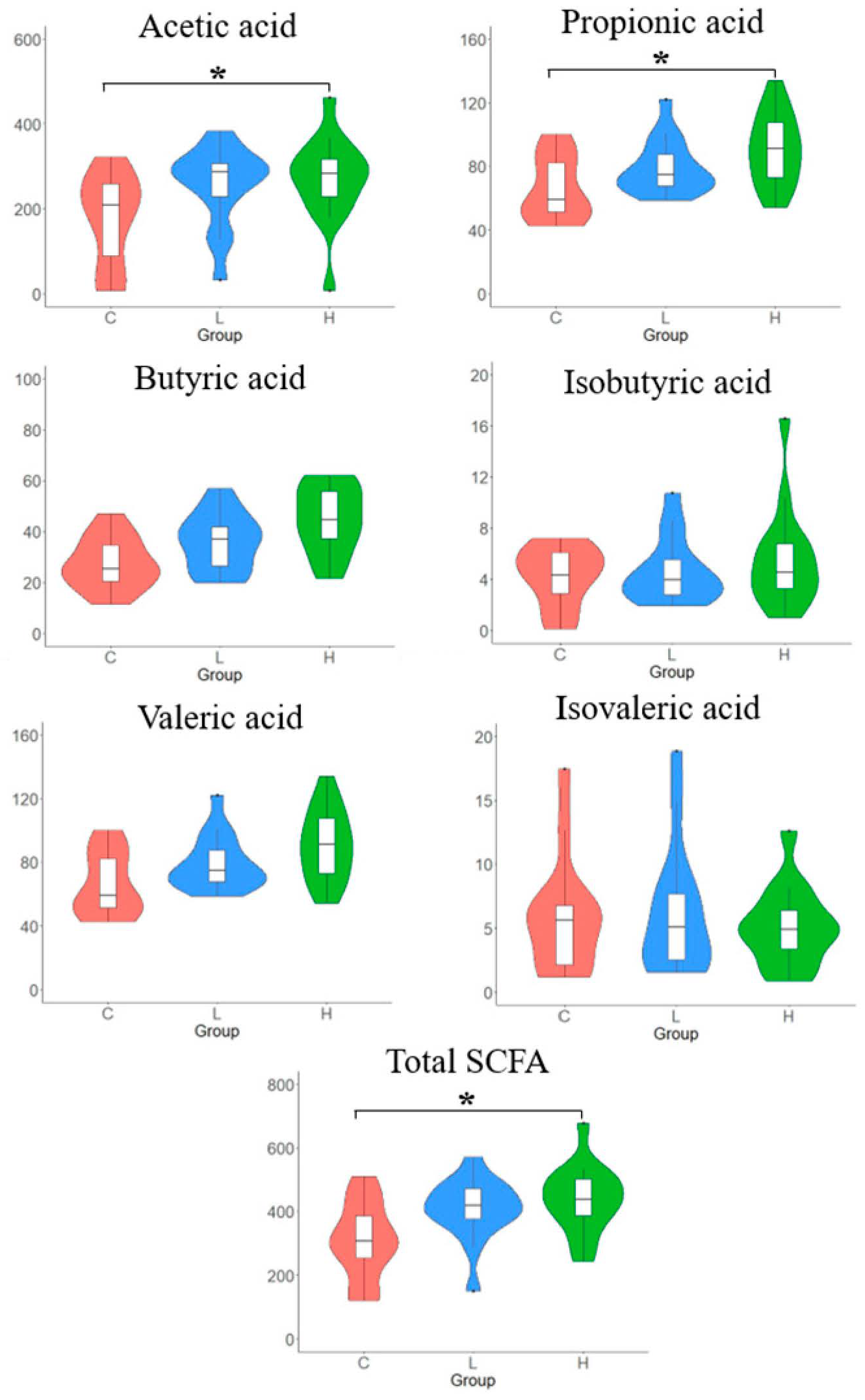
3.7. SCFAs Concentrations in Caecal Content
3.8. Microbial Functional Abundance Prediction and Metabolic Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Huis, A.; Gasco, L. Insects as feed for livestock production. Science 2023, 379, 138–139. [Google Scholar] [CrossRef]
- Yu, Z.; Zhao, L.; Zhao, J.-L.; Xu, W.; Guo, Z.; Zhang, A.-Z.; Li, M.-Y. Dietary Taraxacum mongolicum Polysaccharide Ameliorates the Growth, Immune Response, and Antioxidant Status in Association with NF-ΚB, Nrf2 and TOR in Jian Carp (Cyprinus carpio Var. Jian). Aquaculture 2022, 547, 737522. [Google Scholar] [CrossRef]
- Purschke, B.; Scheibelberger, R.; Axmann, S.; Adler, A.; Jäger, H. Impact of substrate contamination with mycotoxins, heavy metals and pesticides on the growth performance and composition of black soldier fly larvae (Hermetia illucens) for use in the feed and food value chain. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess 2017, 34, 1410–1420. [Google Scholar] [CrossRef]
- Pacheco, W.J.; Stark, C.R.; Ferket, P.R.; Brake, J. Effects of Trypsin Inhibitor and Particle Size of Expeller-Extracted Soybean Meal on Broiler Live Performance and Weight of Gizzard and Pancreas. Poult. Sci. 2014, 93, 2245–2252. [Google Scholar] [CrossRef]
- Hender, A.; Siddik, M.A.B.; Howieson, J.; Fotedar, R. Black Soldier Fly, Hermetia illucens as an Alternative to Fishmeal Protein and Fish Oil: Impact on Growth, Immune Response, Mucosal Barrier Status, and Flesh Quality of Juvenile Barramundi, Lates calcarifer (Bloch, 1790). Biology 2021, 10, 505. [Google Scholar] [CrossRef]
- Homska, N.; Kowalska, J.; Bogucka, J.; Ziółkowska, E.; Rawski, M.; Kierończyk, B.; Mazurkiewicz, J. Dietary Fish Meal Replacement with Hermetia illucens and Tenebrio molitor Larval Meals Improves the Growth Performance and Nutriphysiological Status of Ide (Leuciscus idus) Juveniles. Animals 2022, 12, 1227. [Google Scholar] [CrossRef]
- Biasato, I.; Renna, M.; Gai, F.; Dabbou, S.; Meneguz, M.; Perona, G.; Martinez, S.; Lajusticia, A.C.B.; Bergagna, S.; Sardi, L.; et al. Partially Defatted Black Soldier Fly Larva Meal Inclusion in Piglet Diets: Effects on the Growth Performance, Nutrient Digestibility, Blood Profile, Gut Morphology and Histological Features. J. Anim. Sci. Biotechnol. 2019, 10, 12. [Google Scholar] [CrossRef]
- Mutisya, M.M.; Agbodzavu, M.K.; Kinyuru, J.N.; Tanga, C.M.; Gicheha, M.; Hailu, G.; Salifu, D.; Khan, Z.; Niassy, S. Can Black Soldier Fly Desmodium Intortum Larvae-based Diets Enhance the Performance of Cobb500 Broiler Chickens and Smallholder Farmers’ Profit in Kenya? Poult. Sci. 2021, 100, 420–430. [Google Scholar] [CrossRef]
- Cheng, V.; Shoveller, A.K.; Huber, L.A.; Kiarie, E.G. Comparative Protein Quality in Black Soldier Fly Larvae Meal vs. Soybean Meal and Fish Meal using Classical Protein Efficiency Ratio (PER) Chick Growth Assay Model. Poult. Sci. 2023, 102, 102255. [Google Scholar] [CrossRef]
- Schiavone, A.; Dabbou, S.; De-Marco, M.; Cullere, M.; Biasato, I.; Biasibetti, E.; Capucchio, M.T.; Bergagna, S.; Dezzutto, D.; Meneguz, M.; et al. Black Soldier Fly Larva Fat Inclusion in Finisher Bbroiler Chicken Diet as an Alternative Fat Source. Animal 2018, 12, 2032–2039. [Google Scholar] [CrossRef]
- Heuel, M.; Sandrock, C.; Leiber, F.; Mathys, A.; Gold, M.; Zurbrüegg, C.; Gangnat, I.D.M.; Kreuzer, M.; Terranova, M. Black Soldier Fly Larvae Meal and Fat as a Replacement for Soybeans in Organic Broiler Diets: Effects on Performance, Body N Retention, Carcase and Meat Quality. Br. Poult. Sci. 2022, 63, 650–661. [Google Scholar] [CrossRef]
- Dabbou, S.; Lauwaerts, A.; Ferrocino, I.; Biasato, I.; Sirri, F.; Zampiga, M.; Bergagna, S.; Pagliasso, G.; Gariglio, M.; Colombino, E.; et al. Modified Black Soldier Fly Larva Fat in Broiler Diet: Effects on Performance, Carcass Traits, Blood Parameters, Histomorphological Features and Gut Microbiota. Animals 2021, 11, 1837. [Google Scholar] [CrossRef]
- Ndotono, E.W.; Khamis, F.M.; Bargul, J.L.; Tanga, C.M. Insights into the Gut Microbial Communities of Broiler Chicken Fed Black Soldier Fly Larvae-Desmodium-Based Meal as a Dietary Protein Source. Microorganisms 2022, 10, 1351. [Google Scholar] [CrossRef]
- Kawasaki, K.; Hashimoto, Y.; Hori, A.; Kawasaki, T.; Hirayasu, H.; Iwase, S.-I.; Hashizume, A.; Ido, A.; Miura, C.; Miura, T.; et al. Evaluation of Black Soldier Fly (Hermetia illucens) Larvae and Pre-Pupae Raised on Household Organic Waste, as Potential Ingredients for Poultry Feed. Animals 2019, 9, 98. [Google Scholar] [CrossRef]
- Ndotono, E.W.; Khamis, F.M.; Bargul, J.L.; Tanga, C.M. Gut Microbiota Shift in Layer Pullets Fed on Black Soldier Fly Larvae-Based Feeds towards Enhancing Healthy Gut Microbial Community. Sci. Rep. 2022, 12, 16714. [Google Scholar] [CrossRef]
- Biasato, I.; Ferrocino, I.; Dabbou, S.; Evangelista, R.; Gai, F.; Gasco, L.; Cocolin, L.; Capucchio, M.T.; Schiavone, A. Black Soldier Fly and Gut Health in Broiler Chickens: Insights into the Relationship between Cecal Microbiota and Intestinal Mucin Composition. J. Anim. Sci. Biotechnol. 2020, 11, 11. [Google Scholar] [CrossRef]
- Lee, K.-C.; Kil, D.Y.; Sul, W.J. Cecal Microbiome Divergence of Broiler Chickens by Sex and Body Weight. J. Microbiol. 2017, 55, 939–945. [Google Scholar] [CrossRef]
- Heuel, M.; Kreuzer, M.; Sandrock, C.; Leiber, F.; Mathys, A.; Gold, M.; Zurbrügg, C.; Gangnat, I.D.M.; Terranova, M. Transfer of Lauric and Myristic Acid from Black Soldier Fly Larval Lipids to Egg Yolk Lipids of Hens Is Low. Lipids 2021, 56, 423–435. [Google Scholar] [CrossRef]
- Zhao, J.; Kawasaki, K.; Miyawaki, H.; Hirayasu, H.; Izumo, A.; Iwase, S.-I.; Kasai, K. Egg Quality and Laying Performance of Julia Laying Hens Fed with Black Soldier Fly (Hermetia illucens) Larvae Meal as a Long-Term Substitute for Fish Meal. Poult. Sci. 2022, 101, 101986. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Kawasaki, K.; Wada, K.; Sato, A.; Zhao, J.; Takao, N.; Sato, M.; Ban, T.; Yano, K. Effects of Dietary Bamboo (Phyllostachys Pubescens Mazel) Culm Powder on Blood Properties and Intestinal Environment of Rabbits. Anim. Sci. J. 2022, 93, e13774. [Google Scholar] [CrossRef]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive Functional Profiling of Microbial Communities Using 16S RRNA Marker Gene Sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for Taxonomy-Based Analysis of Pathways and Genomes. Nucleic Acids Res. 2023, 51, D587–D592. [Google Scholar] [CrossRef]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical Analysis of Taxonomic and Functional Profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef]
- LaNoue, K.F.; Bryla, J.; Bassett, D.J. Energy-Driven Aspartate Efflux from Heart and Liver Mitochondria. J. Biol. Chem. 1974, 249, 7514–7521. [Google Scholar] [CrossRef]
- Belghit, I.; Waagbø, R.; Lock, E.-J.; Liland, N.S. Insect-Based Diets High in Lauric Acid Reduce Liver Lipids in Freshwater Atlantic Salmon. Aquac. Nutr. 2019, 25, 343–357. [Google Scholar] [CrossRef]
- Onsongo, V.O.; Osuga, I.M.; Gachuiri, C.K.; Wachira, A.M.; Miano, D.M.; Tanga, C.M.; Ekesi, S.; Nakimbugwe, D.; Fiaboe, K.K.M. Insects for Income Generation through Animal Feed: Effect of Dietary Replacement of Soybean and Fish Meal with Black Soldier Fly Meal on Broiler Growth and Economic Performance. J. Econ. Entomol. 2018, 111, 1966–1973. [Google Scholar] [CrossRef]
- Peng, K.; Mo, W.; Xiao, H.; Wang, G.; Huang, Y. Effects of Black Soldier Fly Pulp on Growth Performance, Histomorphology and Lipid Metabolism Gene Expression of Micropterus salmoides. Aquac. Rep. 2021, 20, 100737. [Google Scholar] [CrossRef]
- Gariglio, M.; Dabbou, S.; Crispo, M.; Biasato, I.; Gai, F.; Gasco, L.; Piacente, F.; Odetti, P.; Bergagna, S.; Plachà, I.; et al. Effects of the Dietary Inclusion of Partially Defatted Black Soldier Fly (Hermetia illucens) Meal on the Blood Chemistry and Tissue (Spleen, Liver, Thymus, and Bursa of Fabricius) Histology of Muscovy Ducks (Cairina Moschata domestica). Animals 2019, 9, 307. [Google Scholar] [CrossRef]
- Loponte, R.; Nizza, S.; Bovera, F.; De Riu, N.; Fliegerova, K.; Lombardi, P.; Vassalotti, G.; Mastellone, V.; Nizza, A.; Moniello, G. Growth Performance, Blood Profiles and Carcass Traits of Barbary Partridge (Alectoris barbara) Fed Two Different Insect Larvae Meals (Tenebrio molitor and Hermetia illucens). Res. Vet. Sci. 2017, 115, 183–188. [Google Scholar] [CrossRef]
- Marono, S.; Loponte, R.; Lombardi, P.; Vassalotti, G.; Pero, M.E.; Russo, F.; Gasco, L.; Parisi, G.; Piccolo, G.; Nizza, S.; et al. Productive Performance and Blood Profiles of Laying Hens Fed Hermetia Illucens Larvae Meal as Total Replacement of Soybean Meal from 24 to 45 Weeks of Age. Poult. Sci. 2017, 96, 1783–1790. [Google Scholar] [CrossRef]
- Kim, B.; Bang, H.T.; Jeong, J.Y.; Kim, M.; Kim, K.H.; Chun, J.L.; Ji, S.Y. Effects of Dietary Supplementation of Black Soldier Fly (Hermetia illucens) Larvae Oil on Broiler Health. J. Poult. Sci. 2021, 58, 222–229. [Google Scholar] [CrossRef]
- Moniello, G.; Ariano, A.; Panettieri, V.; Tulli, F.; Olivotto, I.; Messina, M.; Randazzo, B.; Severino, L.; Piccolo, G.; Musco, N.; et al. Intestinal Morphometry, Enzymatic and Microbial Activity in Laying Hens Fed Different Levels of a Hermetia illucens Larvae Meal and Toxic Elements Content of the Insect Meal and Diets. Animals 2019, 9, 86. [Google Scholar] [CrossRef]
- Cutrignelli, M.I.; Messina, M.; Tulli, F.; Randazzo, B.; Olivotto, I.; Gasco, L.; Loponte, R.; Bovera, F. Evaluation of an Insect Meal of the Black Soldier Fly (Hermetia illucens) as Soybean Substitute: Intestinal Morphometry, Enzymatic and Microbial Activity in Laying Hens. Res. Vet. Sci. 2018, 117, 209–215. [Google Scholar] [CrossRef]
- Bovera, F.; Loponte, R.; Marono, S.; Piccolo, G.; Parisi, G.; Iaconisi, V.; Gasco, L.; Nizza, A. Use of Larvae Meal as Protein Source in Broiler Diet: Effect on Growth Performance, Nutrient Digestibility, and Carcass and Meat Traits. J. Anim. Sci. 2016, 94, 639–647. [Google Scholar] [CrossRef]
- Ueki, A.; Goto, K.; Ohtaki, Y.; Kaku, N.; Ueki, K. Description of Anaerotignum aminivorans Gen. Nov., Sp. Nov., a Strictly Anaerobic, Amino-Acid-Decomposing Bacterium Isolated from a Methanogenic Reactor, and Reclassification of Clostridium Propionicum, Clostridium neopropionicum and Clostridium lactatifermentans as Species of the Genus Anaerotignum. Int. J. Syst. Evol. Microbiol. 2017, 67, 4146–4153. [Google Scholar] [CrossRef]
- Kläring, K.; Hanske, L.; Bui, N.; Charrier, C.; Blaut, M.; Haller, D.; Plugge, C.M.; Clavel, T. Intestinimonas butyriciproducens Gen. Nov., Sp. Nov., a Butyrate-Producing Bacterium from the Mouse Intestine. Int. J. Syst. Evol. Microbiol. 2013, 63, 4606–4612. [Google Scholar] [CrossRef]
- Deb, D.; Das, S.; Adak, A.; Khan, M.R. Traditional Rice Beer Depletes Butyric Acid-Producing Gut Bacteria Faecalibacterium and Roseburia along with Fecal Butyrate Levels in the Ethnic Groups of Northeast India. 3 Biotech 2020, 10, 283. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, L.-L.; Liu, L.; Tian, Y.-Q.; Liu, X.-F.; Li, W.-J.; Dai, Y.-M. Paludicola psychrotolerans Gen. Nov., Sp. Nov., a Novel Psychrotolerant Chitinolytic Anaerobe of the Family Ruminococcaceae. Int. J. Syst. Evol. Microbiol. 2017, 67, 4100–4103. [Google Scholar] [CrossRef]
- Chen, Y.; Akhtar, M.; Ma, Z.; Hu, T.; Liu, Q.; Pan, H.; Zhang, X.; Nafady, A.A.; Ansari, A.R.; Abdel-Kafy, E.M.; et al. Chicken Cecal Microbiota Reduces Abdominal Fat Deposition by Regulating Fat Metabolism. NPJ Biofilms Microbiomes 2023, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Seaton, S.C.; Ndousse-Fetter, S.; Adhikari, A.A.; DiBenedetto, N.; Mina, A.I.; Banks, A.S.; Bry, L.; Devlin, A.S. A Selective Gut Bacterial Bile Salt Hydrolase Alters Host Metabolism. eLife 2018, 7, e37182. [Google Scholar] [CrossRef] [PubMed]
- Li, D.K.; Chaudhari, S.N.; Lee, Y.; Sojoodi, M.; Adhikari, A.A.; Zukerberg, L.; Shroff, S.; Barrett, S.C.; Tanabe, K.; Chung, R.T.; et al. Inhibition of Microbial Deconjugation of Micellar Bile Acids Protects Against Intestinal Permeability and Liver Injury. Sci. Adv. 2022, 8, eabo2794. [Google Scholar] [CrossRef] [PubMed]
- Weththasinghe, P.; Lagos, L.; Cortés, M.; Hansen, J.Ø.; Øverland, M. Dietary Inclusion of Black Soldier Fly (Hermetia illucens) Larvae Meal and Paste Improved Gut Health but Had Minor Effects on Skin Mucus Proteome and Immune Response in Atlantic Salmon (Salmo salar). Front. Immunol. 2021, 12, 599530. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acids (mmol/L) | Diets | p-Value | ||
|---|---|---|---|---|
| C Diet | L Diet | H Diet | ||
| Alanine | 1.694 ± 0.076 | 1.648 ± 0.084 | 1.865 ± 0.133 | 0.527 |
| Arginine | 2.223 ± 0.130 | 2.045 ± 0.085 | 2.724 ± 0.238 | 0.123 |
| Asparagine | 0.363 ± 0.029 | 0.342 ± 0.031 | 0.491 ± 0.053 | 0.059 |
| Aspartate | 1.749 ± 0.105 bc | 1.610 ± 0.080 c | 2.275 ± 0.176 a | 0.023 |
| Glutamate | 2.486 ± 0.143 | 2.293 ± 0.128 | 2.868 ± 0.237 | 0.145 |
| Glutamine | 2.752 ± 0.183 | 2.543 ± 0.149 | 3.317 ± 0.316 | 0.181 |
| Glycine | 0.757 ± 0.039 | 0.760 ± 0.041 | 0.922 ± 0.072 | 0.171 |
| Histidine | 5.480 ± 0.274 | 5.178 ± 0.270 | 5.809 ± 0.491 | 0.783 |
| Isoleucine | 1.139 ± 0.068 | 1.066 ± 0.046 | 1.435 ± 0.200 | 0.070 |
| Leucine | 1.250 ± 0.073 | 1.179 ± 0.049 | 1.582 ± 0.132 | 0.072 |
| Lysine | 3.126 ± 0.184 | 3.103 ± 0.152 | 4.090 ± 0.365 | 0.145 |
| Methionine | 2.684 ± 0.146 | 2.495 ± 0.109 | 3.378 ± 0.278 | 0.052 |
| Phenylalanine | 1.348 ± 0.077 | 1.229 ± 0.058 | 1.436 ± 0.112 | 0.450 |
| Proline | 2.399 ± 0.130 | 2.287 ± 0.105 | 2.865 ± 0.245 | 0.333 |
| Serine | 5.326 ± 0.232 | 4.754 ± 0.221 | 5.539 ± 0.377 | 0.611 |
| Threonine | 1.473 ± 0.082 | 1.341 ± 0.062 | 1.785 ± 0.146 | 0.065 |
| Tryptophan | 0.192 ± 0.010 | 0.180 ± 0.009 | 0.191 ± 0.012 | 0.950 |
| Tyrosine | 1.482 ± 0.069 | 1.350 ± 0.063 | 1.487 ± 0.117 | 0.418 |
| Valine | 2.504 ± 0.137 | 2.250 ± 0.102 | 2.598 ± 0.219 | 0.458 |
| Fatty Acids (mg/g) | Diets | p-Value | ||
|---|---|---|---|---|
| C Diet | L Diet | H Diet | ||
| Linoleic acid (C22: 2(n-6)) | 0.80 ± 0.16 | 0.76 ± 0.09 | 0.84 ± 0.15 | 0.826 |
| Oleic acid (C22: 0) | 5.01 ± 0.73 | 6.27 ± 1.07 | 5.85 ± 1.02 | 0.665 |
| Palmitic acid (C16: 0) | 1.32 ± 0.21 | 1.84 ± 0.45 | 1.54 ± 0.26 | 0.693 |
| Stearic acid (C18: 0) | 0.77 ± 0.17 | 0.92 ± 0.18 | 0.83 ± 0.13 | 0.377 |
| Parameters | Diets | p-Value | ||
|---|---|---|---|---|
| C Diet | L Diet | H Diet | ||
| ALP (IU/L) | 322.86 ± 42.82 | 365.23 ± 59.12 | 281.73 ± 25.27 | 0.806 |
| GOT (IU/L) | 112.14 ± 10.86 | 144.62 ± 11.71 | 123.20 ± 9.88 | 0.069 |
| GGT (IU/L) | 17.86 ± 1.33 | 19.38 ± 1.75 | 18.80 ± 1.26 | 0.762 |
| LDH (IU/L) | 214.86 ± 20.76 | 336.46 ± 46.00 | 307.60 ± 44.73 | 0.138 |
| HDL-c (mg/dL) | 22.79 ± 7.05 | 20.67 ± 9.04 | 19.57 ± 8.76 | 0.305 |
| T-cho (mg/dL) | 104.00 ± 27.30 | 100.33 ± 26.92 | 103.33 ± 27.40 | 0.901 |
| Ca (mg/dL) | 22.73 ± 1.40 | 25.75 ± 1.38 | 23.85 ± 1.55 | 0.160 |
| Mg (mg/dL) | 2.80 ± 0.18 | 3.28 ± 0.22 | 2.97 ± 0.20 | 0.264 |
| IP (mg/dL) | 7.30 ± 0.80 | 8.11 ± 0.78 | 7.49 ± 0.79 | 0.513 |
| IgG (mg/mL) | 7.43 ± 1.37 | 7.56 ± 1.62 | 9.69 ± 1.27 | 0.479 |
| TNF-α (pg/mL) | 148.78 ± 37.49 | 134.87 ± 54.60 | 134.94 ± 43.69 | 0.834 |
| Disaccharidase Activities (mU/mg) | Diets | p-Value | ||
|---|---|---|---|---|
| C Diet | L Diet | H Diet | ||
| Jejunum | ||||
| Maltase activity | 0.949 ± 0.060 | 0.845 ± 0.058 | 0.868 ± 0.079 | 0.971 |
| Sucrase activity | 0.678 ± 0.028 | 0.688 ± 0.033 | 0.687 ± 0.032 | 0.344 |
| Ileum | ||||
| Maltase activity | 0.397 ± 0.030 ab | 0.318 ± 0.026 b | 0.429 ± 0.032 a | 0.025 |
| Sucrase activity | 0.165 ± 0.042 | 0.111 ± 0.025 | 0.240 ± 0.078 | 0.218 |
| Intestines | Diets | p-Value | ||
|---|---|---|---|---|
| C Diet | L Diet | H Diet | ||
| Duodenum | ||||
| Villi height (µm) | 1082.95 ± 51.08 | 1031.17 ± 51.81 | 1040.39 ± 26.87 | 0.803 |
| Crypt depth (µm) | 168.77 ± 5.96 | 174.62 ± 5.43 | 176.09 ± 7.14 | 0.681 |
| Villi/Crypt | 6.57 ± 0.44 | 5.95 ± 0.30 | 6.09 ± 0.35 | 0.462 |
| Goblet cell density (103/mm2) | 2.21 ± 0.16 | 1.86 ± 0.07 | 1.78 ± 0.08 | 0.088 |
| Jejunum | ||||
| Villi height (µm) | 793.47 ± 53.03 | 771.34 ± 37.3 | 817.66 ± 44.70 | 0.773 |
| Crypt depth (µm) | 104.52 ± 4.80 | 113.1 ± 4.46 | 108.38 ± 4.59 | 0.320 |
| Villi/Crypt | 7.63 ± 0.44 | 6.89 ± 0.32 | 7.62 ± 0.43 | 0.337 |
| Goblet cell density (103/mm2) | 2.02 ± 0.14 | 1.62 ± 0.12 | 1.61 ± 0.17 | 0.085 |
| Ileum | ||||
| Villi height (µm) | 529.44 ± 24.64 | 561.27 ± 31.87 | 545.65 ± 14.03 | 0.787 |
| Crypt depth (µm) | 87.45 ± 4.50 | 90.81 ± 3.96 | 93.49 ± 6.50 | 0.705 |
| Villi/Crypt | 6.14 ± 0.26 | 6.20 ± 0.25 | 6.08 ± 0.31 | 0.957 |
| Goblet cell density (103/mm2) | 1.78 ± 0.14 | 1.65 ± 0.13 | 1.44 ± 0.06 | 0.139 |
| Pathways | Pathway Enrichment Ratio (%) | p-Value | ||
|---|---|---|---|---|
| C Diet | L Diet | H Diet | ||
| β-glucosidase | 2.37 ± 0.10 a | 2.23 ± 0.06 ab | 2.07 ± 0.09 bc | 0.041 |
| Long-chain acyl-CoA synthetase | 2.41 ± 0.10 a | 2.20 ± 0.06 ab | 2.06 ± 0.10 bc | 0.031 |
| UDP glucose 6-dehydrogenase | 2.36 ± 0.09 a | 2.25 ± 0.06 ab | 2.06 ± 0.09 bc | 0.041 |
| periplasmic protein TonB | 2.43 ± 0.13 a | 2.18 ± 0.08 ab | 2.05 ± 0.13 bc | 0.043 |
| Bile acid: Na+ symporter, BASS family | 2.41 ± 0.12 a | 2.20 ± 0.07 ab | 2.05 ± 0.11 bc | 0.041 |
| K+-stimulated pyrophosphate-energized sodium pump | 2.33 ± 0.08 a | 2.26 ± 0.05 ab | 2.08 ± 0.07 bc | 0.045 |
| Carboxynorspermidine decarboxylase | 2.34 ± 0.08 a | 2.25 ± 0.06 ab | 2.08 ± 0.07 bc | 0.034 |
| Saccharopine dehydrogenase | 2.33 ± 0.08 a | 2.25 ± 0.05 ab | 2.09 ± 0.07 bc | 0.050 |
| Alpha-D-xyloside xylohydrolase | 2.41 ± 0.09 a | 2.17 ± 0.07 ab | 2.09 ± 0.08 bc | 0.020 |
| 6-pyruvoyltetrahydropterin synthase | 2.38 ± 0.10 a | 2.23 ± 0.21 ab | 2.06 ± 0.10 bc | 0.050 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Ban, T.; Miyawaki, H.; Hirayasu, H.; Izumo, A.; Iwase, S.-i.; Kasai, K.; Kawasaki, K. Long-Term Dietary Fish Meal Substitution with the Black Soldier Fly Larval Meal Modifies the Caecal Microbiota and Microbial Pathway in Laying Hens. Animals 2023, 13, 2629. https://doi.org/10.3390/ani13162629
Zhao J, Ban T, Miyawaki H, Hirayasu H, Izumo A, Iwase S-i, Kasai K, Kawasaki K. Long-Term Dietary Fish Meal Substitution with the Black Soldier Fly Larval Meal Modifies the Caecal Microbiota and Microbial Pathway in Laying Hens. Animals. 2023; 13(16):2629. https://doi.org/10.3390/ani13162629
Chicago/Turabian StyleZhao, Junliang, Takuma Ban, Hironori Miyawaki, Hirofumi Hirayasu, Akihisa Izumo, Shun-ichiro Iwase, Koji Kasai, and Kiyonori Kawasaki. 2023. "Long-Term Dietary Fish Meal Substitution with the Black Soldier Fly Larval Meal Modifies the Caecal Microbiota and Microbial Pathway in Laying Hens" Animals 13, no. 16: 2629. https://doi.org/10.3390/ani13162629
APA StyleZhao, J., Ban, T., Miyawaki, H., Hirayasu, H., Izumo, A., Iwase, S.-i., Kasai, K., & Kawasaki, K. (2023). Long-Term Dietary Fish Meal Substitution with the Black Soldier Fly Larval Meal Modifies the Caecal Microbiota and Microbial Pathway in Laying Hens. Animals, 13(16), 2629. https://doi.org/10.3390/ani13162629