
Screening and Carrier Rate of Neuronal Ceroid Lipofuscinosis in Chihuahua Dogs in Japan


 , ,
, ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Storage
2.2. Genotyping of the c.846delT Mutation in the Canine CLN7/MFSD8 Gene
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chalkley, M.D.; Armien, A.G.; Gilliam, D.H.; Johnson, G.S.; Zeng, R.; Wünschmann, A.; Kovi, R.C.; Katz, M.L. Characterization of neuronal ceroid lipofuscinosis in 3 cats. Vet. Pathol. 2014, 51, 796–804. [Google Scholar] [CrossRef]
- Jolly, R.D.; Palmer, D.N. The neuronal ceroid lipofuscinoses (Batten disease): Comparative aspects. Neuropathol. Appl. Neurobiol. 1995, 21, 50–60. [Google Scholar] [CrossRef]
- Jolly, R.D.; Palmer, D.N.; Studdert, V.P.; Sutton, R.H.; Kelly, W.R.; Koppang, N.; Dahme, G.; Hartley, W.J.; Patterson, J.S.; Riis, R.C. Canine ceroid lipofuscinosis: A review and classification. J. Small Anim. Pract. 1994, 35, 299–306. [Google Scholar] [CrossRef]
- Katz, M.L.; Rustad, E.; Robinson, G.O.; Whiting, R.E.H.; Student, J.T.; Coates, J.R.; Narfstrom, K. Canine neuronal ceroid lipofuscinoses: Promising models for preclinical testing of therapeutic interventions. Neurobiol. Dis. 2017, 108, 277–287. [Google Scholar] [CrossRef]
- NCL Resource—A Gateway for Batten Disease. Available online: https://www.ucl.ac.uk/ncl/ (accessed on 27 April 2022).
- Katz, M.L.; Khan, S.; Awano, T.; Shahid, S.A.; Sialotos, A.N.; Siakotos, G.S.; Johnson, G.S. A mutation in the CLN8 gene in English Setter dogs with neuronal ceroid-lipofuscinosis. Biochem. Biophys. Res. Commun. 2005, 327, 541–547. [Google Scholar] [CrossRef]
- Asakawa, M.G.; MacKillop, E.; Olby, N.J.; Robertson, I.D.; Cullen, J.M. Imaging diagnosis—Neuronal ceroid lipofuscinosis with a chronic subdural hematoma. Vet. Radiol. Ultrasound 2010, 51, 155–158. [Google Scholar] [CrossRef] [PubMed]
- Sanders, D.N.; Farias, F.H.; Johnson, G.S.; Chiang, V.; Cook, J.R.; O’Brien, D.P.; Hofmann, S.L.; Lu, J.Y.; Katz, M.L. A mutation in canine PPT1 causes early onset neuronal ceroid lipofuscinosis in a Dachshund. Mol. Genet. Metab. 2010, 100, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; O’Brien, D.P.; Mhlanga-Mutangadura, T.; Olby, N.J.; Taylor, J.F.; Schnabel, R.D.; Katz, M.L.; Johnson, G.S. A rare homozygous MFSD8 single-base-pair deletion and frameshift in the whole genome sequence of a Chinese Crested Dog with neuronal ceroid lipofuscinosis. BMC Vet. Res. 2015, 10, 960. [Google Scholar] [CrossRef] [Green Version]
- Lingaas, F.; Guttersrud, O.A.; Arnet, E.; Espenes, A. Neuronal ceroid lipofuscinosis in salukis is caused by a single pair insertion in CLN8. Anim. Genet. 2018, 49, 52–58. [Google Scholar] [CrossRef]
- Rossmeisl, J.H., Jr.; Duncan, R.; Fox, J.; Herring, E.S.; Inzana, K.D. Neuronal ceroid-lipofuscinosis in a Labrador Retriever. J. Vet. Diagn. Investig. 2003, 15, 457–460. [Google Scholar] [CrossRef] [Green Version]
- Koie, H.; Shibuya, H.; Sato, T.; Sato, A.; Nawa, K.; Nawa, Y.; Kitagawa, M.; Sakai, M.; Takahashi, T.; Yamaya, Y.; et al. Magnetic resonance imaging of neuronal ceroid lipofuscinosis in a Border Collie. J. Vet. Med. Sci. 2004, 66, 1453–1456. [Google Scholar] [CrossRef] [Green Version]
- Mizukami, K.; Chang, H.S.; Yabuki, A.; Kawamichi, T.; Kawahara, N.; Hayashi, D.; Hossain, M.A.; Rahman, M.M.; Uddin, M.M.; Yamato, O. Novel rapid genotyping assays for neuronal ceroid lipofuscinosis in Border Collie dogs and high frequency of the mutant allele in Japan. J. Vet. Diagn. Investig. 2011, 23, 1131–1139. [Google Scholar] [CrossRef] [Green Version]
- Mizukami, K.; Kawamichi, T.; Koie, H.; Tamura, S.; Matsunaga, S.; Imamoto, S.; Saito, M.; Hasegawa, D.; Matsuki, N.; Tamahara, S.; et al. Neuronal ceroid lipofuscinosis in Border Collie dogs in Japan: Clinical and molecular epidemiological study (2000–2011). Sci. World J. 2012, 2012, 383174. [Google Scholar] [CrossRef] [Green Version]
- Minatel, L.; Underwood, S.C.; Carfagnini, J.C. Ceroid-lipofuscinosis in a Cocker Spaniel dog. Vet. Pathol. 2000, 37, 488–490. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.; Katz, M.L.; Levesque, D.; Shelton, G.D.; de Lahunta, A.; O’Brien, D.P. A variant form of neuronal ceroid lipofuscinosis in American Bulldogs. J. Vet. Intern. Med. 2005, 19, 44–51. [Google Scholar] [CrossRef]
- O’Brien, D.P.; Katz, M.L. Neuronal ceroid lipofuscinosis in 3 Australian Shepherd littermates. J. Vet. Intern. Med. 2008, 22, 472–475. [Google Scholar] [CrossRef]
- Katz, M.L.; Farias, F.H.; Sanders, D.N.; Zeng, R.; Khan, S.; Johnson, G.S.; O’Brien, D.P. A missense mutation in canine CLN6 in an Australian Shepherd with neuronal ceroid lipofuscinosis. J. Biomed. Biotechnol. 2011, 2011, 198042. [Google Scholar] [CrossRef] [Green Version]
- Nakamoto, Y.; Yamato, O.; Uchida, K.; Nibe, K.; Tamura, S.; Ozawa, T.; Ueoka, N.; Nukaya, A.; Yabuki, A.; Nakaichi, M. Neuronal ceroid-lipofuscinosis in Longhaired Chihuahuas: Clinical, pathologic, and MRI findings. J. Am. Anim. Hosp. Assoc. 2011, 47, e64–e70. [Google Scholar] [CrossRef]
- Ashwini, A.; D’Angelo, A.; Yamato, O.; Giordano, C.; Cagnotti, G.; Harcourt-Brown, T.; Mhlanga-Mutangadura, T.; Guo, J.; Johnson, G.S.; Katz, M.L. Neuronal ceroid lipofuscinosis associated with an MFSD8 mutation in Chihuahuas. Mol. Genet. Metab. 2016, 118, 326–332. [Google Scholar] [CrossRef]
- Kolicheski, A.; Johnson, G.S.; O’Brien, D.P.; Mhlanga-Mutangadura, T.; Gilliam, D.; Guo, J.; Anderson-Sieg, T.D.; Schnabel, R.D.; Taylor, J.F.; Lebowitz, A.; et al. Australian cattle dogs with neuronal ceroid lipofuscinosis are homozygous for a CLN5 nonsense mutation previously identified in Border Collies. J. Vet. Intern. Med. 2016, 30, 1149–1158. [Google Scholar] [CrossRef] [Green Version]
- Schmutz, I.; Jagannathan, V.; Bartenschlager, F.; Stein, V.M.; Gruber, A.D.; Leeb, T.; Katz, M.L. ATP13A2 missense variant in Australian Cattle Dogs with late onset neuronal ceroid lipofuscinosis. Mol. Genet. Metab. 2019, 127, 95–106. [Google Scholar] [CrossRef]
- Katz, M.L.; Narfström, K.; Johnson, G.S.; O’Brien, D.P. Assessment of retinal function and characterization of lysosomal storage body accumulation in the retinas and brains of Tibetan terriers with ceroid-lipofuscinosis. Am. J. Vet. Res. 2005, 66, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Farias, F.H.G.; Zeng, R.; Johnson, G.S.; Wininger, F.A.; Taylor, J.F.; Schnabel, R.D.; McKay, S.D.; Sanders, D.N.; Lohi, H.; Seppälä, E.H.; et al. A truncating mutation in ATP13A2 is responsible for adult-onset neuronal ceroid lipofuscinosis in Tibetan terriers. Neurobiol. Dis. 2011, 42, 468–474. [Google Scholar] [CrossRef]
- Wöhlke, A.; Philipp, U.; Bock, P.; Beineke, A.; Lichtner, P.; Meitinger, T.; Distl, O. A one base pair deletion in the canine ATP13A2 gene causes exon skipping and late-onset neuronal ceroid lipofuscinosis in the Tibetan Terrier. PLOS Genet. 2011, 7, e1002304. [Google Scholar] [CrossRef] [Green Version]
- Drögemüller, C.; Wöhlke, A.; Distl, O. Evaluation of the canine TPP1 gene as a candidate for neuronal ceroid lipofuscinosis in Tibetan Terrier and Polish Owczarek Nizinny dogs. Anim. Genet. 2005, 36, 178–179. [Google Scholar] [CrossRef]
- Abitbol, M.; Thibaud, J.L.; Olby, N.J.; Hitte, C.; Puech, J.P.; Maurer, M.; Pilot-Storck, F.; Hédan, B.; Dréano, S.; Brahimi, S.; et al. A canine Arylsulfatase G (ARSG) mutation leading to a sulfatase deficiency is associated with neuronal ceroid lipofuscinosis. Proc. Natl. Acad. Sci. USA 2010, 107, 14775–14780. [Google Scholar] [CrossRef] [Green Version]
- Kolicheski, A.; Barnes Heller, H.L.; Arnold, S.; Schnabel, R.D.; Taylor, J.F.; Knox, C.A.; Mhlanga-Mutangadura, T.; O’Brien, D.P.; Johnson, G.S.; Dreyfus, J.; et al. Homozygous PPT1 splice donor mutation in a Cane Corso dog with neuronal ceroid lipofuscinosis. J. Vet. Intern. Med. 2017, 31, 149–157. [Google Scholar] [CrossRef]
- Hirz, M.; Drögemüller, M.; Schänzer, A.; Jagannathan, V.; Dietschi, E.; Goebel, H.H.; Hecht, W.; Laubner, S.; Schmidt, M.J.; Steffen, F.; et al. Neuronal ceroid lipofuscinosis (NCL) is caused by the entire deletion of CLN8 in the Alpenländische Dachsbracke dog. Mol. Genet. Metab. 2017, 120, 269–277. [Google Scholar] [CrossRef]
- Guo, J.; Johnson, G.S.; Cook, J.; Harris, O.K.; Mhlanga-Mutangadura, T.; Schnabel, R.D.; Jensen, C.A.; Katz, M.L. Neuronal ceroid lipofuscinosis in a German Shorthaired Pointer associated with a previously reported CLN8 nonsense variant. Mol. Genet. Metab. Rep. 2019, 21, 100521. [Google Scholar] [CrossRef]
- Gilliam, D.; Kolicheski, A.; Johnson, G.S.; Mhlanga-Mutangadura, T.; Taylor, J.F.; Schnabel, R.D.; Katz, M.L. Golden Retriever dogs with neuronal ceroid lipofuscinosis have a two-base-pair deletion and frameshift in CLN5. Mol. Genet. Metab. 2015, 115, 101–109. [Google Scholar] [CrossRef]
- Bichsel, P.; Vandevelde, M. A case of ceroid-liposuscinosis in a Yugoslavian shepherd dog. Schweiz. Arch. Tierheilkd. 1982, 124, 413–418. (In French) [Google Scholar]
- Goebel, H.H.; Bilzer, T.; Malkusch, F. Morphological studies in canine (Dalmetian) neuronal ceroid-lipofuscinosis. Am. J. Med. Genet. Suppl. 1998, 5, 127–139. [Google Scholar]
- Tamura, S.; Tsuboi, M.; Ueoka, N.; Doi, S.; Tamura, Y.; Uchida, K.; Yabuki, A.; Yamato, O. Adult-onset neuronal ceroid lipofuscinosis in a Shikoku Inu. Vet. Sci. 2021, 8, 227. [Google Scholar] [CrossRef]
- Guo, J.; Johnson, G.S.; Brown, H.A.; Provencher, M.L.; da Costa, R.C.; Mhlanga-Mutangadura, T.; Taylor, J.F.; Schnabel, R.D.; O’Brien, D.P.; Katz, M.L. A CLN8 nonsense mutation in the whole genome sequence of a mixed breed dog with neuronal ceroid lipofuscinosis and Australian Shepherd ancestry. Mol. Genet. Metab. 2014, 112, 302–309. [Google Scholar] [CrossRef]
- Villani, N.A.; Bullock, G.; Michaels, J.R.; Yamato, O.; O’Brien, D.P.; Mhlanga-Mutangadura, T.; Johnson, G.S.; Katz, M.L. A mixed breed dog with neuronal ceroid lipofuscinosis is homozygous for a CLN5 nonsense mutation previously identified in Broder Collies and Australian Cattle Dogs. Mol. Genet. Metab. 2019, 127, 107–115. [Google Scholar] [CrossRef]
- Rac, R.; Giesecke, P.R. Letter: Lysosomal storage disease in chihuahuas. Aust. Vet. J. 1975, 51, 403–404. [Google Scholar] [PubMed]
- Jolly, R.D.; Hartley, W.J. Storage diseases of domestic animals. Aust. Vet. J. 1977, 53, 1–8. [Google Scholar] [CrossRef]
- Kuwamura, M.; Hattori, R.; Yamate, J.; Kotani, T.; Sasai, K. Neuronal ceroid-lipofuscinosis and hydrocephalus in a chihuahua. J. Small Anim. Pract. 2003, 44, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Faller, K.M.E.; Bras, J.; Sharpe, S.J.; Anderson, G.W.; Darwent, L.; Kun-Rodrigues, C.; Alroy, J.; Penderis, J.; Mole, S.E.; Gutierrez-Quintana, R.; et al. The Chihuahua dog: A new animal model for neuronal ceroid lipofuscinosis CLN7 disease? J. Neurosci. Res. 2016, 94, 339–347. [Google Scholar] [CrossRef] [Green Version]
- Karli, P.; Oevermann, A.; Bauer, A.; Jagannathan, V.; Leeb, T. MFSD8 single-base pair deletion in a Chihuahua with neuronal ceroid lipofuscinosis. Anim. Genet. 2016, 47, 631. [Google Scholar] [CrossRef]
- von Kleist, L.; Ariunbat, K.; Braren, I.; Stauber, T.; Storch, S.; Danyukova, T. A newly generated neuronal cell model of CLN7 disease reveals aberrant lysosome motility and impaired cell survival. Mol. Genet. Metab. 2019, 126, 196–205. [Google Scholar] [CrossRef]
- Rahman, M.M.; Yabuki, A.; Kohyama, M.; Mitani, S.; Mizukami, K.; Uddin, M.M.; Chang, H.S.; Kushida, K.; Kishimoto, M.; Yamabe, R.; et al. Real-time PCR genotyping assay for GM2 gangliosidosis variant 0 in Toy Poodles and the mutant allele frequency in Japan. J. Vet. Med. Sci. 2014, 76, 295–299. [Google Scholar] [CrossRef] [Green Version]
- Uddin, M.M.; Arata, S.; Takeuchi, Y.; Chang, H.S.; Mizukami, K.; Yabuki, A.; Rahman, M.M.; Kohyama, M.; Hossain, M.A.; Takayama, K.; et al. Molecular epidemiology of canine GM1 gangliosidosis in the Shiba Inu breed in Japan: Relationship between regional prevalence and carrier frequency. BMC Vet. Res. 2013, 9, 132. [Google Scholar] [CrossRef] [Green Version]
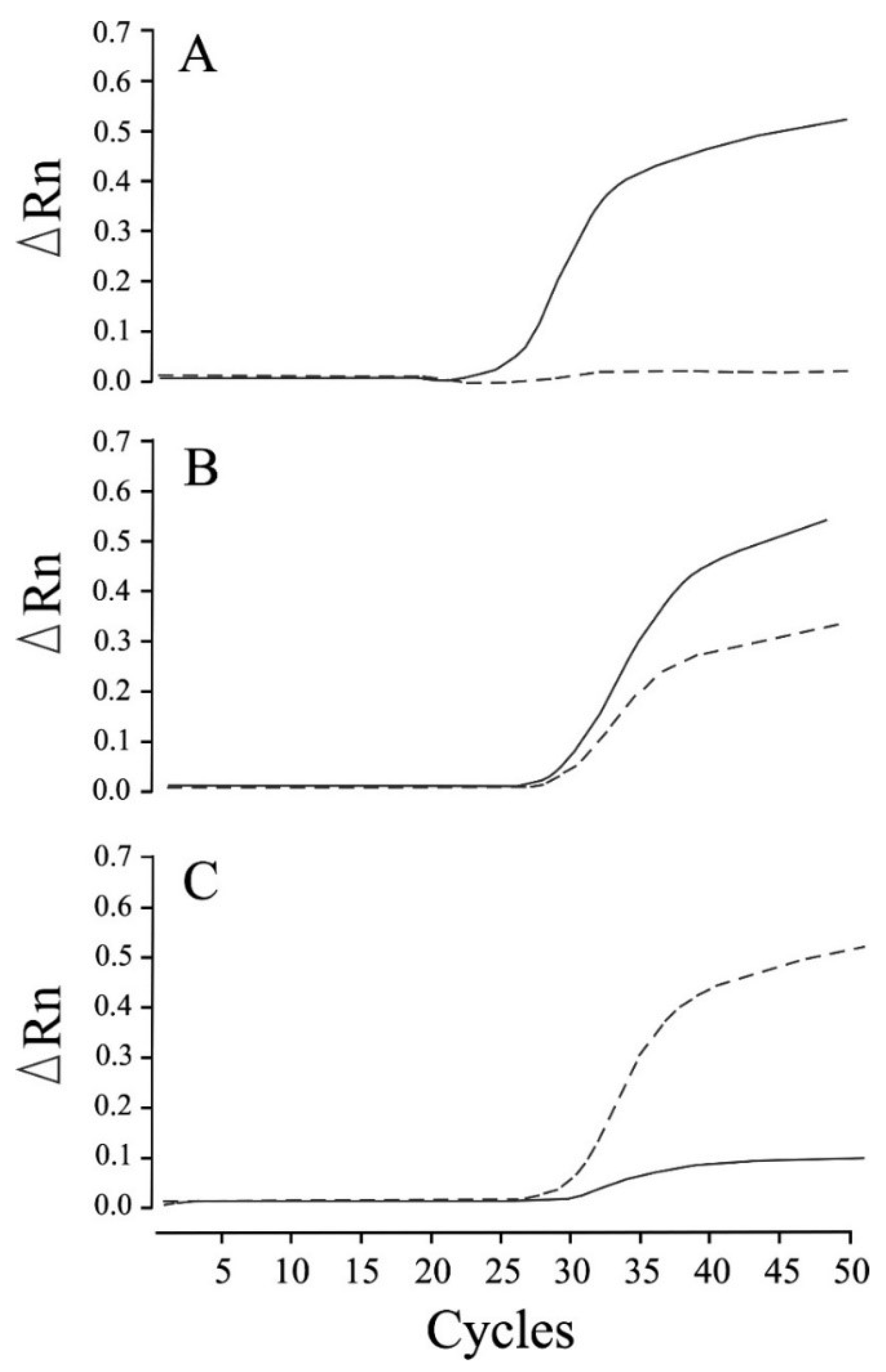
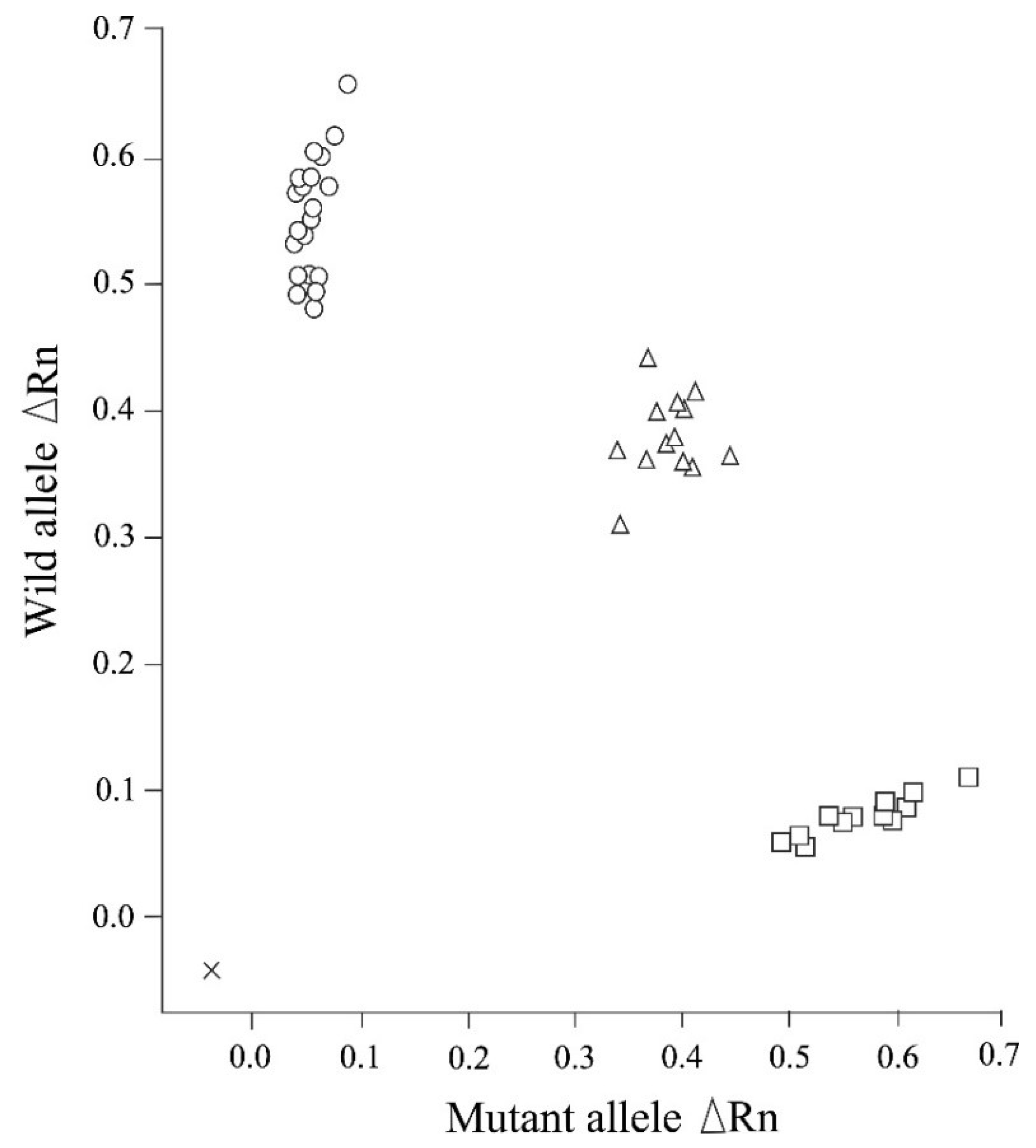

{kind=link}
{kind=link}
| Primer/Probe | Sequence 5′ to 3′ (mer) | Position | Reporter (5′) | Quencher (3′) | Tm (°C) | Concentration (nM) |
|---|---|---|---|---|---|---|
| Real-time PCR: | ||||||
| Forward primer | GCTGTTGTGGCCACTAATATTGTG (24) | c.805_828 | NA | NA | 66.1 | 450 |
| Reverse primer | AGAATAAAACTTACGTTTCAAAAAGGGCAA (30) | c.851_866+14 | NA | NA | 68.1 | 450 |
| Probe for wild-type allele | TTCGTGATTCTATTTATCT (19) | c.832_850 | FAM | NFQ | 48.4 | 100 |
| Probe for mutant allele | TTTTTCGTGATTCTATTATCT (21) | c.829_850 | VIC | NFQ | 52.3 | 100 |
| Sanger sequencing: | ||||||
| Forward primer | GTCATAGAATTTGCTACATATAATTTC (27) | Intron 7 | NA | NA | 57.1 | NA |
| Reverse primer | GTTTTGAGAACATTGATATGCTTGAT (26) | Intron 8 | NA | NA | 62.9 | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pervin, S.; Islam, M.S.; Tada, N.; Tsutsui, T.; Rahman, M.M.; Yabuki, A.; Tacharina, M.R.; Rakib, T.M.; Maki, S.; Yamato, O. Screening and Carrier Rate of Neuronal Ceroid Lipofuscinosis in Chihuahua Dogs in Japan. Animals 2022, 12, 1210. https://doi.org/10.3390/ani12091210
Pervin S, Islam MS, Tada N, Tsutsui T, Rahman MM, Yabuki A, Tacharina MR, Rakib TM, Maki S, Yamato O. Screening and Carrier Rate of Neuronal Ceroid Lipofuscinosis in Chihuahua Dogs in Japan. Animals. 2022; 12(9):1210. https://doi.org/10.3390/ani12091210
Chicago/Turabian StylePervin, Shahnaj, Md Shafiqul Islam, Naomi Tada, Toshihiko Tsutsui, Mohammad Mahbubur Rahman, Akira Yabuki, Martia Rani Tacharina, Tofazzal Md Rakib, Shinichiro Maki, and Osamu Yamato. 2022. "Screening and Carrier Rate of Neuronal Ceroid Lipofuscinosis in Chihuahua Dogs in Japan" Animals 12, no. 9: 1210. https://doi.org/10.3390/ani12091210
APA StylePervin, S., Islam, M. S., Tada, N., Tsutsui, T., Rahman, M. M., Yabuki, A., Tacharina, M. R., Rakib, T. M., Maki, S., & Yamato, O. (2022). Screening and Carrier Rate of Neuronal Ceroid Lipofuscinosis in Chihuahua Dogs in Japan. Animals, 12(9), 1210. https://doi.org/10.3390/ani12091210