


Feeding Flaxseed and Lupins during the Transition Period in Dairy Cows: Effects on Production Performance, Fertility and Biochemical Blood Indices
 , ,
, ,  , , , , ,
, , , , ,  and
and (This article belongs to the Section Animal Products)
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Guidelines of the Animal Research
2.2. Animals and Experimetal Design
2.3. Physical Evaluations and Clinical Examinations
2.4. Determination of Milk Yield and Composition
2.5. Hematological and Biochemical Examinations
2.6. Uterine Examinations
2.7. Ovarian Resumption, Estrus Expression and Pregnancy Diagnosis
2.8. Endometrial Cytology
2.9. Statistical Analysis
3. Results
3.1. Nutrotional Values, Body Condition Scoring and Milk Yield and Composition
3.2. Incidence of Diseases and Ovarian Resumption, Days to Conception
3.3. Serum TBARS and Catalase Activity
3.4. Serum NEFA, Hpt and SAA Values
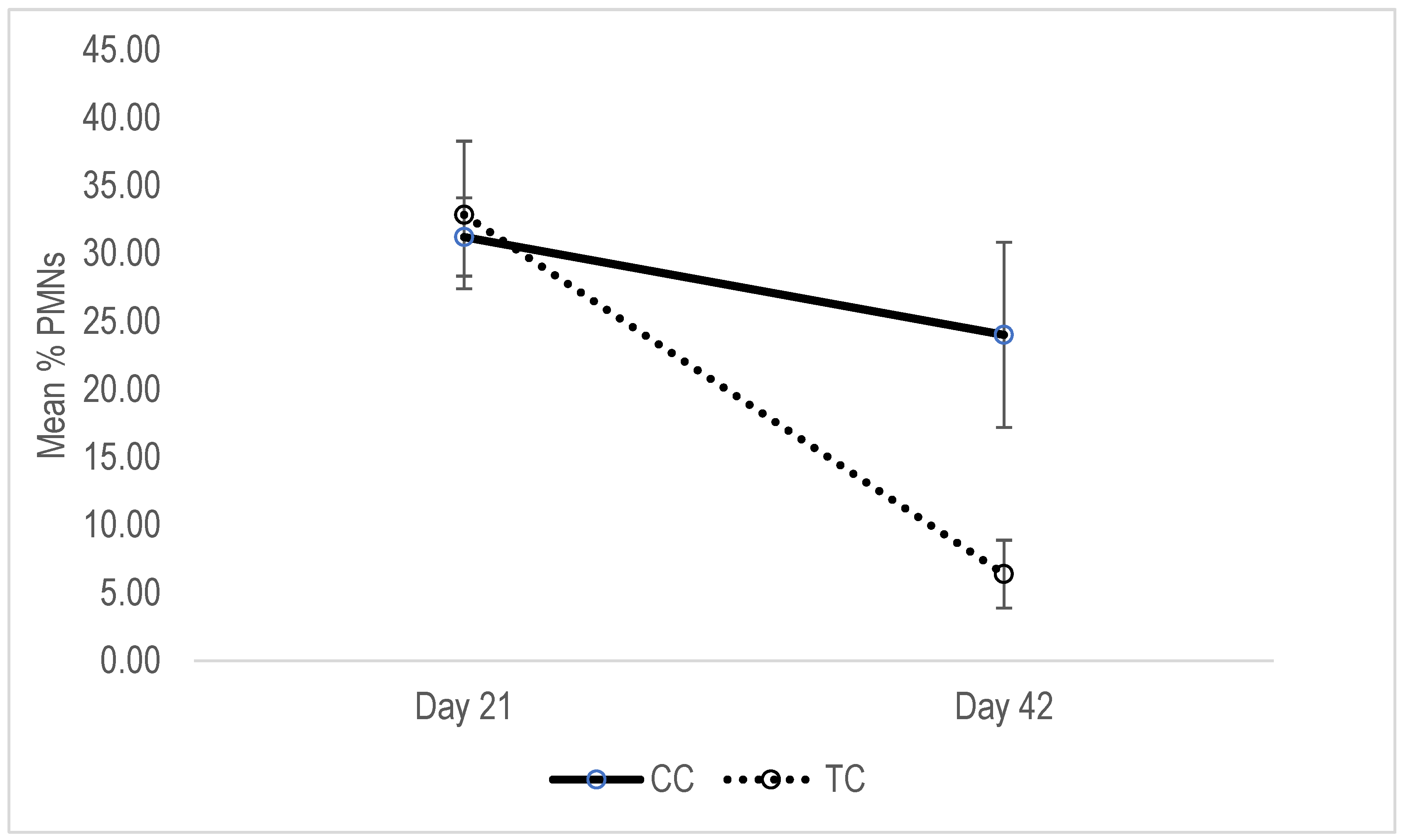
3.5. Cytology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Noziere, P.; Sauvant, D.; Delaby, L. INRA Feeding System for Ruminants; Wageningen Academic Publishers: Wageningen, The Netherlands, 2018; 640p. [Google Scholar]
- De Visser, C.L.M.; Schreuder, R.; Stoddard, F. The EU’s dependency on soya bean import for the animal feed industry and potential for EU produced alternatives. Oilseeds Fats Crops Lipids 2014, 21, 4. [Google Scholar] [CrossRef]
- Niavis, S.; Kleisiari, C.; Kyrgiakos, L.-S.; Vlontzos, G. Assessing the sustainability of legumes production in South Europe. In Bio-Economy and Agri-Production; Academic Press: Cambridge, MA, USA, 2021; pp. 117–129. [Google Scholar]
- Britt, J.H.; Cushman, R.A.; Dechow, C.D.; Dobson, H.; Humblot, P.; Hutjens, M.F.; Sheldon, I.M.; Stevenson, J.S. Invited review: Learning from the future—A vision for dairy farms and cows in 2067. J. Dairy Sci. 2018, 101, 3722–3741. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.F.; Chouinard, P.Y.; Christensen, D.A. Effects of feeding micronised flaxseed on yield and composition of milk from Holstein cows. J. Sci. Food Agric. 2003, 83, 920–926. [Google Scholar] [CrossRef]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 Fatty Acids EPA and DHA: Health Benefits throughout Life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zachut, M.; Arieli, A.; Lehrer, H.; Livshitz, L.; Yakoby, S.; Moallem, U. Effects of increased supplementation of n-3 fatty acids to transition dairy cows on performance and fatty acid profile in plasma, adipose tissue, and milk fat. J. Dairy Sci. 2010, 93, 5877–5889. [Google Scholar] [CrossRef]
- Yoshimura, E.H.; Santos, N.W.; Machado, E.; Agustinho, B.C.; Pereira, L.M.; De Aguiar, S.C.; Zeoula, L.M. Effects of dairy cow diets supplied with flaxseed oil and propolis extract, with or without vitamin E, on the ruminal microbiota, biohydrogenation, and digestion. Anim. Feed. Sci. Technol. 2018, 241, 163–172. [Google Scholar] [CrossRef]
- White, C.L.; Staines, V.E. A review of the nutritional value of lupins for dairy cows. Aust. J. Agric. Res. 2007, 58, 185–202. [Google Scholar] [CrossRef]
- Fernández-Pascual, M.; Pueyo, J.J.; Felipe, M.; Golvano, M.P.; Lucas, M.M. Singular features of the Bradyrhizobium-Lupinus symbiosis. Dyn. Soil Dyn. Plant 2007, 1, 1–16. [Google Scholar]
- Coba de la Pena, T.; Pueyo, J.J. Legumes in the reclamation of marginal soils, from cultivar and inoculant selection to transgenic approaches. Agron. Sustain. Dev. 2012, 32, 65–91. [Google Scholar] [CrossRef]
- Joch, M.; Kudrna, V. Partial replacement of soybean meal by white lupine seeds in the diet of dairy cows. Asian-Australas. J. Anim. Sci. 2020, 33, 957–964. [Google Scholar] [CrossRef]
- Caixeta, L.S.; Omontese, B.O. Monitoring and Improving the Metabolic Health of Dairy Cows during the Transition Period. Animals 2021, 11, 352. [Google Scholar] [CrossRef] [PubMed]
- Kerwin, A.L.; Burhans, W.S.; Mann, S.; Tetreault, M.; Nydam, D.V.; Overton, T.R. Transition cow nutrition and management strategies of dairy herds in the northeastern United States: Part II—Associations of metabolic- and inflammation-related analytes with health, milk yield, and reproduction. J. Dairy Sci. 2022, 105, 5349–5369. [Google Scholar] [CrossRef] [PubMed]
- Wankhade, P.R.; Manimaran, A.; Kumaresan, A.; Jeyakumar, S.; Ramesha, K.P.; Sejian, V.; Varghese, M.R. Metabolic and immunological changes in transition dairy cows: A review. Vet. World 2017, 10, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Drackley, J.; Cardoso, F. Prepartum and postpartum nutritional management to optimize fertility in high-yielding dairy cows in confined TMR systems. Animal 2014, 8, 5–14. [Google Scholar] [CrossRef]
- Directive 2010/63/EU of the European Parliament and of the Council on the protection of animals used for scientific purposes. Off. J. Eur. Union 2010, 276, 33–79.
- Edmonson, A.J.; Lean, I.J.; Weaver, L.D.; Farver, T.; Webster, G. A Body Condition Scoring Chart for Holstein Dairy Cows. J. Dairy Sci. 1989, 72, 68–78. [Google Scholar] [CrossRef]
- Nanas, I.; Chouzouris, T.-M.; Dovolou, E.; Dadouli, K.; Stamperna, K.; Kateri, I.; Barbagianni, M.; Amiridis, G.S. Early embryo losses, progesterone and pregnancy associated glycoproteins levels during summer heat stress in dairy cows. J. Therm. Biol. 2021, 98, 102951. [Google Scholar] [CrossRef]
- Ahn, D.U.; Olson, D.G.; Jo, C.; Love, J.; Jin, S.K. Volatiles Production and Lipid Oxidation in Irradiated Cooked Sausage as Related to Packaging and Storage. J. Food Sci. 1999, 64, 226–229. [Google Scholar] [CrossRef]
- Góth, L.A. Simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar] [CrossRef]
- Gilbert, R.O.; Shin, S.T.; Guard, C.L.; Erb, H.N.; Frajblat, M. Prevalence of endometritis and its effects on reproductive performance of dairy cows. Theriogenology 2005, 64, 1879–1888. [Google Scholar] [CrossRef]
- Melcher, Y.; Prunner, I.; Drillich, M. Degree of variation and reproducibility of different methods for the diagnosis of subclinical endometritis. Theriogenology 2014, 82, 57–63. [Google Scholar] [CrossRef]
- Giannenas, I.; Bonos, E.; Anestis, V.; Filioussis, G.; Papanastasiou, D.K.; Bartzanas, T.; Papaioannou, N.; Tzora, A.; Skoufos, I. Effects of Protease Addition and Replacement of Soybean Meal by Corn Gluten Meal on the Growth of Broilers and on the Environmental Performances of a Broiler Production System in Greece. PLoS ONE 2017, 12, e0169511. [Google Scholar] [CrossRef] [PubMed]
- Caroprese, M.; Marzano, A.; Marino, R.; Gliatta, G.; Muscio, A.; Sevi, A. Flaxseed supplementation improves fatty acid profile of cow milk. J. Dairy Sci. 2010, 93, 2580–2588. [Google Scholar] [CrossRef] [PubMed]
- Zachut, M.; Arieli, A.; Moallem, U. Incorporation of dietary n-3 fatty acids into ovarian compartments in dairy cows and the effects on hormonal and behavioral patterns around estrus. Reproduction 2011, 141, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Neveu, C.; Baurhoo, B.; Mustafa, A. Effect of feeding extruded flaxseed with different forage:concentrate ratios on the performance of dairy cows. J. Dairy Sci. 2013, 96, 3886–3894. [Google Scholar] [CrossRef] [PubMed]
- Petit, H.V.; Palin, M.F.; Doepel, L. Hepatic Lipid Metabolism in Transition Dairy Cows Fed Flaxseed. J. Dairy Sci. 2007, 90, 4780–4792. [Google Scholar] [CrossRef]
- Brossillon, V.; Reis, S.F.; Moura, D.C.; Galvão, J.G.B., Jr.; Oliveira, A.S.; Côrtes, C.; Brito, A.F. Production, milk and plasma fatty acid profile, and nutrient utilization in Jersey cows fed flaxseed oil and corn grain with different particle size. J. Dairy Sci. 2018, 101, 2127–2143. [Google Scholar] [CrossRef]
- Moallem, U. Invited review: Roles of dietary n-3 fatty acids in performance, milk fat composition, and reproductive and immune systems in dairy cattle. J. Dairy Sci. 2018, 101, 8641–8661. [Google Scholar] [CrossRef]
- Hurtaud, C.; Faucon, F.; Couvreur, S.; Peyraud, J.-L. Linear relationship between increasing amounts of extruded linseed in dairy cow diet and milk fatty acid composition and butter properties. J. Dairy Sci. 2010, 93, 1429–1443. [Google Scholar] [CrossRef]
- Ferlay, A.; Doreau, M.; Martin, C.; Chilliard, Y. Effects of incremental amounts of extruded linseed on the milk fatty acid composition of dairy cows receiving hay or corn silage. J. Dairy Sci. 2013, 96, 6577–6595. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Corl, B.A.; Dwyer, D.A.; Sæbø, A.; Bauman, D.E. Identification of the conjugated linoleic acid isomer that inhibits milk fat synthesis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, 179–184. [Google Scholar] [CrossRef]
- Akraim, F.; Nicot, M.C.; Juaneda, P.; Enjalbert, F. Conjugated linolenic acid (CLnA), conjugated linoleic acid (CLA) and other biohydrogenation intermediates in plasma and milk fat of cows fed raw or extruded linseed. Animal 2007, 1, 835–843. [Google Scholar] [CrossRef]
- Mendowski, S.; Chapoutot, P.; Chesneau, G.; Ferlay, A.; Enjalbert, F.; Cantalapiedra-Hijar, G.; Germain, A.; Nozière, P. Effects of replacing soybean meal with raw or extruded blends containing faba bean or lupin seeds on nitrogen metabolism and performance of dairy cows. J. Dairy Sci. 2019, 102, 5130–5147. [Google Scholar] [CrossRef]
- Robinson, P.H.; McNiven, M.A. Nutritive value of raw and roasted sweet white lupins (Lupinus albus) for lactating dairy cows. Anim. Feed. Sci. Technol. 1993, 43, 275–290. [Google Scholar] [CrossRef]
- Bayourthe, C.; Moncoulon, R.; Enjalbert, F. Effect of extruded lupin seeds as a protein source on lactational performance of dairy cows. Anim. Feed. Sci. Technol. 1998, 72, 121–131. [Google Scholar] [CrossRef]
- Ambrose, D.J.; Kastelic, J.P.; Corbett, R.; Pitney, P.A.; Petit, H.V.; Small, J.A.; Zalkovic, P. Lower Pregnancy Losses in Lactating Dairy Cows Fed a Diet Enriched in α-Linolenic Acid. J. Dairy Sci. 2006, 89, 3066–3074. [Google Scholar] [CrossRef]
- Robinson, R.S.; Pushpakumara, P.G.; Cheng, Z.; Peters, A.R.; Abayasekara, D.R.; Wathes, D.C. Effects of dietary polyunsaturated fatty acids on ovarian and uterine function in lactating dairy cows. Reproduction 2002, 124, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Petit, H.V.; Dewhurst, R.J.; Proulx, J.G.; Khalid, M.; Haresign, W.; Twagiramungu, H. Milk production, milk composition, and reproductive function of dairy cows fed different fats. Can. J. Anim. Sci. 2001, 81, 263–271. [Google Scholar] [CrossRef]
- Ulfina, G.G.; Kimothi, S.P.; Oberoi, P.S.; Baithalu, R.K.; Kumaresan, A.; Mohanty, T.K.; Imtiwati, P.; Dang, A.K. Modulation of post-partum reproductive performance in dairy cows through supplementation of long or short-chain fatty acids during transition period. J. Anim. Physiol. Anim. Nutr. 2015, 99, 1056–1064. [Google Scholar] [CrossRef]
- Jahani-Moghadam, E.; Mahjoubi, E.; Dirandeh, E. Effect of linseed feeding on blood metabolites, incidence of cystic follicles, and productive and reproductive performance in fresh Holstein dairy cows. J. Dairy Sci. 2015, 98, 1828–1835. [Google Scholar] [CrossRef]
- Chouzouris, T.M.; Dovolou, E.; Georgoulias, P.; Rekkas, A.; Dafopoulos, K.; Athanasiou, L.; Fthenakis, G.C.; Amiridis, G.S. Effects of pregnancy and short-lasting acute feed restriction on total ghrelin concentration and metabolic parameters in dairy cattle. Theriogenology 2018, 106, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Gautam, G.; Nakao, T.; Yamada, K.; Yoshida, C. Defining delayed resumption of ovarian activity postpartum and its impact on subsequent reproductive performance in Holstein cows. Theriogenology 2010, 73, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Chagas, L.M.; Bass, J.J.; Blache, D.; Burke, C.; Kay, J.; Lindsay, D.; Lucy, M.; Martin, G.; Meier, S.; Rhodes, F.; et al. Invited Review: New Perspectives on the Roles of Nutrition and Metabolic Priorities in the Subfertility of High-Producing Dairy Cows. J. Dairy Sci. 2007, 90, 4022–4032. [Google Scholar] [CrossRef]
- Bobe, G.; Young, J.W.; Beitz, D.C. Invited Review: Pathology, Etiology, Prevention, and Treatment of Fatty Liver in Dairy Cows. J. Dairy Sci. 2004, 87, 3105–3124. [Google Scholar] [CrossRef] [PubMed]
- Garnsworthy, P.C.; Gong, J.G.; Armstrong, D.G.; Newbold, J.R.; Marsden, M.; Richards, S.E.; Mann, G.E.; Sinclair, K.D.; Webb, R. Nutrition, Metabolism, and Fertility in Dairy Cows: 3. Amino Acids and Ovarian Function. J. Dairy Sci. 2008, 91, 4190–4197. [Google Scholar] [CrossRef] [PubMed]
- Somchit, A.; Campbell, B.K.; Khalid, M.; Kendall, N.R.; Scaramuzzi, R.J. The effect of short-term nutritional supplementation of ewes with lupin grain (Lupinus luteus), during the luteal phase of the estrous cycle on the number of ovarian follicles and the concentrations of hormones and glucose in plasma and follicular fluid. Theriogenology 2007, 68, 1037–1046. [Google Scholar] [CrossRef]
- Petit, H.V.; Benchaar, C. Milk production, milk composition, blood composition, and conception rate of transition dairy cows fed different fat sources. Can. J. Anim. Sci. 2007, 87, 591–600. [Google Scholar] [CrossRef]
- Petit, H.V.; Twagiramungu, H. Conception rate and reproductive function of dairy cows fed different fat sources. Theriogenology 2006, 66, 1316–1324. [Google Scholar] [CrossRef]
- Shin, D.H.; Jeong, J.K.; Choi, I.S.; Moon, S.H.; Lee, S.C.; Kang, H.G.; Park, S.B.; Kim, I.H. Associations between serum haptoglobin concentration and peri- and postpartum disorders, milk yield, and reproductive performance in dairy cows. Livest. Sci. 2018, 213, 14–18. [Google Scholar] [CrossRef]
- Chan, J.P.W.; Chang, C.-C.; Hsu, W.-L.; Liu, W.-B.; Chen, T.-H. Association of increased serum acute-phase protein concentrations with reproductive performance in dairy cows with postpartum metritis. Vet. Clin. Pathol. 2010, 39, 72–78. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Dobson, H. Postpartum uterine health in cattle. Anim. Reprod. Sci. 2004, 82–83, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Regassa, F.; Noakes, D.E. Acute phase protein response of ewes and tge release of PGFM in relation to uterine involution and the presence of intrauterine bacteria. Vet. Rec. 1999, 144, 502–506. [Google Scholar] [CrossRef] [PubMed]
- Tóthová, C.; Nagy, O.; Kováč, G. Relationship between some variables of protein profile and indicators of lipomobilization in dairy cows after calving. Archiv. Tierzucht. 2014, 57, 19. [Google Scholar] [CrossRef]
- Eckersal, P.D.; Bell, R. Acute phase proteins; biomarkers of infection and inflammation in veterinary medicine. Vet. J. 2010, 185, 23–27. [Google Scholar] [CrossRef]
- Ametaj, B.N.; Bradford, B.J.; Bobe, G.; Nafikov, R.A.; Lu, Y.; Young, J.W.; Beitz, D.C. Strong relationships between mediators of the acute phase response and fatty liver in dairy cows. Can. J. Anim. Sci. 2005, 85, 165–175. [Google Scholar] [CrossRef]
- Adewuyi, A.A.; Gruys, E.; van Eerdenburg, F.J.C.M. Non esterified fatty acids (NEFA) in dairy cattle. A review. Vet. Q. 2005, 27, 117–126. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Williams, E.J.; Miller, A.N.A.; Nash, D.M.; Herath, S. Uterine diseases in cattle after parturition. Vet. J. 2008, 176, 115–121. [Google Scholar] [CrossRef]
- Galvão, K.N.; Frajblat, M.; Brittin, S.; Butler, W.; Guard, C.; Gilbert, R. Effect of prostaglandin F2α on subclinical endometritis and fertility in dairy cows. J. Dairy Sci. 2009, 92, 4906–4913. [Google Scholar] [CrossRef]
- Kasimanickam, R.; Duffield, T.F.; Foster, R.A.; Gartley, C.J.; Leslie, K.E.; Walton, J.S.; Johnson, W.H. Endometrial cytology and ultrasonography for the detection of subclinical endometritis in postpartum dairy cows. Theriogenology 2004, 62, 9–23. [Google Scholar] [CrossRef]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Cellular and Molecular Immunology, 6th ed.; Elsevier Health Sciences: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Boschin, B.; Arnoldi, A. Legumes are valuable sources of tocopherols. Food Chem. 2011, 127, 1199–1203. [Google Scholar] [CrossRef]
- Kelley, D.S. Modulation of human immune and inflammatory responses by dietary fatty acids. Nutrition 2001, 17, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Dietary modification of inflammation with lipids. Proc. Nutr. Soc. 2002, 61, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Yaqoob, P.; Calder, P.C. The effect of dietary lipid manipulation on the production of murine T cell-derived cytokines. Cytokine 1995, 7, 548–553. [Google Scholar] [CrossRef]
- Lessard, M.; Gagnon, N.; Petit, H.V. Immune Response of Postpartum Dairy Cows Fed Flaxseed. J. Dairy Sci. 2003, 86, 2647–2657. [Google Scholar] [CrossRef] [PubMed]
- Lessard, M.; Gagnon, N.; Godson, L.; Petit, H.V. Influence of Parturition and Diets Enriched in n-3 or n-6 Polyunsaturated Fatty Acids on Immune Response of Dairy Cows during the Transition Period. J. Dairy Sci. 2004, 87, 2197–2210. [Google Scholar] [CrossRef]
- Caroprese, M.; Marzano, A.; Entrican, G.; Wattegedera, S.; Albenzio, M.; Sevi, A. Immune response of cows fed polyunsaturated fatty acids under high ambient temperatures. J. Dairy Sci. 2009, 92, 2796–2803. [Google Scholar] [CrossRef]
- Ingvartsen, K.L.; Moyes, K. Nutrition, immune function and health of dairy cattle. Animal 2013, 7, 112–122. [Google Scholar] [CrossRef]
- Hammon, D.S.; Evjen, I.M.; Dhiman, T.R.; Goff, J.P.; Walters, J.L. Neutrophil function and energy status in Holstein cows with uterine health disorders. Vet. Immunol. 2006, 113, 21–29. [Google Scholar] [CrossRef]
- Yasui, T.; McArt, J.A.A.; Ryan, C.M.; Gilbert, R.O.; Nydam, D.V.; Valdez, F.; Griswold, K.E.; Overton, T.R. Effects of chromium propionate supplementation during the periparturient period and early lactation on metabolism, performance, and cytological endometritis in dairy cows. J. Dairy Sci. 2014, 97, 6400–6410. [Google Scholar] [CrossRef]
- Brandão, A.P.; Cooke, R.F.; Corrá, F.N.; Piccolo, M.B.; Gennari, R.; Leiva, T.; Vasconcelos, J.L.M. Physiologic, health, and production responses of dairy cows supplemented with an immunomodulatory feed ingredient during the transition period. J. Dairy Sci. 2016, 99, 5562–5572. [Google Scholar] [CrossRef]
- Santos, N.W.; Yoshimura, E.H.; Machado, E.; Matumoto-Pintro, P.T.; Montanher, P.F.; Visentainer, J.V.; dos Santos, G.T.; Zeoula, L.M. Antioxidant effects of a propolis extract and vitamin E in blood and milk of dairy cows fed diet containing flaxseed oil. Livest. Sci. 2016, 191, 132–138. [Google Scholar] [CrossRef]
- Bu, D.P.; Wang, J.Q.; Dhiman, T.R.; Liu, S.J. Effectiveness of Oils Rich in Linoleic and Linolenic Acids to Enhance Conjugated Linoleic Acid in Milk from Dairy Cows. J. Dairy Sci. 2007, 90, 998–1007. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Diet | ||
|---|---|---|
| Item | Group C | Group T |
| Corn silage (kg/DM) | 6.90 | 6.90 |
| Alfalfa hay (kg/DM) | 2.43 | 2.43 |
| Wheat straw (kg/DM) | 2.00 | 2.00 |
| Corn (kg/DM) | 2.61 | 1.74 |
| Barley (kg/DM) | 1.05 | 0.87 |
| Soymeal (CP: 47%) (kg/DM) | 2.46 | 1.23 |
| Rapeseed meal (CP: 32%) (kg/DM) | 1.85 | 1.76 |
| Molasses (kg/DM) | 1.93 | 1.54 |
| White Lupin (kg/DM) | - | 1.08 |
| Flaxseed (kg/DM) | - | 1.08 |
| Mineral and vitamin premix 1 (kg/DM) | 0.65 | 0.65 |
| Chemical analysis 2 | ||
| DM (g) | 21.88 | 21.28 |
| CP% DM | 15.85 | 15.91 |
| ADF % DM | 18.52 | 22.20 |
| NDF % DM | 37.69 | 41.25 |
| Ether extract % DM | 7.55 | 10.04 |
| UFL/kg | 0.501 | 0.499 |
| Starch % DM | 17.00 | 14.50 |
| Sugars % DM | 8.90 | 7.90 |
| PDIA g/kg DM | 59.45 | 82.31 |
| PDI g/kg DM | 54.08 | 76.90 |
| RPB | 43.88 | 45.98 |
| Ca % DM | 0.33 | 0.31 |
| P % DM | 0.33 | 0.3 |
| Ca,absorb(g) | 28.1 | 27.4 |
| P,absorb(g) | 24.7 | 25.1 |
| Mg % DM | 0.19 | 0.18 |
| Na % DM | 0.10 | 0.09 |
| S % DM | 0.28 | 0.25 |
| Item | Group C 1 | Group T | SEM 2 | p-Value |
|---|---|---|---|---|
| Dry matter intake/day | ||||
| Multiparous cows | 21.30 | 21.25 | 0.085 | NS 3 |
| Primiparous cows | 19.40 | 19.45 | 0.078 | NS |
| Item | Group C 1 | Group T | SEM 2 | p-Value |
|---|---|---|---|---|
| Protein (%) | 3.37 | 3.39 | 0.170 | NS 3 |
| Fat (%) | 3.95 | 3.88 | 0.325 | NS |
| SCC (×1000) | 158.5 | 157.0 | 0.086 | NS |
| CFU (×1000) | 8.20 | 8.75 | 0.158 | NS |
| Type of Animals | Group C | Group T | SEM 2 | p-Value |
|---|---|---|---|---|
| All animals | 61.3 | 51.7 | 0.153 | NS 3 |
| Multiparous cows | 60.1 a | 47.1 b | 0.128 | * |
| Primiparous cows | 62.7 | 56.3 | 0.163 | NS |
| Type of Animals | Group C | Group T | SEM 2 | p-Value |
|---|---|---|---|---|
| All animals | 61.0 | 69.8 | 0.110 | NS 3 |
| Multiparous cows | 57.7 a | 70.9 b | 0.074 | * |
| Primiparous cows | 67.3 | 68 | 0.035 | NS |
| Item | Group C | Group T | SEM 2 | p-Value |
|---|---|---|---|---|
| Serum TBARS (ng/mL) | ||||
| Day 1 | 55.58 | 68.72 | 0.282 | NS 3 |
| Day 3 | 57.02 | 86.66 | 0.850 | NS |
| Serum catalase (nmol/min/mL) | 12.35 | 13.09 | 0.147 | NS |
| Item | CC | TC | CH | TH |
|---|---|---|---|---|
| Serum NEFA (mMol/L) | ||||
| Day −7 | 0.39 ± 0.28 | 0.42 ± 0.25 | 0.31 ± 0.18 | 0.34 ± 0.21 |
| Day 0 | 0.44 ± 0.36 | 0.49 ± 0.37 | 0.35 ± 0.16 | 0.39 ± 0.22 |
| Day 7 | 0.50 ± 0.34 | 0.30 ± 0.22 | 0.47 ± 0.26 | 0.41 ± 0.32 |
| Day 14 | 0.73 ± 0.20 * | 0.43 ± 0.23 | 0.55 ± 0.30 | 0.45 ± 0.29 |
| Day 21 | 0.56 ± 0.48 | 0.41 ± 0.29 | 0.56 ± 0.48 | 0.47 ± 0.30 |
| Day 42 | 0.38 ± 0.33 * | 0.21 ± 0.16 | 0.47 ± 0.23 | 0.39 ± 0.27 |
| Serum Haptoglobin (μg/mL) | ||||
| Day −7 | 52.7 ± 11.1 * | 35.4 ± 20.3 | 59.7 ± 9.8 * | 39.4 ± 5.7 |
| Day 7 | 26.4 ± 11.9 | 23.3 ± 3.4 | 34.5 ± 8.6 * | 23.3 ± 3.4 |
| Day 21 | 28.6 ± 19.4 | 26.9 ± 3.1 | 28.2 ± 5.5 | 26.9 ± 3.1 |
| Serum SAA (mg/L) | ||||
| Day −7 | 36.8 ± 7.6 | 22.9 ± 4.1 | 78.0 ± 14.6 * | 28.8 ±17.7 |
| Day 7 | 22.9 ± 8.9 | 15.2 ± 5.2 | 69.3 ± 18.6 | 63.6 ± 13.1 |
| Day 21 | 50.0 ± 10.4 | 30.6 ± 4.8 | 118.2 ± 22.5 | 85.5 ± 23.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nanas, I.; Dokou, S.; Athanasiou, L.V.; Dovolou, E.; Chouzouris, T.M.; Vasilopoulos, S.; Grigoriadou, K.; Giannenas, I.; Amiridis, G.S. Feeding Flaxseed and Lupins during the Transition Period in Dairy Cows: Effects on Production Performance, Fertility and Biochemical Blood Indices. Animals 2023, 13, 1972. https://doi.org/10.3390/ani13121972
Nanas I, Dokou S, Athanasiou LV, Dovolou E, Chouzouris TM, Vasilopoulos S, Grigoriadou K, Giannenas I, Amiridis GS. Feeding Flaxseed and Lupins during the Transition Period in Dairy Cows: Effects on Production Performance, Fertility and Biochemical Blood Indices. Animals. 2023; 13(12):1972. https://doi.org/10.3390/ani13121972
Chicago/Turabian StyleNanas, Ioannis, Stella Dokou, Labrini V. Athanasiou, Eleni Dovolou, Thomas M. Chouzouris, Stelios Vasilopoulos, Katerina Grigoriadou, Ilias Giannenas, and Georgios S. Amiridis. 2023. "Feeding Flaxseed and Lupins during the Transition Period in Dairy Cows: Effects on Production Performance, Fertility and Biochemical Blood Indices" Animals 13, no. 12: 1972. https://doi.org/10.3390/ani13121972
APA StyleNanas, I., Dokou, S., Athanasiou, L. V., Dovolou, E., Chouzouris, T. M., Vasilopoulos, S., Grigoriadou, K., Giannenas, I., & Amiridis, G. S. (2023). Feeding Flaxseed and Lupins during the Transition Period in Dairy Cows: Effects on Production Performance, Fertility and Biochemical Blood Indices. Animals, 13(12), 1972. https://doi.org/10.3390/ani13121972