1. Introduction
Human consumption of fish as an animal protein source has increased over the last decades because of the high nutrient quality of the fish. This consumption has to be covered by the aquaculture species since the capture fisheries are not able to meet the consumer demands and their quota in most countries has been stabilized over the last years [
1]. A major concern for most aquaculture farms is that nutrition and the associated cost of the use of fish meal and fish oil is increasing; it is considered a necessity in an aquafeed. The availability of the last two raw materials is sometimes limited and their prices fluctuate, largely because of the increase in aquaculture production [
2]. In order to reduce the cost of the nutrition, many efforts have been made to use alternative raw materials as a source of protein, such as plant-based proteins [
1,
2,
3,
4,
5], some of these studies showed that a part of fishmeal and fish oil could be replaced in a fish diet [
6]. Apart from the nutrition issues, the breeding programs that are running, sometimes on a large scale in aquaculture farming, have a target to improve the genetic gain for specific phenotypes per generation [
7,
8,
9,
10]. The main selection trait is body weight [
8] and the average genetic gain for body weight that is observed is a 12.7% improvement per generation with a range of 2.3% to 42%, based on Gjedrem and Rye [
9], who summarized the information from most of the studied species. In gilthead seabream, 7.3% and 22% is the expected genetic gain per generation for the body weight [
9,
11,
12]. However, some phenotypes, especially for the quantitative traits such as body weight and growth, showed changes in performance in different environments, which indicates the existence of genotype interaction with the environment. In simple terms, not only is the phenotype affected by the environmental factors but also the performance of different genotypes changes. As a result, the ranking of the selection candidates in the genetic evaluation is fluctuating [
7,
10]. Thus, two major concerns for the partial or total replacement of the fishmeal/fish oil in diets for fish farming species for a breeding strategy are the potential re-ranking of the fish, which affects the selected candidates per generation, along with the query of the establishment of a fish line selection used for plant-based diet [
13]. In some species, the replacement of the fishmeal/fish oil has been studied at the genetic level and the genotype by diet interaction has been investigated (
Oncorhynchus mykiss,
Coregonus lavaretus,
Ictalurus punctatus and
Dicentrarchus labrax, [
13,
14,
15,
16,
17,
18,
19,
20,
21]. Some of those studied showed the presence of G X D [
13,
17,
18,
19,
20,
21], which affected the re-ranking of the families [
13,
17,
18]. However, the establishment of a fish line selection used for a plant-based diet might not be needed (i.e., prediction of the genetic gain [
20]). Focusing on the gilthead seabream, even though the replacement of the part or the total of fishmeal has been investigated [
1,
2,
3,
4,
5,
6], the existence of genotype by diet interaction has not been studied yet for the body weight and fat. So, the two main concerns of the re-ranking of the selection candidates within different selection lines per diet remain unanswered.
The aim of the study is first to investigate behavior of the estimated genetic parameters for the body weight, growth and fat under different protein source diets to identify any possible genotype × diet interaction using a plant-based diet in comparison with a standard commercial diet (standard diet using fishmeal/fish oil), to study any potential effects of possible genotype by diet interaction on the genetic evaluation and the selection decision on the broodstock candidates and finally to investigate the expected genetic gain on a plant-protein-based diet and on a fishmeal/fish-oil-based diet (standard diet).
4. Discussion
In the present study, using a substantial population of 8356 gilthead seabream selection candidates from a commercial breeding program and originating from two different reproductive years, full-sibs from each family were randomly divided into two sea cages each year and were fed on different diets: a high-plant-protein diet containing 85% plant proteins and a high-animal-protein diet containing 30% marine animal protein. These fish were used to estimate the genetic parameters for the body weight performance and fat content under different protein source diets, in order to: (a) investigate any possible genotype by diet interactions, (b) to study the effects of genotype by diet interaction on the genetic evaluation and (c) to investigate the expected genetic gain on a plant-protein-based diet and on a fishmeal/fish-oil-based diet (standard diet).
The heritability estimates using the multivariate animal model in the literature for gilthead seabream body weight at 130, 165 and 509 DPH were 0.28, 0.32 and 0.34, respectively [
25]. At an older age, at 689–690 DPH the heritability estimates of the body weight were 0.25 or 0.29 [
26,
27]; similarly, at 980 DPH, the heritability estimate was 0.24 [
28]. However, Gulzar et al. [
29], estimated the heritability of the harvest weight (approximately 400 g) equal to 0.37 in a Greek farm (Galaxidi, Greece) and 0.55 in a farm in Spain. In our study, the heritability of the tagging weight at 130–169 DPH, was higher than the estimation of Navaro et al. [
25]. However, our heritability decreased at each stage and in the final body weight (549–644 DPH) looks similar (0.39) to the heritability that is described by the aforementioned range and closer to the estimated heritability in the Greek farm in Galaxidi. When the analysis was performed for each batch separately, the heritability estimates for the body weight were slightly different between batches, higher heritability was estimated in batch 16 and lower in batch 17 (
Table 5). The growth also followed same pattern: higher heritability in batch 16 than 17. As far as the fat content is concerned, different heritability estimates were reported for the heritability of the muscular fat (0.31, García-Celdrán et al.) [
26] and for the fillet fat in a Greek site (0.46, Galaxidi) [
29] and in a Spanish site (0.55, Cudomar) [
29]. In our study the heritability estimate of fat content (0.41) was closer to the heritability estimates for fillet fat, which was estimated at the Galaxidi site.
Furthermore, several studies investigating the genotype × diet interaction have been made in other species, such as rainbow trout, European seabass, etc., using plant diets, revealing in general the presence of genotype × diet interaction using a plant protein diet [
13,
17,
19,
20,
30]. However, no information has been found in the literature studying the genotype × diet interaction in gilthead seabream using a plant-based diet. In rainbow trout, Le Boucher et al. (2011) [
19] studied the presence of genotype by diet interaction and reported significant interaction of sire and diet and of dam and diet for body weight at 343–465 days post fertilization (DPF). A significant family × diet interaction was detected in the body weight at the age of 18 weeks, whereas the same interaction was not significant for growth rate in rainbow trout [
14]. In European sea bass, Le Boucher et al. [
13] identified a significant sire x diet interaction. In our study, a significant interaction between dam and diet, as it was reported by Le Boucher et al. [
19], and not between sire and diet was detected in body weight performance, as was reported by Le Boucher et al. [
13,
19] in European sea bass and in rainbow trout. Studying the fat content, a significant interaction was detected between sire and diet as it was detected in the last measurements of the fillet fat by Le Boucher et al. [
13].
In rainbow trout, Le Boucher et al. [
19] showed high genetic correlations (0.67–0.90) for body weight at different growth stages (213–464 DPF) indicating a G × D between the last three measurements of the body weight. In European sea bass, Le Boucher et al. [
13] reported that the genetic correlation for the body weight was 0.51 to 0.81 (679–850 DPF), and for fillet lipid content was from 0.80 to 0.87 (736–787 rDPF). Also, Le Boucher et al. [
20] also investigating the G × D, found that the genetic correlation of the body weight between diets ranged from 0.7 to 0.8 in European seabass. Focusing on the genetic correlations from the bivariate animal models in our study, they were high (0.90–0.99,
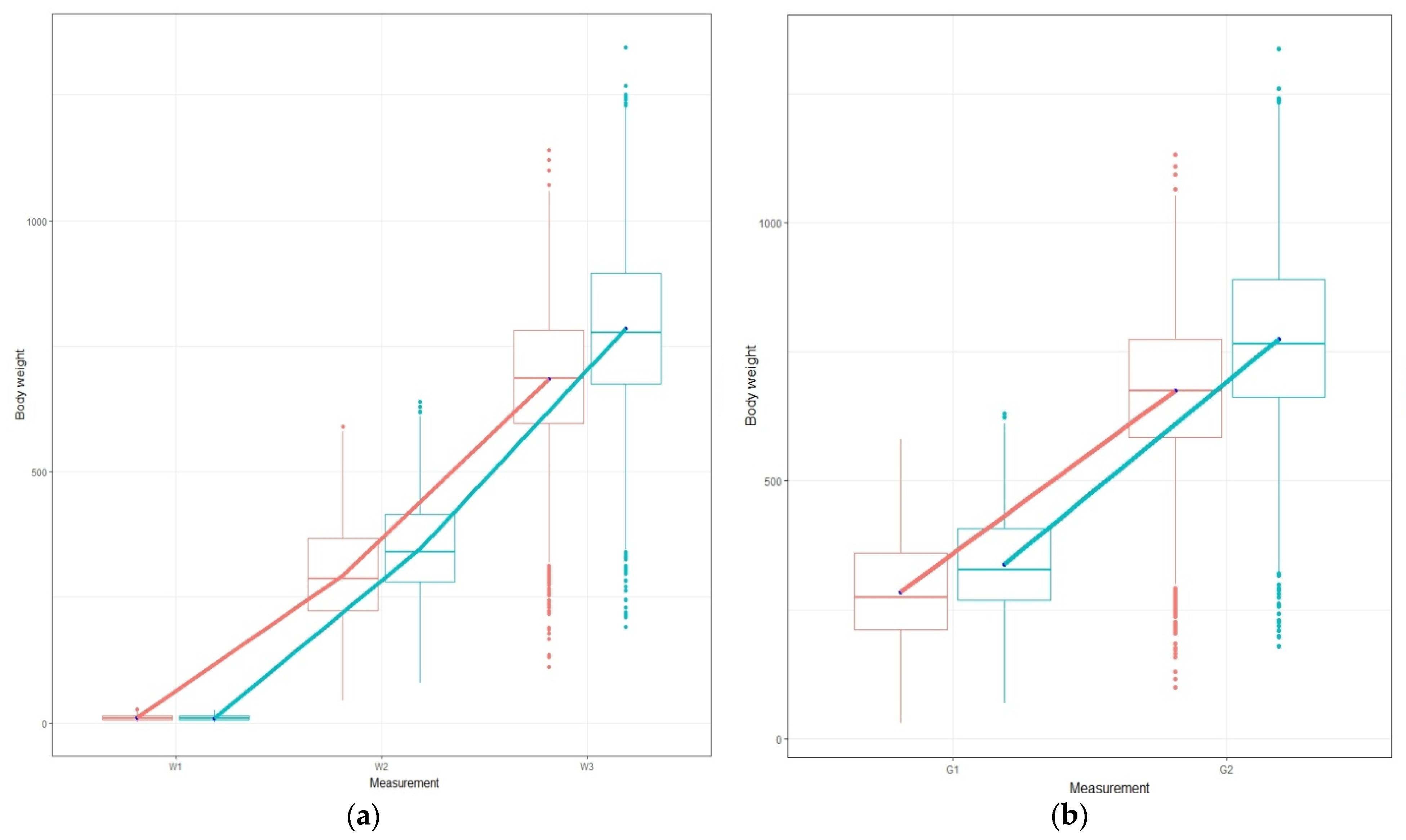
Table 5) for each studied trait, illustrating that selecting candidates based on the standard commercial diet could lead to parallel genetic responses (
Figure 1) in the plant diet [
7]. Generally, when the genetic correlation between two traits is close to one, a possible pleiotropy and/or linkage among the genes affecting these traits is indicated [
10] and consequently they may infer that the phenotypes are under the same genetic control in each environment (i.e., diet in our case). A threshold value for the genetic correlation, which can be used to consider whether an effect of G × E on each of the traits is significant or not, was suggested to be 0.8 [
7,
31]. For example, according to Robertson [
31] a threshold value for genetic correlation of 0.8 between the same trait measurements under two different environments determines whether a different selection lines strategy (one for each environment) should be used in a breeding program. Based on that criterion, the effect of the G × E is considered “strong” and a two different lines strategy is required. Although such a threshold value could not indicate the statistical significance of the G × E, it is generally the “rule of thumb” in an applied breeding program. If the genetic correlation values are close to 0.8, it is necessary that a two breeding lines strategy is utilized. However, in such a scenario it is mandatory to perform a sib test with different diets, which should have a cost/benefit analysis. Such sib testing procedures have been used in the poultry breeding industry (D. Chatziplis, personal communications).
Nevertheless, Le Boucher et al. [
13], when studying the European seabass, identified a strong G × D in the last measurements of the body weight; however, in this case a total replacement of the fish meal was used, whereas in our study, only a part of the fish meal was replaced. Possibly, the percentage of the replacement of the fishmeal could affect the presence of the G × D [
18]. For the gilthead seabream, a significant G × D interaction has been found when studying the specific growth rate (growth divided per day) but with a weak effect using a linear mixed model and the phenotypic correlation [
32]. In our study, however, we used the body weight at three different times and growth, as a difference between them, and the fat. Even though the effect of G × D is not very strong compared with the aforementioned threshold (0.8, [
7,
31]), the re-ranking of the selection candidates in a genetic evaluation is quite high. Palti et al. [
15] reported that the ranking of the families between the diets was similar in rainbow trout, whereas in European sea bass, Le Boucher et al. [
13] identified a significant re-ranking, following the genetic evaluation, of the sire families between diets for the body weight (605–850 DPF) and fillet lipid content (736–850 DPF). We notice a significant re-ranking of the fish using the bivariate animal model for the body weight, growth and fat content as was mentioned earlier. The re-ranking is further enhanced in the selected brooders after applying the same selection intensity in the selection candidates. However, the re-ranking of the selection candidates is quite extensive (0.18–0.33), which means that only 18–33% of the fish were in the same position independently of diet. This proportion could possibly justify the design of a breeding program based on the diet (i.e., a selection line for a plant-based diet and a selection line for the standard commercial diet) but it could not offer significant improvement of the breeding goal in gilthead seabream in the aquaculture farm. This can be observed since a higher genetic gain can be achieved by selecting while using the standard commercial diet instead of a plant diet in the terms of the body weight performance (based on the estimated genetic gain which was achieved per diet in the present study (
Table 7)). The aforementioned findings agreed with Le Boucher et al. [
20], in which higher genetic gain can be achieved for body weight using the control diet for 5% of selection intensity. However, a different pattern was observed in relation to the selection for fat content (%), where a higher genetic gain is expected using the plant diet; however, it should be noted that the average values for fat content were lower in the plant-based diet than the control. According to Sae-Lim et al. [
7], a higher genetic gain can be achieved using high selection intensity when G × E exists, and indeed, in our study a high genetic gain was detected with selection intensities (1.26%); however, the ranking correlation coefficient between the plant and standard commercial diet was extremely low, which indicates a strong re-ranking of the fish (
Table 7). Nevertheless, the establishment of a plant-based diet breeding strategy could be possible, if the reduction in the cost of aquafeed could balance the lower genetic gain that is expected to be achieved in each generation. However, it should be noted that this has to be re-evaluated when more selection objectives and more selection criteria are utilized in a breeding program. Alternatively, the cost effectiveness of a sib testing procedure with the alternative diet (e.g., plant-based diet) should be examined and reviewed before its incorporation.
Focusing on the selected methodology of the present study for the investigation of the genotype by diet interaction, firstly a linear model fitting the interaction effect Equation (2) was selected in order to identify the presence of G × D and more specifically the interaction between parents and diet, and secondly the bivariate animal model (using as trait 1 the phenotypes of the plant-based diet and as trait 2 the phenotypes of the standard commercial diet for each trait measured) was used in order to identify the effects of G × D in detail to estimate the existence of heterogeneity of variance and to investigate the re-ranking of the selection candidates for the studied traits [
7]. According to Sae-Lim et al. [
7], for a trait with moderate heritability approximately 10 fish per family and 200 families are needed to identify the G × E using the multitrait animal model. In our study, based on the heritability of the studied traits (which ranged between 0.39–0.42), there is a satisfactory representation of each family since the range of offspring was 12 to 48 offspring (apart from one family which included only one offspring) and 216 families were included in the present analysis. The optimal sample size is 2000 fish for a trait with moderate heritability; in our study, 8356 gilthead seabream fish from a commercial breeding program were used.
Even though two-year batches were included in the present analysis, no selection of candidates based only on the plant diet was performed in order to structure the families and to evaluate the theoretical genetic gain (
Table 8) using two generations and a selection line for plant diet and standard commercial diet. Selecting candidates based on the plant diet could provide a validation of the overall estimations in the present analysis, since the present estimation of the genetic gain appears to be biased because fish were selected from many generations using the standard commercial diet.

 ,
, 

{kind=link}