
Influence of Media Composition on the Level of Bovine Satellite Cell Proliferation
 , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Muscle Satellite Cells
2.2. Selection of the Components of the Tested Media
2.3. Cell Culture and Estimation of Proliferation Rate and Visualisation
2.4. Statistical Analysis
2.5. Analysis of the Expression of Genes of Myogenesis
2.6. Statistical Analysis of the Expression of Genes
3. Results
3.1. Full Factorial Design of Tested Components
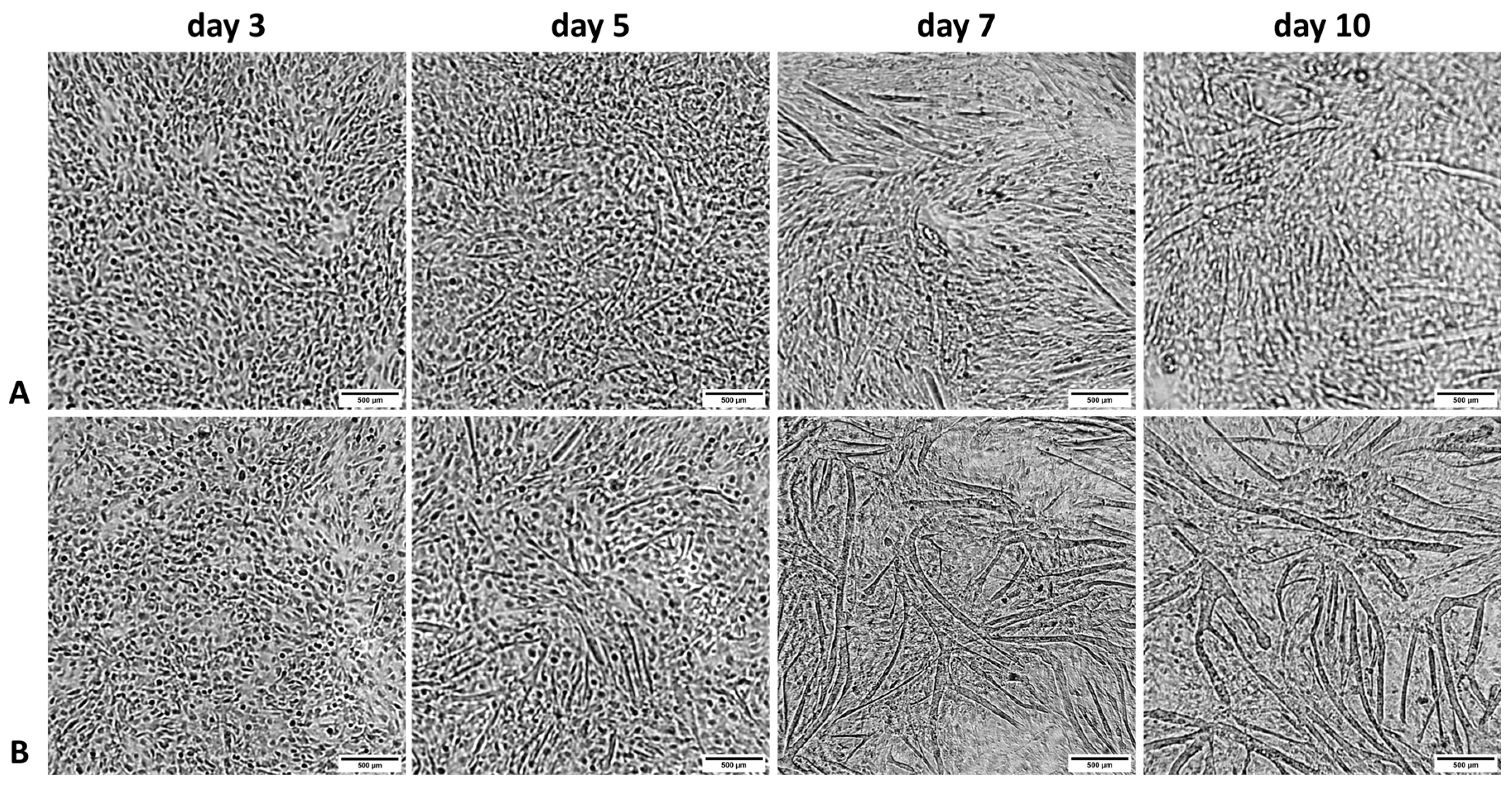
3.2. Microscopic Observations of Proliferating BSCs
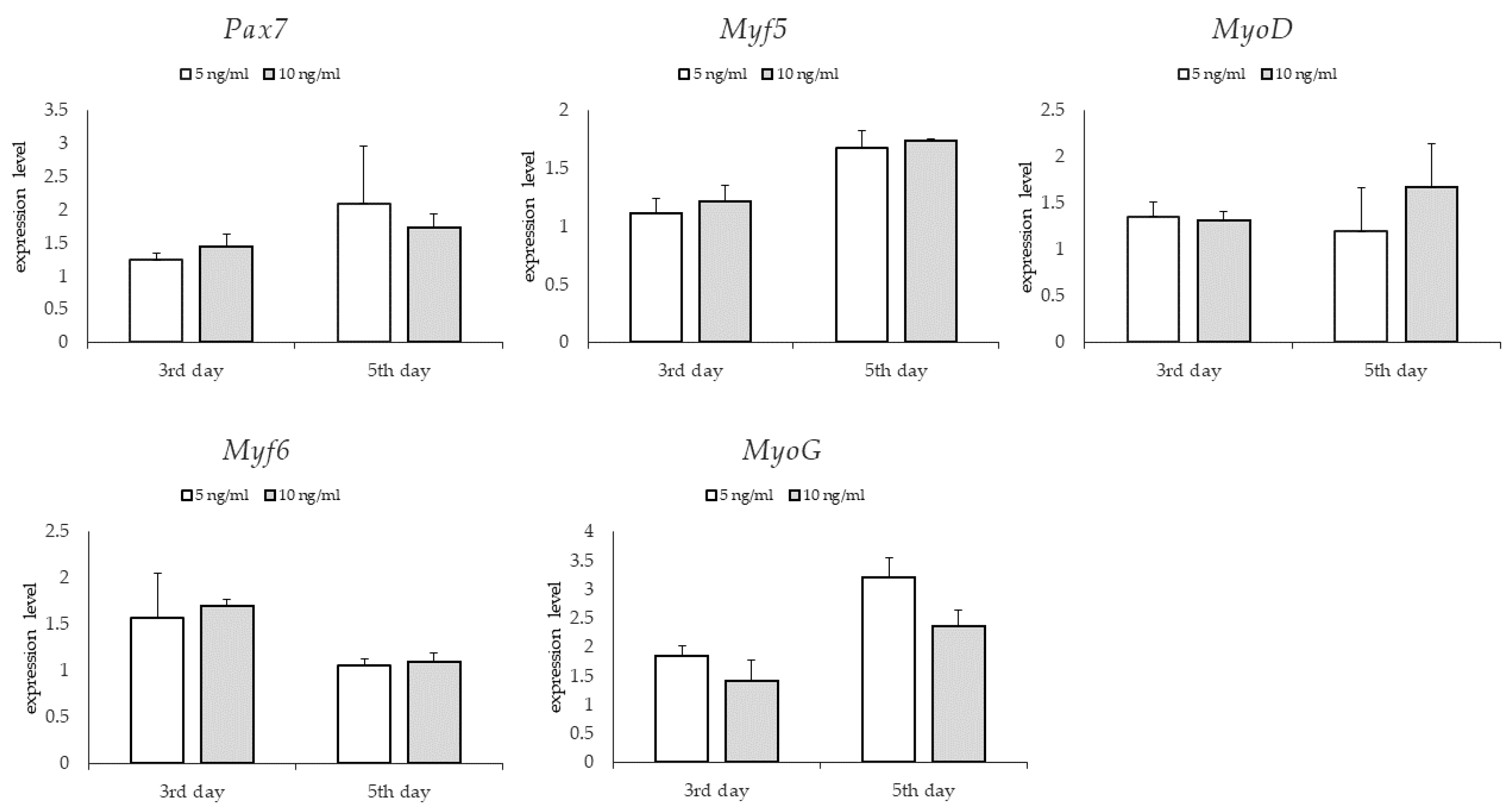
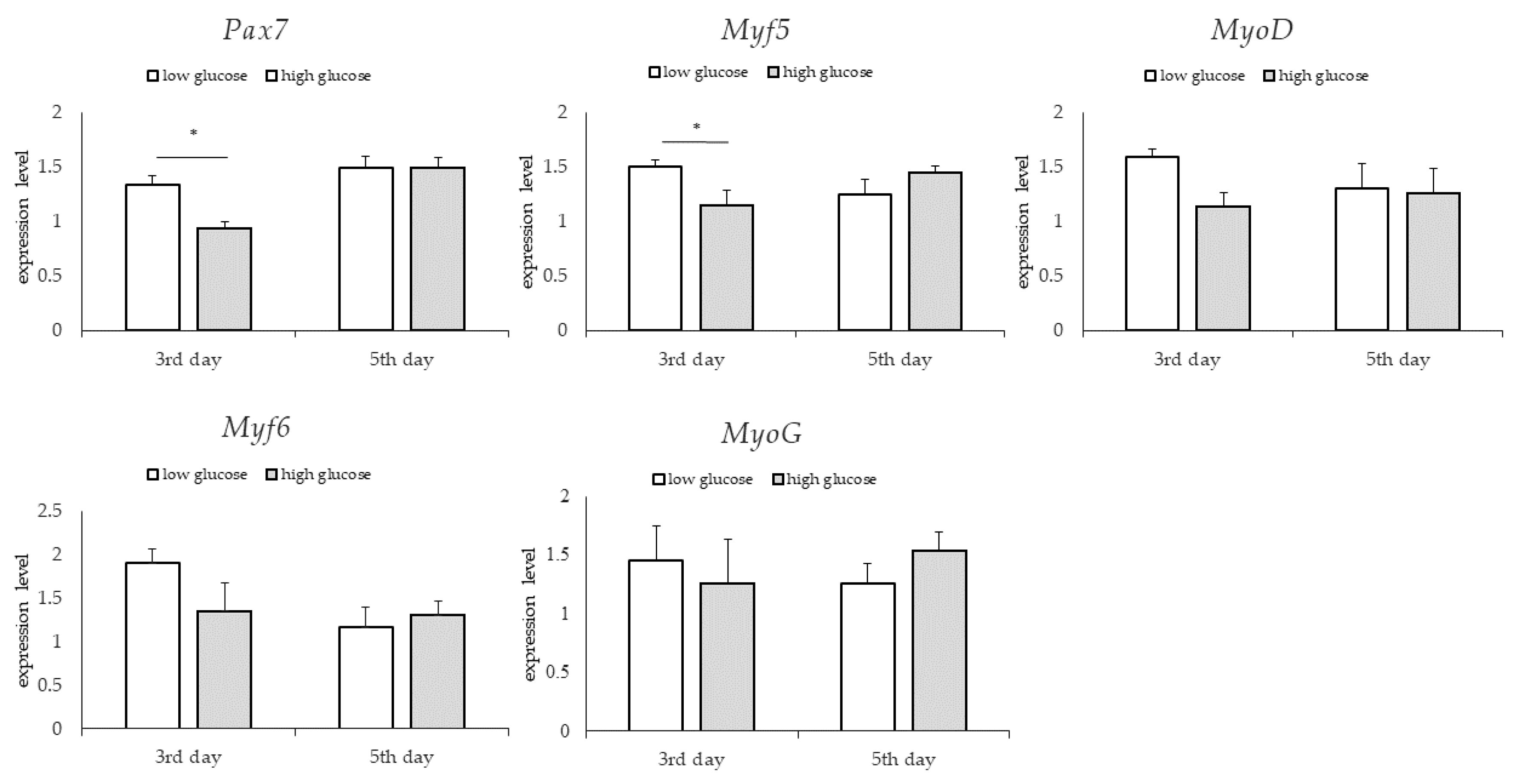
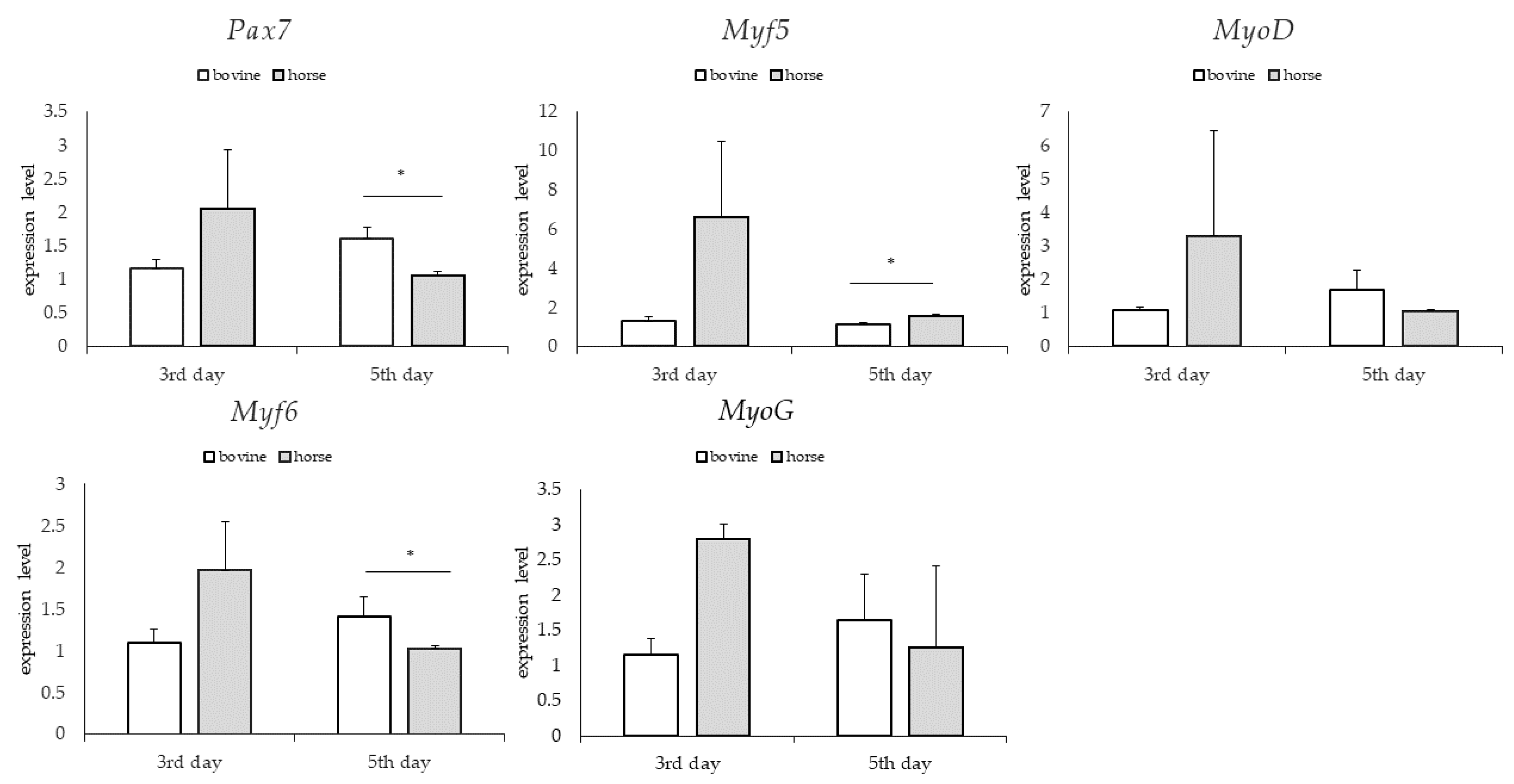
3.3. Gene Expression: Genes Involved in Myogenesis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations. World Population Prospects: The 2019 Revision. Population Database. Available online: https://population.un.org/wpp/ (accessed on 29 May 2023).
- FAO-FAOSTAT. Land Statistics and Indicators. Global, Regional and Country Trends, 2000–2020. Faostat Analytical Brief 48. 2022. Available online: https://www.fao.org/3/cc0963en/cc0963e (accessed on 20 January 2023).
- FAO. Livestock’s Long Shadow. In Environmental Issues and Options; Food and Agriculture Organization of the United Nations: Rome, Italy, 2006. [Google Scholar]
- Risner, D.; Kim, Y.; Nguyen, C.; Siegel, J.B.; Spang, E.S. Environmental impacts of cultured meat: A cradle-to-gate life cycle assessment. bioRxiv 2023, 537778. [Google Scholar] [CrossRef]
- Shapiro, P. Clean meat: How growing meat without animals will revolutionize dinner and the world. Science 2018, 359, 399. [Google Scholar]
- Bhat, Z.F.; Fayaz, H. Prospectus of cultured meat—Advancing meat alternatives. J. Food Sci. Technol. 2011, 48, 125–140. [Google Scholar] [CrossRef]
- Kumar, P.; Sharma, N.; Sharma, S.; Mehta, N.; Verma, A.K.; Chemmalar, S.; Sazili, A.Q. In-vitro meat: A promising solution for sustainability of meat sector. J. Anim. Sci. Technol. 2021, 63, 693–724. [Google Scholar] [CrossRef]
- Mauro, A. Satellite cell of skeletal muscle fibers. J. Biophys. Biochem. Cytol. 1961, 9, 493–495. [Google Scholar] [CrossRef]
- Chargé, S.B.P.; Rudnicki, M.A. Cellular and Molecular Regulation of Muscle Regeneration. Physiol. Rev. 2004, 84, 209–238. [Google Scholar] [CrossRef]
- Chal, J.; Pourquié, O. Making muscle: Skeletal myogenesis in vivo and in vitro. Development 2017, 144, 2104–2122. [Google Scholar] [CrossRef]
- Kuang, S.; Rudnicki, M.A. The emerging biology of satellite cells and their therapeutic potential. Trends Mol. Med. 2008, 14, 82–91. [Google Scholar] [CrossRef]
- Will, K.; Schering, L.; Albrecht, E.; Kalbe, C.; Maak, S. Differentiation of bovine satellite cell-derived myoblasts under different culture conditions. Vitro Cell. Dev. Biol.-Anim. 2015, 51, 885–889. [Google Scholar] [CrossRef]
- Dodson, M.V.; Martin, E.L.; Brannon, M.A.; Mathison, B.A.; McFarland, D.C. Optimization of bovine satellite cell-derived myotube formation in vitro. Tissue Cell. 1987, 19, 159–166. [Google Scholar] [CrossRef]
- Kowalski, K.; dos Santos, M.; Maire, P.; Ciemerych, M.A.; Brzoska, E. Induction of bone marrow-derived cells myogenic identity by their interactions with the satellite cell niche. Stem Cell Res. Ther. 2018, 9, 258. [Google Scholar] [CrossRef]
- Musarò, A.; Barberi, L. Isolation and culture of mouse satellite cells. Methods Mol. Biol. 2010, 633, 101–111. [Google Scholar]
- Zhang, J.; Li, Q.; Yan, Y.; Sun, B.; Wang, Y.; Tang, L.; Wang, E.; Yu, J.; Nogoy, K.M.C.; Li, X.; et al. Effect of ciglitazone on adipogenic transdifferentiation of bovine skeletal muscle satellite cells. J. Anim. Sci. Technol. 2021, 63, 934–953. [Google Scholar] [CrossRef]
- Guthridge, M.; Wilson, M.; Cowling, J.; Bertolini, J.; Hearn, M.T.W. The Role of Basic Fibroblast Growth Factor in Skeletal Muscle Regeneration. Growth Factors 1992, 6, 53–63. [Google Scholar] [CrossRef]
- Yasui, R.; Sekine, K.; Taniguchi, H. Clever Experimental Designs: Shortcuts for Better iPSC Differentiation. Cells 2021, 10, 3540. [Google Scholar] [CrossRef]
- Kolkmann, A.M.; Van Essen, A.; Post, M.J.; Moutsatsou, P. Development of a Chemically Defined Medium for in vitro Expansion of Primary Bovine Satellite Cells. Front. Bioeng. Biotechnol. 2022, 10, 895289. [Google Scholar] [CrossRef]
- Li, J.; Gonzalez, J.M.; Walker, D.K.; Hersom, M.J.; Ealy, A.D.; Johnson, S.E. Evidence of heterogeneity within bovine satellite cells isolated from young and adult animals. J. Anim. Sci. 2011, 89, 1751–1757. [Google Scholar] [CrossRef]
- Lyu, P.; Settlage, R.E.; Jiang, H. Genome-wide identification of enhancers and transcription factors regulating the myogenic differentiation of bovine satellite cells. BMC Genom. 2021, 22, 901. [Google Scholar] [CrossRef]
- Dai, Y.; Wang, Y.M.; Zhang, W.R.; Liu, X.F.; Li, X.; Ding, X.B.; Guo, H. The role of microRNA-1 and microRNA-206 in the proliferation and differentiation of bovine skeletal muscle satellite cells. Vitro Cell. Dev. Biol.-Anim. 2016, 52, 27–34. [Google Scholar] [CrossRef]
- Shahini, A.; Vydiam, K.; Choudhury, D.; Rajabian, N.; Nguyen, T.; Lei, P.; Andreadis, S.T. Efficient and high yield isolation of myoblasts from skeletal muscle. Stem Cell Res. 2018, 30, 122–129. [Google Scholar] [CrossRef]
- Pezzanite, L.; Chow, L.; Griffenhagen, G.; Dow, S.; Goodrich, L. Impact of Three Different Serum Sources on Functional Properties of Equine Mesenchymal Stromal Cells. Front. Vet. Sci. 2021, 8, 634064. [Google Scholar] [CrossRef]
- Franke, J.; Abs, V.; Zizzadoro, C.; Abraham, G. Comparative study of the effects of fetal bovine serum versus horse serum on growth and differentiation of primary equine bronchial fibroblasts. BMC Vet. Res. 2014, 10, 119. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 7, 676–682. [Google Scholar] [CrossRef]
- CLAHE (Contrast Limited Adaptive Histogram Equalization). Available online: https://imagej.nih.gov/ij/plugins/clahe/index.html (accessed on 10 January 2023).
- Van Rossum, G.; Drake, F.L. Python 3 Reference Manual; CreateSpace: Scotts Valley, CA, USA, 2009. [Google Scholar]
- Harris, C.R.; Millman, K.J.; van der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Oliphant, T.E.; et al. Array programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0. Fundamental Algorithms for Scientific Computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef]
- Seabold, S.; Perktold, J. Statsmodels: Econometric and statistical modeling with python. In Proceedings of the 9th Python in Science Conference, Austin, TX, USA, 28 June–3 July 2010; pp. 92–96. [Google Scholar]
- McKinney, W. Data structures for statistical computing in python. In Proceedings of the 9th Python in Science Conference, Austin, TX, USA, 28 June–3 July 2010; Volume 445, pp. 56–61. [Google Scholar]
- The Pandas Development Team. Pandas-Dev/Pandas: Pandas. Zenodo. 2020. Available online: https://zenodo.org/record/7979740 (accessed on 30 January 2023).
- Hunter, J.D. Matplotlib: A 2D Graphics Environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Woods, T.L.; Smith, C.W.; Zeece, M.G.; Jones, S.J. Conditions for the culture of bovine embryonic myogenic cells. Tissue Cell 1997, 29, 207–215. [Google Scholar] [CrossRef]
- Soleimani, V.D.; Punch, V.G.; Kawabe, Y.; Jones, A.E.; Palidwor, G.A.; Porter, C.J.; Cross, J.W.; Carvajal, J.J.; Kockx, C.E.M.; van IJcken, W.F.J.; et al. Transcriptional Dominance of Pax7 in Adult Myogenesis Is Due to High-Affinity Recognition of Homeodomain Motifs. Dev. Cell. 2012, 22, 1208–1220. [Google Scholar] [CrossRef]
- Hinterberger, T.J.; Sassoon, D.A.; Rhodes, S.J.; Konieczny, S.F. Expression of the muscle regulatory factor MRF4 during somite and skeletal myofiber development. Dev. Biol. 1991, 147, 144–156. [Google Scholar] [CrossRef]
- Lazure, F.; Blackburn, D.M.; Corchado, A.H.; Sahinyan, K.; Karam, N.; Sharanek, A.; Nguyen, D.; Lepper, C.; Najafabadi, H.S.; Perkins, T.J.; et al. Myf6/MRF4 is a myogenic niche regulator required for the maintenance of the muscle stem cell pool. EMBO Rep. 2020, 21, 12. [Google Scholar] [CrossRef]
- Pawlikowski, B.; Vogler, T.O.; Gadek, K.; Olwin, B.B. Regulation of skeletal muscle stem cells by fibroblast growth factors. Dev. Dyn. 2017, 246, 359–367. [Google Scholar] [CrossRef]
- Park, J.; Lee, J.; Song, K.D.; Kim, S.J.; Kim, D.C.; Lee, S.C.; Son, Y.J.; Choi, H.W.; Shim, K. Growth factors improve the proliferation of Jeju black pig muscle cells by regulating myogenic differentiation 1 and growth-related genes. Anim. Biosci. 2021, 34, 1392–1402. [Google Scholar] [CrossRef] [PubMed]
- Furuichi, Y.; Kawabata, Y.; Aoki, M.; Mita, Y.; Fujii, N.L.; Manabe, Y. Excess Glucose Impedes the Proliferation of Skeletal Muscle Satellite Cells Under Adherent Culture Conditions. Front. Cell Dev. Biol. 2021, 9, 640399. [Google Scholar] [CrossRef]
- Aguiari, P.; Leo, S.; Zavan, B.; Vindigni, V.; Rimessi, A.; Bianchi, K.; Franzin, C.; Cortivo, R.; Rossato, M.; Vettor, R. High glucose induces adipogenic differentiation of muscle-derived stem cells. Proc. Nat. Acad. Sci. USA 2008, 105, 1226–1231. [Google Scholar] [CrossRef] [PubMed]
- Kolkmann, A.M.; Post, M.J.; Rutjens, M.A.M.; van Essen, A.L.M.; Moutsatsou, P. Serum-free media for the growth of primary bovine myoblasts. Cytotechnology 2020, 72, 111–120. [Google Scholar] [CrossRef]
- Stout, A.J.; Mirliani, A.B.; Rittenberg, M.L.; Shub, M.; White, E.C.; Yuen, J.S.K., Jr.; Kaplan, D.L. Simple and effective serum-free medium for sustained expansion of bovine satellite cells for cell cultured meat. Commun. Biol. 2022, 5, 466. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition of Media for BSCs Cultivation | References |
|---|---|
| Ham’s F-10 Nutrient Mix or DMEM/F-12 as basal medium, supplemented with 20% fbs and 5 ng/mL FGF-2 | [19] |
| Low-glucose DMEM as basal medium, supplemented with 10% hs | [20] |
| DMEM/F12 as basal medium, supplemented with 0.02 M glutamine and 10% fbs | [12,21] |
| DMEM as a basal medium, supplemented with 20% fbs, and 5 ng/mL basic FGF | [22] |
| Variables | Factors | Minimum Value (−1) | Maximum Value (+1) |
|---|---|---|---|
| X1 | Glucose | Low | High |
| X2 | Serum | 20% hs | 20% fbs |
| X3 | bFGF | 5 ng/mL | 10 ng/mL |
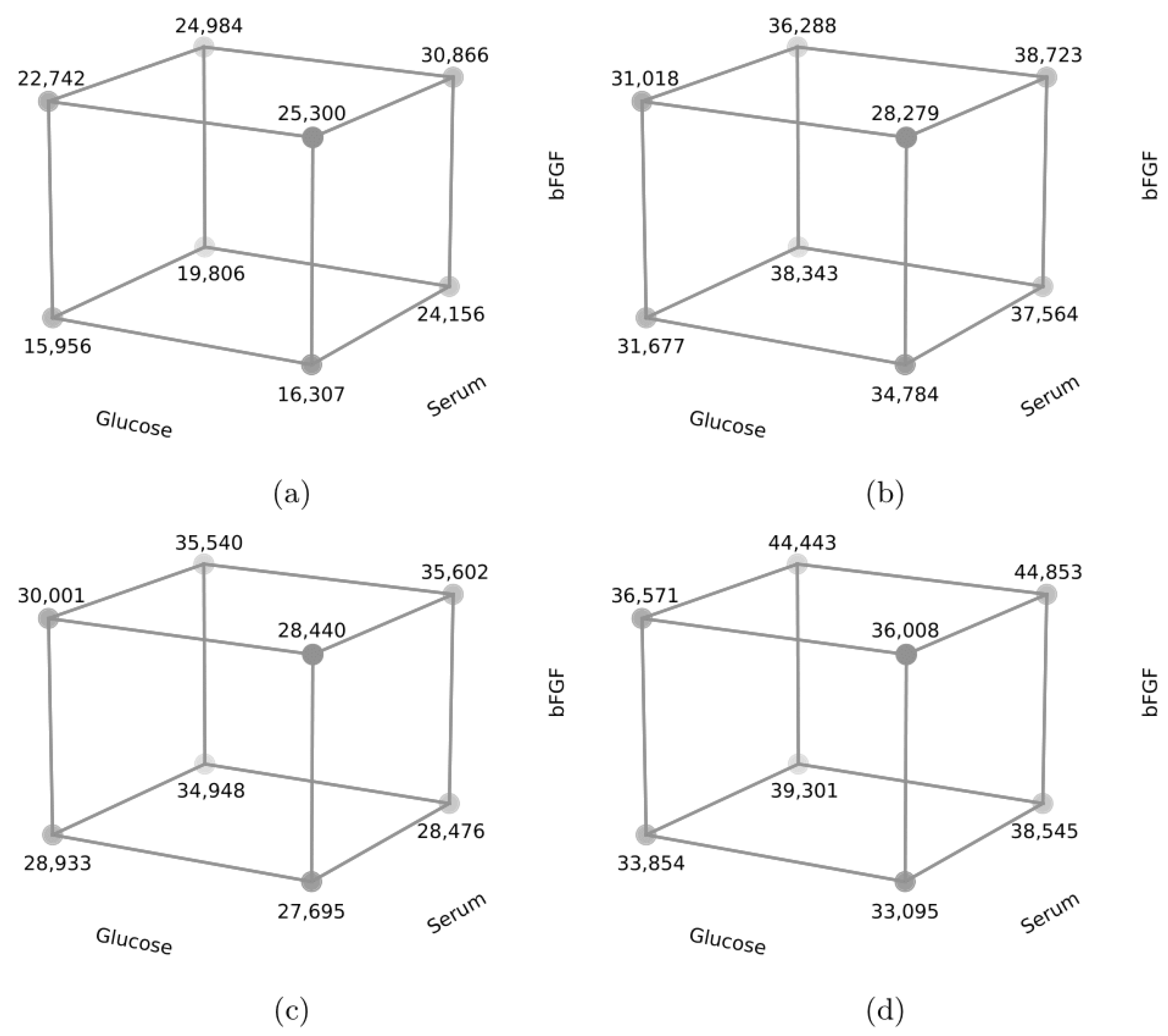
| Glucose | Serum | bFGF | Day | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Experiment | (A) | X1 | (B) | X2 | (C) | X3 | 3 | 5 | 7 | 10 |
| Cell R1 | ||||||||||
| 1 | Low | −1 | 20% hs | −1 | 5 | −1 | 15,955.75 | 31,677.25 | 28,933.00 | 33,853.67 |
| 2 | High | 1 | 20% hs | −1 | 5 | −1 | 16,306.75 | 34,783.50 | 27,695.00 | 33,094.92 |
| 3 | Low | −1 | 20% fbs | 1 | 5 | −1 | 19,805.75 | 38,343.00 | 34,947.75 | 39,300.67 |
| 4 | High | 1 | 20% fbs | 1 | 5 | −1 | 24,156.25 | 37,564.50 | 28,476.25 | 38,544.92 |
| 5 | Low | −1 | 20% hs | −1 | 10 | 1 | 22,741.75 | 31,017.50 | 30,000.75 | 36,570.17 |
| 6 | High | 1 | 20% hs | −1 | 10 | 1 | 25,299.75 | 28,279.25 | 28,439.75 | 36,007.92 |
| 7 | Low | −1 | 20% fbs | 1 | 10 | 1 | 24,983.50 | 36,288.25 | 35,539.75 | 44,442.42 |
| 8 | High | 1 | 20% fbs | 1 | 10 | 1 | 30,865.50 | 38,722.75 | 35,602.00 | 44,852.17 |
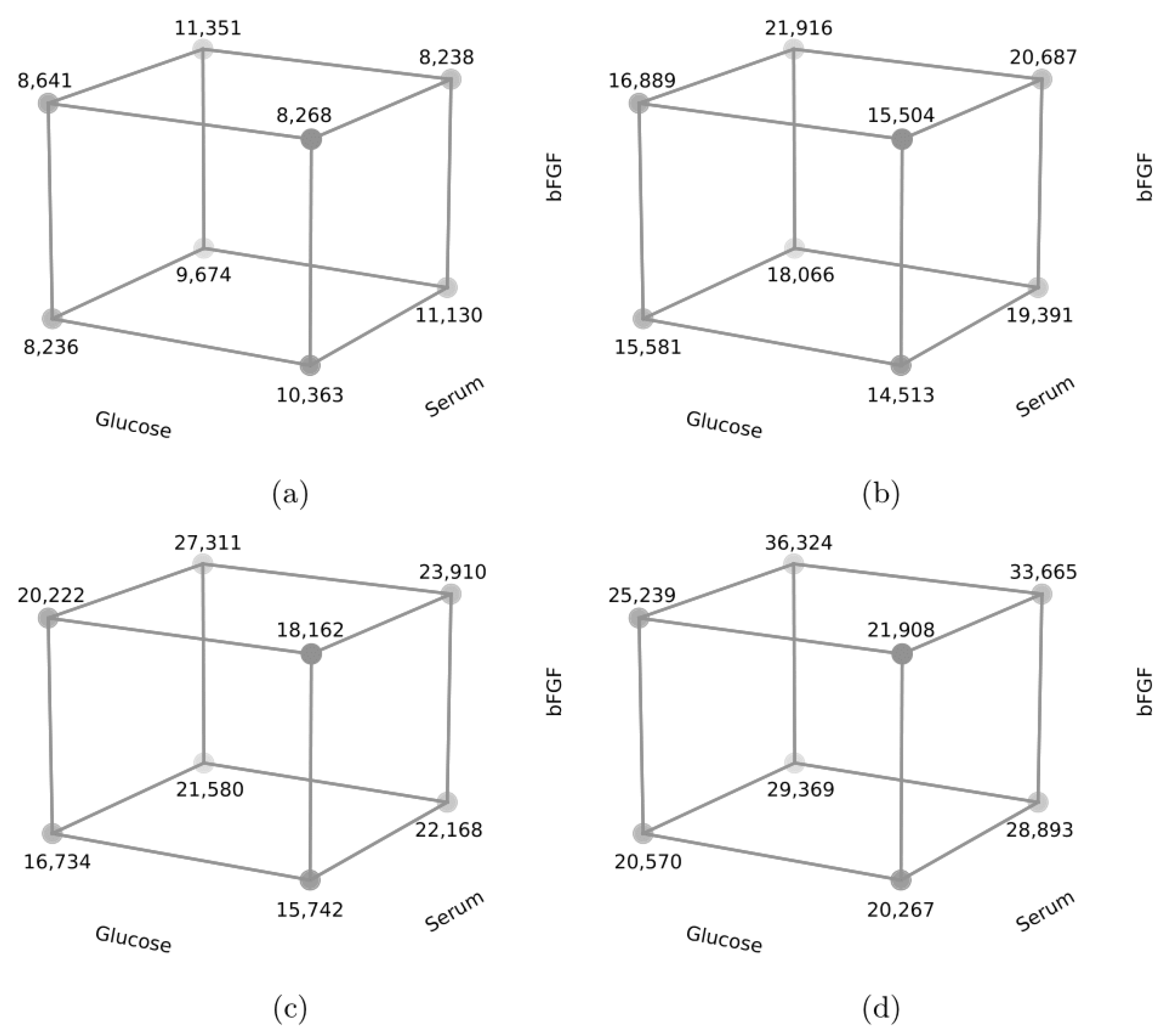
| Cell R2 | ||||||||||
| 1 | Low | −1 | 20% hs | −1 | 5 | −1 | 8235.83 | 15,581.58 | 16,733.53 | 20,570.58 |
| 2 | High | 1 | 20% hs | −1 | 5 | −1 | 10,363.08 | 14,512.83 | 15,741.53 | 20,267.58 |
| 3 | Low | −1 | 20% fbs | 1 | 5 | −1 | 9674.08 | 18,065.83 | 21,579.53 | 29,369.08 |
| 4 | High | 1 | 20% fbs | 1 | 5 | −1 | 11,130.58 | 19,391.58 | 22,167.53 | 28,893.08 |
| 5 | Low | −1 | 20% hs | −1 | 10 | 1 | 8640.83 | 16,889.08 | 20,221.50 | 25,239.33 |
| 6 | High | 1 | 20% hs | −1 | 10 | 1 | 8268.33 | 15,504.33 | 18,161.50 | 21,907.83 |
| 7 | Low | −1 | 20% fbs | 1 | 10 | 1 | 11,351.08 | 21,916.33 | 27,310.75 | 36,324.83 |
| 8 | High | 1 | 20% fbs | 1 | 10 | 1 | 8238.33 | 20,687.58 | 23,910.00 | 33,665.08 |
| Source | Cell Line R1 | Cell Line R2 | ||||||
|---|---|---|---|---|---|---|---|---|
| Day 3 | Day 5 | Day 7 | Day 10 | Day 3 | Day 5 | Day 7 | Day 10 | |
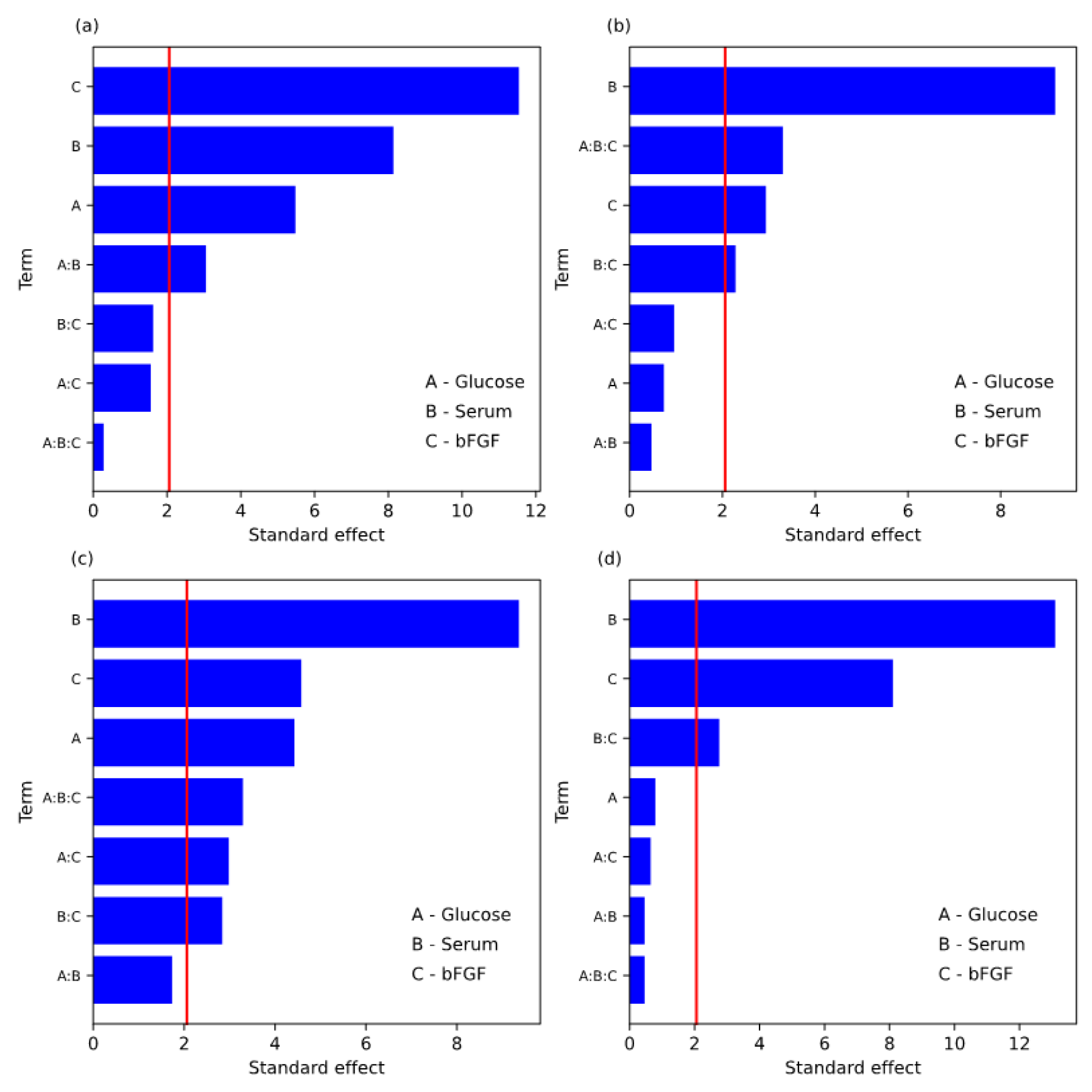
| Main effects | 2.00 × 10−12 | 1.94 × 10−8 | 8.79 × 10−10 | 1.35 × 10−12 | 7.12 × 10−4 | 3.07 × 10−7 | 2.39 × 10−16 | 3.56 × 10−14 |
| 2-way interactions | 9.29 × 10−3 | 1.24 × 10−1 | 1.99 × 10−3 | 6.47 × 10−2 | 8.76 × 10−6 | 5.17 × 10−2 | 3.17 × 10−3 | 4.37 × 10−2 |
| 3-way interaction | 7.81 × 10−1 | 2.99 × 10−3 | 3.06 × 10−3 | 6.50 × 10−1 | 8.69 × 10−2 | 6.58 × 10−4 | 2.72 × 10−2 | 7.34 × 10−1 |
| Gene | Starter Forward | Starter Reverse | Length of the Product |
|---|---|---|---|
| Pax7 | 5′ AAGCGGACAAGAAGGAGGAG 3′ | 5′ CGGGTTCTGACTCCACATCT 3′ | 114 |
| Myf5 | 5′ TGCTTAGGGAACAGGTGGAA 3′ | 5′ AACTGCTGCTCTTTCTGGAC 3′ | 135 |
| MyoD | 5′ AACACTACAGCGGCGACT 3′ | 5′ GTAGTAAGTGCGGTCGTAGC 3′ | 122 |
| Myf6 | 5′ CCCTTCAGCTACAGACCCAA 3′ | 5′ CCTTGGCAGTTATCACGAGC 3′ | 118 |
| MyoG | 5′ TCCAGTACATAGAGCGCCTG 3′ | 5′ CTATGGGAGCTGCATTCACTG 3′ | 121 |
| RPL27 | 5′ ATAATCACCTCATGCCCACAA 3′ | 5′ CATGACCTTTGCCTCTCGTC 3′ | 206 |
| OAZ1 | 5′ TTCGCCAGAGAGAAGGAAGG 3′ | 5′ GGACCCAGGTTACTACAGCA 3′ | 143 |
| Term | Effect | Coefficient | SE Coef | t-Value | p-Value |
|---|---|---|---|---|---|
| Constant | 4.50 × 104 | 22,514.38 | 299.62 | 75.14 | 5.34 × 10−30 |
| A | 3.29 × 103 | 1642.69 | 299.62 | 5.48 | 1.23 × 10−5 |
| B | 4.88 × 103 | 2438.38 | 299.62 | 8.14 | 2.33 × 10−8 |
| C | 6.92 × 103 | 3458.25 | 299.62 | 11.54 | 2.79 × 10−11 |
| A:B | 1.83 × 103 | 915.44 | 299.62 | 3.06 | 5.44 × 10−3 |
| A:C | 9.35 × 102 | 467.31 | 299.62 | 1.56 | 1.32 × 10−1 |
| B:C | −9.73 × 102 | −486.50 | 299.62 | −1.62 | 1.18 × 10−1 |
| A:B:C | −168.875 | −84.44 | 299.62 | −0.28 | 7.81 × 10−1 |
| AIC | BIC | R2 | R2adj | ||
| 573.47 | 585.20 | 0.91 | 0.88 |
| Term | Effect | Coefficient | SE Coef | t-Value | p-Value |
|---|---|---|---|---|---|
| Constant | 6.92 × 104 | 34,584.50 | 342.80 | 100.89 | 4.63 × 10−33 |
| A | 5.06 × 102 | 253.00 | 342.80 | 0.74 | 4.68 × 10−1 |
| B | 6.29 × 103 | 3145.13 | 342.80 | 9.17 | 2.57 × 10−9 |
| C | −2.02 × 103 | −1007.56 | 342.80 | −2.94 | 7.17 × 10−3 |
| A:B | 3.22 × 102 | 161.00 | 342.80 | 0.47 | 6.43 × 10−1 |
| A:C | −6.58 × 102 | −328.94 | 342.80 | −0.96 | 3.47 × 10−1 |
| B:C | 1.57 × 103 | 783.44 | 342.80 | 2.29 | 3.14 × 10−2 |
| A:B:C | 2264.375 | 1132.19 | 342.80 | 3.30 | 2.99 × 10−3 |
| AIC | BIC | R2 | R2adj | ||
| 582.09 | 593.81 | 0.82 | 0.77 |
| Term | Effect | Coefficient | SE Coef | t-Value | p-Value |
|---|---|---|---|---|---|
| Constant | 6.24 × 104 | 31,204.28 | 260.24 | 119.90 | 7.40 × 10−35 |
| A | −2.30 × 103 | −1151.03 | 260.24 | −4.42 | 1.80 × 10−4 |
| B | 4.87 × 103 | 2437.16 | 260.24 | 9.36 | 1.74 × 10−9 |
| C | 2.38 × 103 | 1191.28 | 260.24 | 4.58 | 1.22 × 10−4 |
| A:B | −9.03 × 102 | −451.28 | 260.24 | −1.73 | 9.57 × 10−2 |
| A:C | 1.55 × 103 | 776.34 | 260.24 | 2.98 | 6.46 × 10−3 |
| B:C | 1.48 × 103 | 738.16 | 260.24 | 2.84 | 9.12 × 10−3 |
| A:B:C | 1714.188 | 857.09 | 260.24 | 3.29 | 3.06 × 10−3 |
| AIC | BIC | R2 | R2adj | ||
| 564.45 | 576.18 | 0.87 | 0.83 |
| Term | Effect | Coefficient | SE Coef | t-Value | p-Value |
|---|---|---|---|---|---|
| Constant | 7.67 × 104 | 38,333.69 | 263.46 | 145.50 | 7.17 × 10−37 |
| A | −4.17 × 102 | −208.38 | 263.46 | −0.79 | 4.37 × 10−1 |
| B | 6.90 × 103 | 3451.69 | 263.46 | 13.10 | 1.99 × 10−12 |
| C | 4.27 × 103 | 2134.81 | 263.46 | 8.10 | 2.51 × 10−8 |
| A:B | 2.44 × 102 | 121.88 | 263.46 | 0.46 | 6.48 × 10−1 |
| A:C | 3.41 × 102 | 170.25 | 263.46 | 0.65 | 5.24 × 10−1 |
| B:C | 1.45 × 103 | 727.44 | 263.46 | 2.76 | 1.09 × 10−2 |
| A:B:C | 242.25 | 121.13 | 263.46 | 0.46 | 6.50 × 10−1 |
| AIC | BIC | R2 | R2adj | ||
| 565.24 | 576.97 | 0.91 | 0.89 |
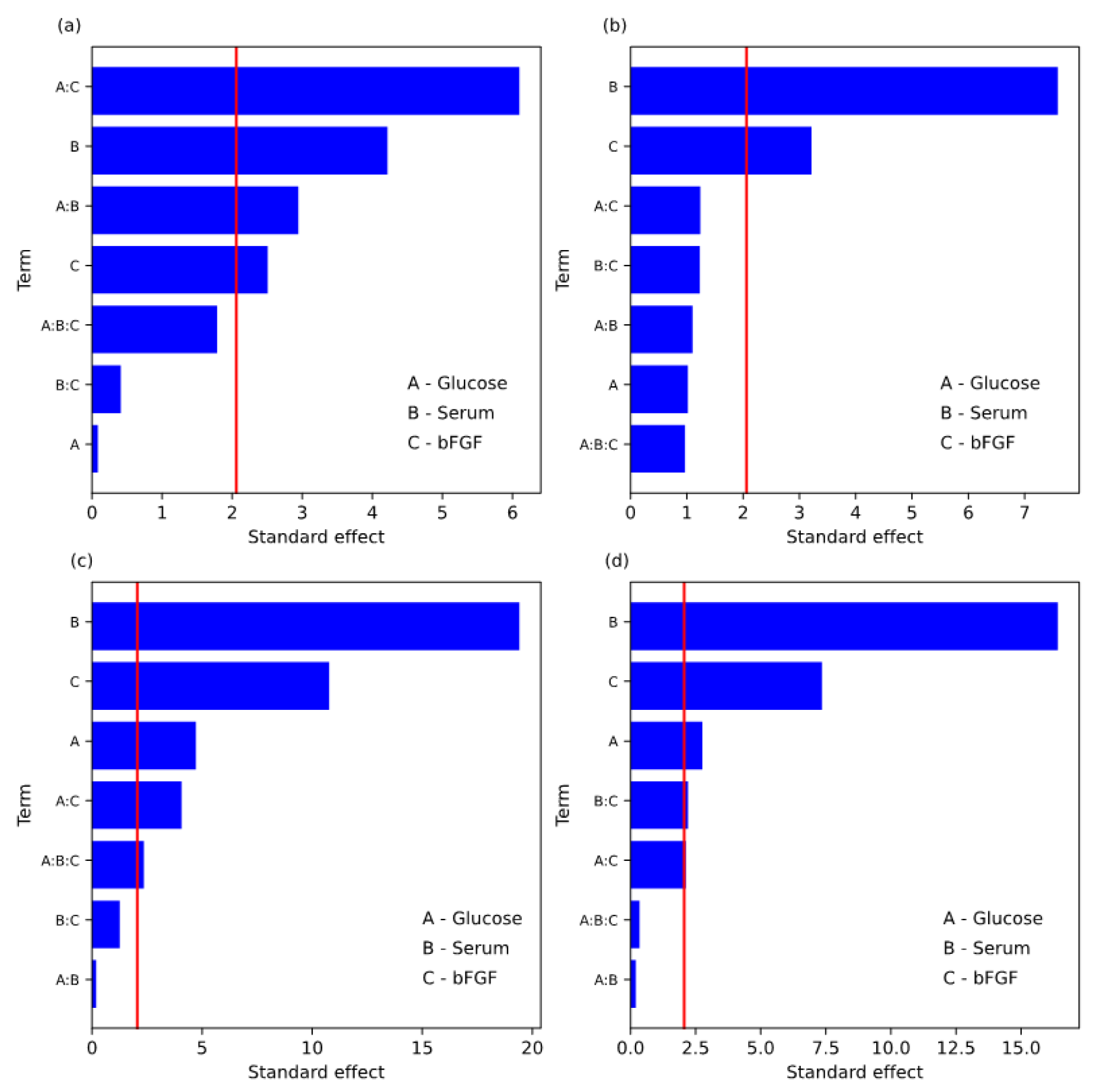
| Term | Effect | Coefficient | SE Coef | t-Value | p-Value |
|---|---|---|---|---|---|
| Constant | 1.90 × 104 | 9487.44 | 144.90 | 65.48 | 1.43 × 10−28 |
| A | 2.46 × 101 | 12.31 | 144.90 | 0.08 | 9.33 × 10−1 |
| B | 1.22 × 103 | 610.75 | 144.90 | 4.22 | 3.06 × 10−4 |
| C | −7.26 × 102 | −363.13 | 144.90 | −2.51 | 1.94 × 10−2 |
| A:B | −8.53 × 102 | −426.38 | 144.90 | −2.94 | 7.11 × 10−3 |
| A:C | −1.77 × 103 | −883.63 | 144.90 | −6.10 | 2.68 × 10−6 |
| B:C | 1.19 × 102 | 59.31 | 144.90 | 0.41 | 6.86 × 10−1 |
| A:B:C | −517.375 | −258.69 | 144.90 | −1.79 | 8.69 × 10−2 |
| AIC | BIC | R2 | R2adj | ||
| 526.98 | 538.70 | 0.75 | 0.68 |
| Term | Effect | Coefficient | SE Coef | t-Value | p-Value |
|---|---|---|---|---|---|
| Constant | 3.56 × 104 | 17,818.31 | 289.42 | 61.57 | 6.22 × 10−28 |
| A | −5.89 × 102 | −294.56 | 289.42 | −1.02 | 3.19 × 10−1 |
| B | 4.39 × 103 | 2196.69 | 289.42 | 7.59 | 7.91 × 10−8 |
| C | 1.86 × 103 | 930.69 | 289.42 | 3.22 | 3.70 × 10−3 |
| A:B | 6.38 × 102 | 318.81 | 289.42 | 1.10 | 2.82 × 10−1 |
| A:C | −7.18 × 102 | −358.81 | 289.42 | −1.24 | 2.27 × 10−1 |
| B:C | 7.12 × 102 | 355.94 | 289.42 | 1.23 | 2.31 × 10−1 |
| A:B:C | −559.625 | −279.81 | 289.42 | −0.97 | 3.43 × 10−1 |
| AIC | BIC | R2 | R2adj | ||
| 571.25 | 582.98 | 0.76 | 0.68 |
| Term | Effect | Coefficient | SE Coef | t-Value | p-Value |
|---|---|---|---|---|---|
| Constant | 4.15 × 104 | 20,728.22 | 155.25 | 133.51 | 5.63 × 10−36 |
| A | −1.47 × 103 | −733.09 | 155.25 | −4.72 | 8.42 × 10−5 |
| B | 6.03 × 103 | 3013.72 | 155.25 | 19.41 | 3.52 × 10−16 |
| C | 3.35 × 103 | 1672.72 | 155.25 | 10.77 | 1.12 × 10−10 |
| A:B | 5.98 × 101 | 29.91 | 155.25 | 0.19 | 8.49 × 10−1 |
| A:C | −1.26 × 103 | −632.09 | 155.25 | −4.07 | 4.40 × 10−4 |
| B:C | 3.91 × 102 | 195.72 | 155.25 | 1.26 | 2.20 × 10−1 |
| A:B:C | −730.1875 | −365.09 | 155.25 | −2.35 | 2.72 × 10−2 |
| AIC | BIC | R2 | R2adj | ||
| 531.39 | 543.12 | 0.96 | 0.94 |
| Term | Effect | Coefficient | SE Coef | t-Value | p-Value |
|---|---|---|---|---|---|
| Constant | 5.41 × 104 | 27,029.34 | 306.69 | 88.13 | 1.18 × 10−31 |
| A | −1.69 × 103 | −846.28 | 306.69 | −2.76 | 1.09 × 10−2 |
| B | 1.01 × 104 | 5033.34 | 306.69 | 16.41 | 1.51 × 10−14 |
| C | 4.51 × 103 | 2254.59 | 306.69 | 7.35 | 1.36 × 10−7 |
| A:B | 1.25 × 102 | 62.34 | 306.69 | 0.20 | 8.41 × 10−1 |
| A:C | −1.30 × 103 | −651.53 | 306.69 | −2.12 | 4.41 × 10−2 |
| B:C | 1.35 × 103 | 677.34 | 306.69 | 2.21 | 3.70 × 10−2 |
| A:B:C | 211.1875 | 105.59 | 306.69 | 0.34 | 7.34 × 10−1 |
| AIC | BIC | R2 | R2adj | ||
| 574.96 | 586.69 | 0.93 | 0.91 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zygmunt, K.; Otwinowska-Mindur, A.; Piórkowska, K.; Witarski, W. Influence of Media Composition on the Level of Bovine Satellite Cell Proliferation. Animals 2023, 13, 1855. https://doi.org/10.3390/ani13111855
Zygmunt K, Otwinowska-Mindur A, Piórkowska K, Witarski W. Influence of Media Composition on the Level of Bovine Satellite Cell Proliferation. Animals. 2023; 13(11):1855. https://doi.org/10.3390/ani13111855
Chicago/Turabian StyleZygmunt, Karolina, Agnieszka Otwinowska-Mindur, Katarzyna Piórkowska, and Wojciech Witarski. 2023. "Influence of Media Composition on the Level of Bovine Satellite Cell Proliferation" Animals 13, no. 11: 1855. https://doi.org/10.3390/ani13111855
APA StyleZygmunt, K., Otwinowska-Mindur, A., Piórkowska, K., & Witarski, W. (2023). Influence of Media Composition on the Level of Bovine Satellite Cell Proliferation. Animals, 13(11), 1855. https://doi.org/10.3390/ani13111855