
Slaughtering of Water Buffalo (Bubalus bubalis) with and without Stunning: A Focus on the Neurobiology of Pain, Hyperalgesia, and Sensitization

 ,
,  ,
,  , ,
, ,  ,
, 
 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Anatomical Features of the Water Buffalo Skull
3. Importance of Stunning and Nociception before Slaughter
3.1. Slaughter with and without Stunning and Its Nociceptive Effects
3.2. Peripheral and Central Sensitization during and before Water Buffalo Slaughter
4. Stunning Methods for Water Buffaloes
Penetrating Captive Bolt Pistol
5. Methods to Evaluate the Quality of Unconsciousness, Return-to-Sensibility Signs, and Death
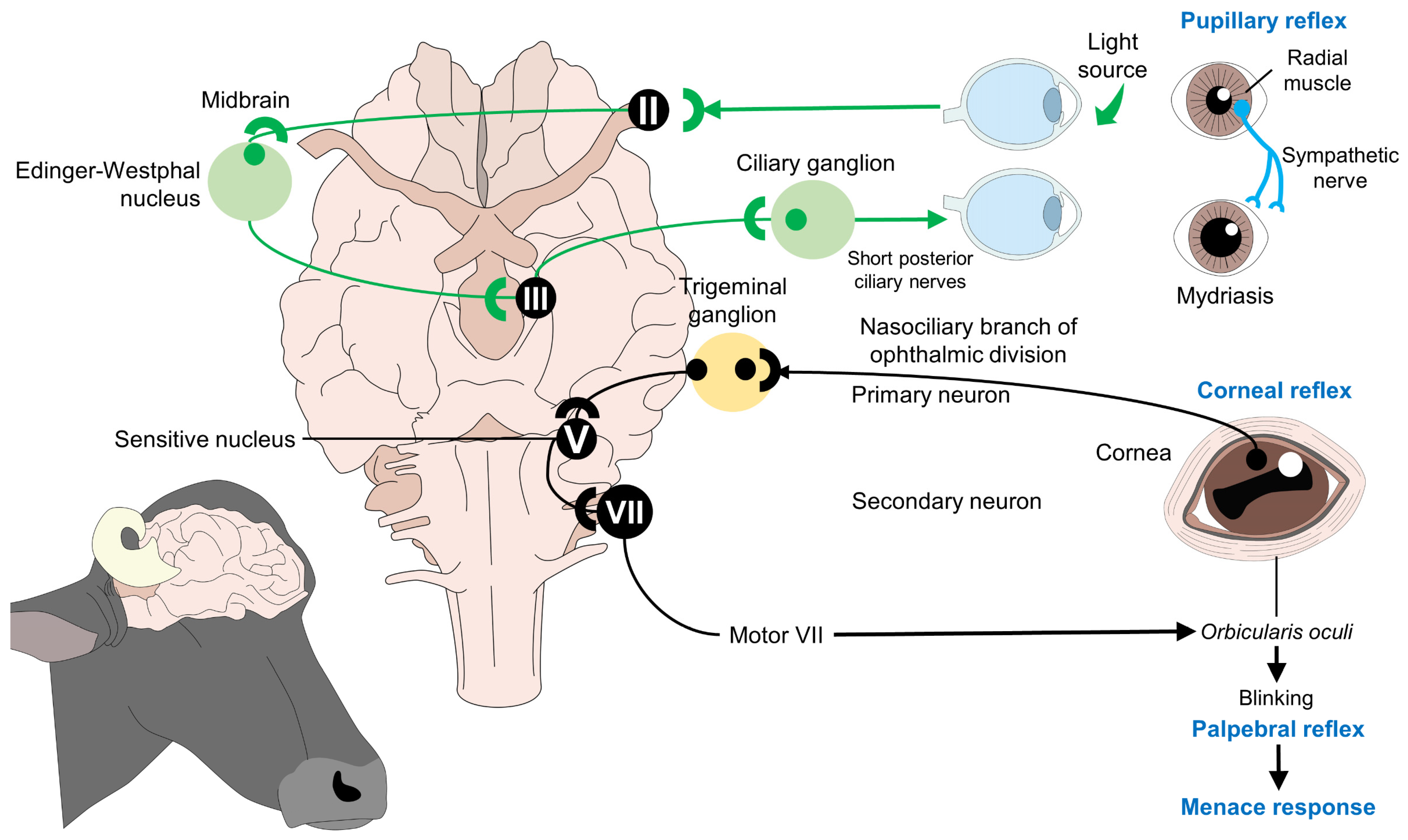
Ocular Signs of Consciousness during Slaughter
6. Future Research Aimed to Improve Stunning Methods in Water Buffaloes
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Alsafy, M.A.M.; El-Gendy, S.A.A.; El Sharaby, A.A. Anatomic reference for computed tomography of paranasal sinuses and their communication in the egyptian buffalo (Bubalus bubalis). Anat. Histol. Embryol. 2013, 42, 220–231. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Ghezzi, M.D.; Napolitano, F.; Rosmini, M.R.; Guerrero-Legarreta, I.; Martínez-Burnes, J.; Lezama-García, K.; Miranda-Cortés, A.; de la Vega, L.T.; Mora-Medina, P.; et al. Quality of death in the river buffalo (Bubalus bubalis). J. Anim. Behav. Biometeorol. 2021, 9, 2115. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Napolitano, F.; Strappini, A.; Orihuela, A.; Ghezzi, M.D.; Hernández-Ávalos, I.; Mora-Medina, P.; Whittaker, A.L. Pain at the slaughterhouse in ruminants with a focus on the neurobiology of sensitisation. Animals 2021, 11, 1085. [Google Scholar] [CrossRef]
- Gregory, N.G.; Spence, J.Y.; Mason, C.W.; Tinarwo, A.; Heasman, L. Effectiveness of poll stunning water buffalo with captive bolt guns. Meat Sci. 2009, 81, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Alarcón-Rojo, A.; Mota-Rojas, D.; García-Galicia, I.; Ramírez-Bribiesca, E.; Olmos-Hernández, A.; Guerrero-Legarreta, I. Dark cutting in large ruminants: Effect of management and environmental factors. Agro Prod. 2021, 13, 93–98. [Google Scholar] [CrossRef]
- Napolitano, F.; Strappini, A.; Ramírez-Bribiesca, E.; Alarcón-Rojo, A.D.; Cruz-Monterrosa, R.G.; Guerrero Legarreta, I.; Rayas-Amor, A.A.; García-Galicia, I.A.; Ghezzi, M.D.; Becerril-Herrera, M.; et al. Estresores previos a la muerte y su efecto en la calidad de la carne del búfalo de agua. In El Búfalo de Agua en Latinoamérica, Hallazgos Recientes, 3rd ed.; Napolitano, F., Mota-Rojas, D., Guerrero-Legarreta, I., Orihuela, A., Eds.; BM Editores: Mexico City, México, 2020; pp. 1178–1233. [Google Scholar]
- Cruz-Monterrosa, R.G.; Mota-Rojas, D.; Ramírez-Bibriesca, E.; Mora-Medina, P.; Guerrero-Legarreta, I. Scientific findings on the quality of river buffalo meat and prospects for future studies. J. Buffalo Sci. 2020, 9, 170–180. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Ghezzi, M.D.; Napolitano, F.; Hernández-Ávalos, I.; Rosmini, M.R.; de la Vega, L.T.; Cruz-Monterrosa, R.; Cajiao, M.N.; Miranda Cortés, A.; Torres-Bernal, F.; et al. Calidad del aturdimiento en búfalos: Reflejos y signos de retorno a la sensibilidad durante la muerte. In El Búfalo de Agua en Latinoamérica, Hallazgos Recientes, 3rd ed.; Napolitano, F., Mota-Rojas, D., Guerrero-Legarreta, I., Orihuela, A., Eds.; BM Editores: Mexico City, México, 2020; pp. 1073–1177. [Google Scholar]
- José-Pérez, N.; Mota-Rojas, D.; Ghezzi, M.D.; Rosmini, M.R.; Mora-Medina, P.; Bertoni, A.; Rodríguez-González, D.; Domínguez-Oliva, A.; Guerrero-Legarreta, I. Effects of transport on water buffaloes (Bubalus bubalis): Factors associated with the frequency of skin injuries and meat quality. J. Anim. Behav. Biometeorol. 2022, 10, 2216. [Google Scholar] [CrossRef]
- Rodríguez-González, D.; Minervino, A.H.H.; Orihuela, A.; Bertoni, A.; Morales-Canela, D.A.D.A.; Álvarez-Macías, A.; José-Pérez, N.; Domínguez-Oliva, A.; Mota-Rojas, D.; Hamad, A.M.; et al. Handling and physiological aspects of the dual-purpose water buffalo production system in the mexican humid tropics. Animals 2022, 12, 608. [Google Scholar] [CrossRef]
- Grandin, T. Making slaughterhouses more humane for cattle, pigs, and sheep. Annu. Rev. Anim. Biosci. 2013, 1, 491–512. [Google Scholar] [CrossRef]
- Velarde, A.; Dalmau, A. Slaughter without Stunning; Elsevier Ltd.: Barcelona, Spain, 2017; Volume I, pp. 222–240. [Google Scholar]
- Gregory, N.; von Wenzlawowicz, M.; von Holleben, K.; Fielding, H.; Gibson, T.; Mirabito, L.; Kolesar, R. Complications during shechita and halal slaughter without stunning in cattle. Anim. Welf. 2012, 21, 81–86. [Google Scholar] [CrossRef]
- Ellison, D.L. Physiology of pain. Crit. Care Nurs. Clin. North Am. 2017, 29, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Gibson, T.J.; Dadios, N.; Gregory, N.G. Effect of neck cut position on time to collapse in halal slaughtered cattle without stunning. Meat Sci. 2015, 110, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Gregory, N.G.; Fielding, H.R.; von Wenzlawowicz, M.; von Holleben, K. Time to collapse following slaughter without stunning in cattle. Meat Sci. 2010, 85, 66–69. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Napolitano, F.; Braghieri, A.; Guerrero-Legarreta, I.; Bertoni, A.; Martínez-Burnes, J.; Cruz-Monterrosa, R.; Gómez, J.; Ramírez-Bribiesca, E.; Barrios-García, H.; et al. Thermal biology in river buffalo in the humid tropics: Neurophysiological and behavioral responses assessed by infrared thermography. J. Anim. Behav. Biometeorol. 2021, 9, 2103. [Google Scholar] [CrossRef]
- Bertoni, A.; Napolitano, F.; Mota-Rojas, D.; Sabia, E.; Álvarez-Macías, A.; Mora-Medina, P.; Morales-Canela, A.; Berdugo-Gutiérrez, J.; Guerrero- Legarreta, I. Similarities and differences between river buffaloes and cattle: Health, physiological, behavioral and productivity aspects. J. Buffalo Sci. 2020, 9, 92–109. [Google Scholar] [CrossRef]
- Schwenk, B.K.; Lechner, I.; Ross, S.G.; Gascho, D.; Kneubuehl, B.P.; Glardon, M.; Stoffel, M.H. Magnetic resonance imaging and computer tomography of brain lesions in water buffaloes and cattle stunned with handguns or captive bolts. Meat Sci. 2016, 113, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Özkan, E.; Siddiq, A.B.; Kahvecioğlu, K.O.; Öztürk, M.; Onar, V. Morphometric analysis of the skulls of domestic cattle (Bostaurus L.) and water buffalo (Bubalus bubalis L.) in Turkey. Turkish J. Vet. Anim. Sci. 2019, 43, 532–539. [Google Scholar] [CrossRef]
- Molnar-Fernández, V.; Muñoz-Vargas, L.; Romero-Zúñiga, J.J.; Araya-Rodríguez, G. Highly effective frontal stunning procedure using a pneumatic penetrating captive bolt in water buffaloes. Animals 2023, 13, 177. [Google Scholar] [CrossRef]
- Nielsen, S.S.; Alvarez, J.; Bicout, D.J.; Calistri, P.; Depner, K.; Drewe, J.A.; Garin-Bastuji, B.; Gonzales Rojas, J.L.; Schmidt, C.G.; Michel, V.; et al. Welfare of cattle at slaughter. EFSA J. 2020, 18, e06275. [Google Scholar] [CrossRef]
- Gregory, N.G.; Lee, C.J.; Widdicombe, J.P. Depth of concussion in cattle shot by penetrating captive bolt. Meat Sci. 2007, 77, 499–503. [Google Scholar] [CrossRef]
- Kamenik, J.; Paral, V.; Pyszko, M.; Voslarova, E. Cattle stunning with a penetrative captive bolt device: A review. Anim. Sci. J. 2019, 90, 307–316. [Google Scholar] [CrossRef]
- Oliveira, S.E.O.; Gregory, N.G.; Dalla Costa, F.A.; Gibson, T.J.; Paranhos da Costa, M.J.R. Efficiency of low versus high airline pressure in stunning cattle with a pneumatically powered penetrating captive bolt gun. Meat Sci. 2017, 130, 64–68. [Google Scholar] [CrossRef]
- Lamont, L.A.; Tranquilli, W.J.; Grimm, K.A. Physiology of pain. Vet. Clin. North Am. Small Anim. Pract. 2000, 30, 703–728. [Google Scholar] [CrossRef] [PubMed]
- Sneddon, L.U. Comparative physiology of nociception and pain. Physiology 2018, 33, 63–73. [Google Scholar] [CrossRef]
- Bell, A. The Neurobiology of acute pain. Vet. J. 2018, 237, 55–62. [Google Scholar] [CrossRef]
- Hernández-Avalos, I.; Mota Rojas, D.; Mendoza-Flores, J.E.; Casas-Alvarado, A.; Flores-Padilla, K.; Miranda-Cortes, A.E.; Torres-Bernal, F.; Gómez-Prado, J.; Mora-Medina, P. Nociceptive pain and axiety in equines: Physiological and behavioral alterations. Vet. World 2021, 14, 2984–2995. [Google Scholar] [CrossRef]
- Velarde, A.; Dalmau, A. Animal welfare assessment at slaughter in Europe: Moving from inputs to outputs. Meat Sci. 2012, 92, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.; Mellor, D.; Hemsworth, P.; Fisher, A. A scientific comment on the welfare of domesticated ruminants slaughtered without stunning. N. Z. Vet. J. 2015, 63, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.S.; Alvarez, J.; Bicout, D.J.; Calistri, P.; Depner, K.; Drewe, J.A.; Garin-Bastuji, B.; Gonzales, J.L.; Gortázar, C.; Michel, V.; et al. Welfare of pigs at slaughter. EFSA J. 2020, 18, e06148. [Google Scholar] [CrossRef] [PubMed]
- Supratikno, S.; Setijanto, H.; Nuraini, H.; Agungpriyono, S. Bleeding time and false aneurysm incidence on cattle slaughtering using non-penetrative pre-slaughter stunning in Indonesia. Trop. Anim. Sci. J. 2022, 45, 482–489. [Google Scholar] [CrossRef]
- Johnson, C.; Gibson, T.; Stafford, K.; Mellor, D. Pain perception at slaughter. Anim. Welf. 2012, 21, 113–122. [Google Scholar] [CrossRef]
- von Holleben, K.; von Wenzlawowicz, N.; Gregory, H.; Anil, A.; Velarde, P.; Rodriguez, B.; Cenci Goga, B.; Catanese, B.; Lambooij, E. Report on Good and Adverse Practices-Animal Welfare Concerns in Relation to Slaughter Practices from the Viewpoint of Veterinary Sciences. Available online: https://web.archive.org/web/20121104005103/http://ec.europa.eu/research/biosociety/inco/pdf/ssa/dialrel.pdf (accessed on 21 May 2023).
- Amakiri, S.F.; Ozoya, S.E.; Ogunnaike, P.O. Nerves and nerve endings in the skin of tropical cattle. Cells Tissues Organs 1978, 100, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Sabow, A.B.; Goh, Y.M.; Zulkifli, I.; Sazili, A.Q.; Ab Kadir, M.Z.A.; Kaka, U.; Khadijah, N.; Adeyemi, K.D.; Ebrahimi, M. Electroencephalographic responses to neck cut and exsanguination in minimally anaesthetized goats. S. Afr. J. Anim. Sci. 2017, 47, 34. [Google Scholar] [CrossRef]
- Muir, W.W. Physiology and pathopjysiology of pain. In Handbook of Veterinary Pain Management; Gaynor, J.S., Muir, W.W., Eds.; Mosby: St. Louis, MI, USA, 2009; Volume 1, pp. 13–49. [Google Scholar]
- Wiese, A.J.; Yaksh, T.L. Nociception and pain mechanism. In Handbook of Veterinary Pain Management; Gaynor, J.S., Muir, W.W., Eds.; Elsevier: Alpharetta, GA, USA, 2009. [Google Scholar]
- McArthur, J.C.; Stocks, E.A.; Hauer, P.; Cornblath, D.R.; Griffin, J.W. Epidermal nerve fiber density. Arch. Neurol. 1998, 55, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- Habig, K.; Schänzer, A.; Schirner, W.; Lautenschläger, G.; Dassinger, B.; Olausson, H.; Birklein, F.; Gizewski, E.R.; Krämer, H.H. Low threshold unmyelinated mechanoafferents can modulate pain. BMC Neurol. 2017, 17, 184. [Google Scholar] [CrossRef] [PubMed]
- Gibson, T.; Johnson, C.; Murrell, J.; Mitchinson, S.; Stafford, K.; Mellor, D. Electroencephalographic responses to concussive non-penetrative captive-bolt stunning in halothane-anaesthetised calves. N. Z. Vet. J. 2009, 57, 90–95. [Google Scholar] [CrossRef]
- Gibson, T.; Johnson, C.; Murrell, J.; Chambers, J.; Stafford, K.; Mellor, D. Amelioration of electroencephalographic responses to slaughter by non-penetrative captive-bolt stunning after ventral-neck incision in halothane-anaesthetised calves. N. Z. Vet. J. 2009, 57, 96–101. [Google Scholar] [CrossRef]
- Raghazli, R.; Othman, A.-H.; Kaka, U.; Abubakar, A.A.; Imlan, J.C.; Hamzah, H.; Sazili, A.Q.; Goh, Y.-M. Physiological and electroencephalogram responses in goats subjected to pre-and during slaughter stress. Saudi J. Biol. Sci. 2021, 28, 6396–6407. [Google Scholar] [CrossRef]
- Momin, A.; McNaughton, P.A. Regulation of firing frequency in nociceptive neurons by pro-inflammatory mediators. Exp. Brain Res. 2009, 196, 45–52. [Google Scholar] [CrossRef]
- Meyer, R.A.; Ringkamp, M.; Campbell, J.N.; Raja, S.N. Peripheral mechanisms of cutaneous nociception. In Wall and Melzack’s Textbook of Pain; McMahon, S.B., Koltzenburg, M., Eds.; Elsevier: London, UK, 2006; pp. 3–34. [Google Scholar]
- Imbe, H. Stress-induced hyperalgesia: Animal models and putative mechanisms. Front. Biosci. 2006, 11, 2179–2192. [Google Scholar] [CrossRef]
- Hultgren, J.; Wiberg, S.; Berg, C.; Cvek, K.; Lunner Kolstrup, C. Cattle behaviours and stockperson actions related to impaired animal welfare at swedish slaughter plants. Appl. Anim. Behav. Sci. 2014, 152, 23–37. [Google Scholar] [CrossRef]
- Villarroel, M.; Maria, G.A.; Sierra, I.; Saitudo, C.; Garcia-Belenguer, S.; Gebresenbet, G. Critical points in the transport of cattle to slaughter in spain that may compromise the animals welfare. Vet. Rec. 2001, 149, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Abubakar, A.A.; Ahmed, M.A.; Hayat, M.N.; Ajat, M.; Kaka, U.; Goh, Y.M.; Sazili, A.Q. Electroencephalogram and physiological responses as affected by slaughter empathy in goats. Animals 2023, 13, 1100. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Napolitano, F.; Strappini, A.; Orihuela, A.; Martínez-Burnes, J.; Hernández-Ávalos, I.; Mora-Medina, P.; Velarde, A. Quality of death in fighting bulls during bullfights: Neurobiology and physiological responses. Animals 2021, 11, 2820. [Google Scholar] [CrossRef]
- Kober, A.; Bari, M.; Rakib, M.; Ali, M. Injuries of cattle and buffaloes during transportation and slaughter at Chittagong City corporation of Bangladesh. Bangladesh J. Anim. Sci. 2014, 43, 74–77. [Google Scholar] [CrossRef]
- Ossipov, M.H.; Dussor, G.O.; Porreca, F. Central modulation of pain. J. Clin. Investig. 2010, 120, 3779–3787. [Google Scholar] [CrossRef]
- Glardon, M.; Schwenk, B.K.; Riva, F.; von Holzen, A.; Ross, S.G.; Kneubuehl, B.P.; Stoffel, M.H. Energy loss and impact of various stunning devices used for the slaughtering of water buffaloes. Meat Sci. 2018, 135, 159–165. [Google Scholar] [CrossRef]
- Oliveira, S.E.O.; Gregory, N.G.; Dalla Costa, F.A.; Gibson, T.J.; Dalla Costa, O.A.; Paranhos da Costa, M.J.R. Effectiveness of pneumatically powered penetrating and non-penetrating captive bolts in stunning cattle. Meat Sci. 2018, 140, 9–13. [Google Scholar] [CrossRef]
- Gascho, D.; Stephan, R.; Bauer, C.; Oesch, M.A.; Richter, H. BigBovid-evaluation of a newly developed 9 mm bullet-shooting stunner for adequate stunning of heavy cattle. Front. Vet. Sci. 2022, 9, 949198. [Google Scholar] [CrossRef] [PubMed]
- Le Neindre, P.; Bernard, E.; Boissy, A.; Boivin, X.; Calandreau, L.; Delon, N.; Deputte, B.; Desmoulin-Canselier, S.; Dunier, M.; Faivre, N.; et al. Animal consciousness. EFSA Support. Publ. 2017, 14, 87. [Google Scholar] [CrossRef]
- Terlouw, C.; Bourguet, C.; Deiss, V. Consciousness, Unconsciousness and death in the context of slaughter. Part I. Neurobiological mechanisms underlying stunning and killing. Meat Sci. 2016, 118, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Strappini, A.; Ghezzi, M.D.; Hernández-Ávalos, I.; Rosmini, M.R.; Ramírez-Bribiesca, E.; Miranda-Cortés, A.; Cruz-Monterrosa, R.; Casas-Alvarado, A.; Rayas-Amor, A.; et al. ¿Cómo evaluar la calidad de muerte en búfalos y reses? Aspectos neurobiológicos. In El Búfalo de Agua en Latinoamérica, Hallazgos Recientes, 3rd ed.; Napolitano, F., Mota-Rojas, D., Guerrero-Legarreta, I., Orihuela, A., Eds.; BM Editores: Mexico City, México, 2020; pp. 1116–1177. [Google Scholar]
- Napolitano, F.; Mota-Rojas, D.; Guerrero-Legarreta, I.; Orihuela, A. El Búfalo de Agua En Latinoamérica, Hallazgos Recientes, 3rd ed.; BM Editores: Mexico City, México, 2020. [Google Scholar]
- Grandin, T. Auditing animal welfare at slaughter plants. Meat Sci. 2010, 86, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Miele, M. Killing Animals for Food: How science, religion and technologies affect the public debate about religious slaughter. Food Ethics 2016, 1, 47–60. [Google Scholar] [CrossRef]
- Atkinson, S.; Velarde, A.; Algers, B. Assessment of stun quality at commercial slaughter in cattle shot with captive bolt. Anim. Welf. 2013, 22, 473–481. [Google Scholar] [CrossRef]
- Terlouw, C.; Bourguet, C.; Deiss, V. Consciousness, unconsciousness and death in the context of slaughter. Part II. Evaluation methods. Meat Sci. 2016, 118, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Terlouw, C.E.M.; Bourguet, C.; Deiss, V.; Mallet, C. Origins of movements following stunning and during bleeding in cattle. Meat Sci. 2015, 110, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, M.T.W.; Gerritzen, M.A.; Hellebrekers, L.J.; Kemp, B. Indicators used in livestock to assess unconsciousness after stunning: A review. Animal 2015, 9, 320–330. [Google Scholar] [CrossRef]
- Comin, M.; Barbieri, S.; Minero, M.; Dalla Costa, E. The feasibility of animal-based indicators of consciousness and unconsciousness for stunning in sheep: A systematic review. Animals 2023, 13, 1395. [Google Scholar] [CrossRef]
- Mills, E.P.; Combs-Ramey, K.; Kwong, G.P.S.; Pang, D.S.J. Development of reference intervals for pupillometry in healthy dogs. Front. Vet. Sci. 2022, 9, 1020710. [Google Scholar] [CrossRef]
- Larson, M.D.; Behrends, M. Portable infrared pupillometry. Anesth. Analg. 2015, 120, 1242–1253. [Google Scholar] [CrossRef]
- Packiasabapathy, S.; Rangasamy, V.; Sadhasivam, S. Pupillometry in perioperative medicine: A narrative review. Can. J. Anaesth. 2021, 68, 566–578. [Google Scholar] [CrossRef] [PubMed]
- Imlan, J.C.; Kaka, U.; Goh, Y.-M.; Idrus, Z.; Awad, E.A.; Abubakar, A.A.; Ahmad, T.; Nizamuddin, H.N.Q.; Sazili, A.Q. Effects of Slaughter Knife Sharpness on Blood Biochemical and Electroencephalogram Changes in Cattle. Animals 2020, 10, 579. [Google Scholar] [CrossRef] [PubMed]
- Gajkowska, B.; Kosson, A.; Sacharczuk, M.; Kosson, P.; Lipkowski, A.W. Blood-brain barrier permeability differentiates Sadowski mouse lines of high and low stress-induced analgesia. Electron microscopy analysis. Folia Neuropathol. 2011, 49, 311–318. [Google Scholar] [PubMed]
- Gregory, N.G.; Schuster, P.; Mirabito, L.; Kolesar, R.; McManus, T. Arrested blood flow during false aneurysm formation in the carotid arteries of cattle slaughtered with and without stunning. Meat Sci. 2012, 90, 368–372. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grandin, T.; Velarde, A.; Strappini, A.; Gerritzen, M.; Ghezzi, M.; Martínez-Burnes, J.; Hernández-Ávalos, I.; Domínguez-Oliva, A.; Casas-Alvarado, A.; Mota-Rojas, D. Slaughtering of Water Buffalo (Bubalus bubalis) with and without Stunning: A Focus on the Neurobiology of Pain, Hyperalgesia, and Sensitization. Animals 2023, 13, 2406. https://doi.org/10.3390/ani13152406
Grandin T, Velarde A, Strappini A, Gerritzen M, Ghezzi M, Martínez-Burnes J, Hernández-Ávalos I, Domínguez-Oliva A, Casas-Alvarado A, Mota-Rojas D. Slaughtering of Water Buffalo (Bubalus bubalis) with and without Stunning: A Focus on the Neurobiology of Pain, Hyperalgesia, and Sensitization. Animals. 2023; 13(15):2406. https://doi.org/10.3390/ani13152406
Chicago/Turabian StyleGrandin, Temple, Antonio Velarde, Ana Strappini, Marien Gerritzen, Marcelo Ghezzi, Julio Martínez-Burnes, Ismael Hernández-Ávalos, Adriana Domínguez-Oliva, Alejandro Casas-Alvarado, and Daniel Mota-Rojas. 2023. "Slaughtering of Water Buffalo (Bubalus bubalis) with and without Stunning: A Focus on the Neurobiology of Pain, Hyperalgesia, and Sensitization" Animals 13, no. 15: 2406. https://doi.org/10.3390/ani13152406
APA StyleGrandin, T., Velarde, A., Strappini, A., Gerritzen, M., Ghezzi, M., Martínez-Burnes, J., Hernández-Ávalos, I., Domínguez-Oliva, A., Casas-Alvarado, A., & Mota-Rojas, D. (2023). Slaughtering of Water Buffalo (Bubalus bubalis) with and without Stunning: A Focus on the Neurobiology of Pain, Hyperalgesia, and Sensitization. Animals, 13(15), 2406. https://doi.org/10.3390/ani13152406