
Attachment towards the Owner Is Associated with Spontaneous Sleep EEG Parameters in Family Dogs


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Behavioral Assessment
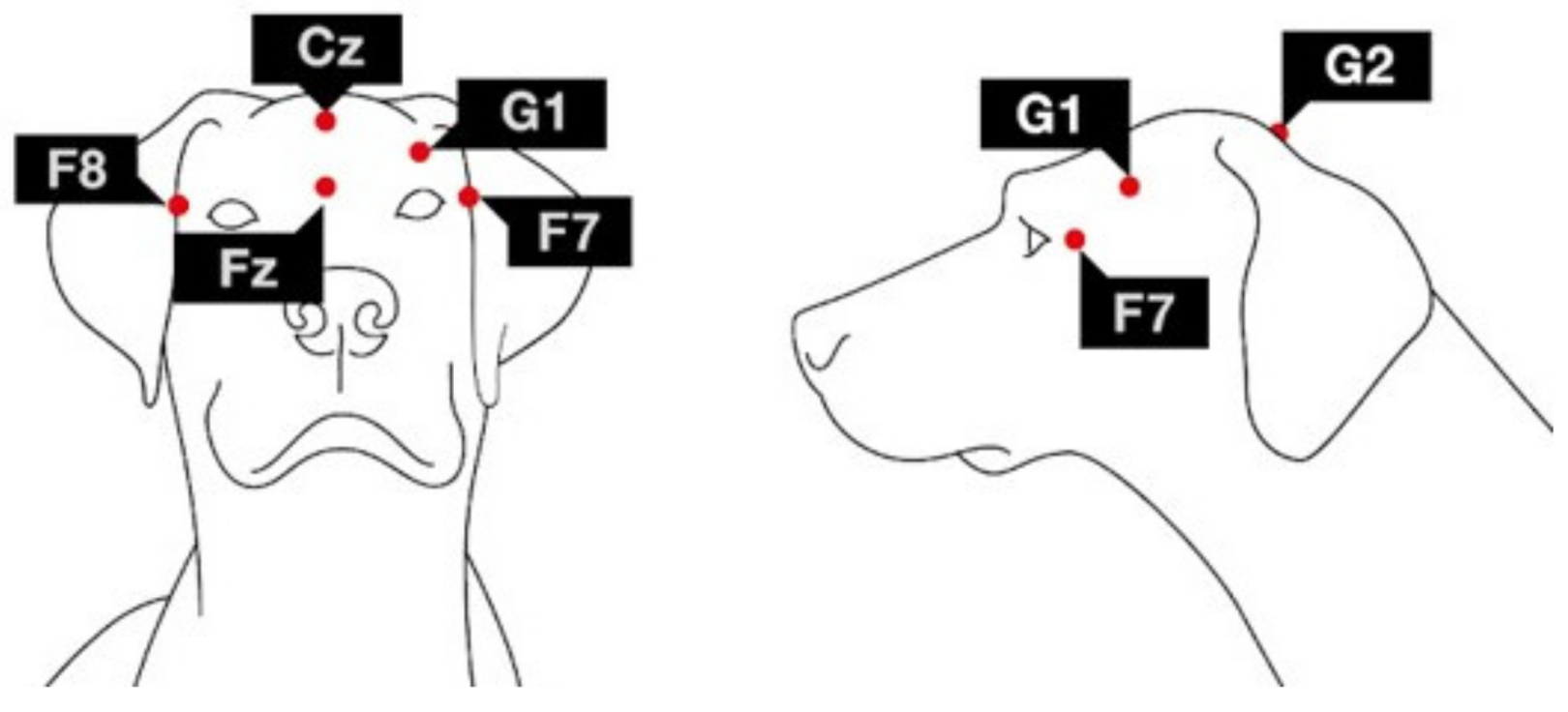
2.3. Sleep EEG Recording
- (1)
- In the case of 25 dogs (59.5% of the total sample), the signal was collected, amplified and digitized at a sampling rate of 1000 Hz/channel using the 40-channels NuAmps amplifier (© 2018 Compumedics Neuroscan) and DC-recording, later saved in .cnt format with the Scan 4.3 Acquire software (© 2018 Compumedics Neuroscan) then converted to .edf format using MatLab EEG Toolbox.
- (2)
- In the case of 17 dogs (40.5% of the total sample), the signal was collected, pre-filtered, amplified and digitized with a sampling rate of 1024 Hz/channel using a SAM 25 R style MicroMed Headbox (MicroMed Inc., Houston, TX, USA). The passband was set at 0.5–256 Hz, using System Plus Evolution software (MicroMed Inc, Houston, TX, USA), which exported data in .edf format.
2.4. Data Analysis
2.5. Statistical Analysis
3. Results
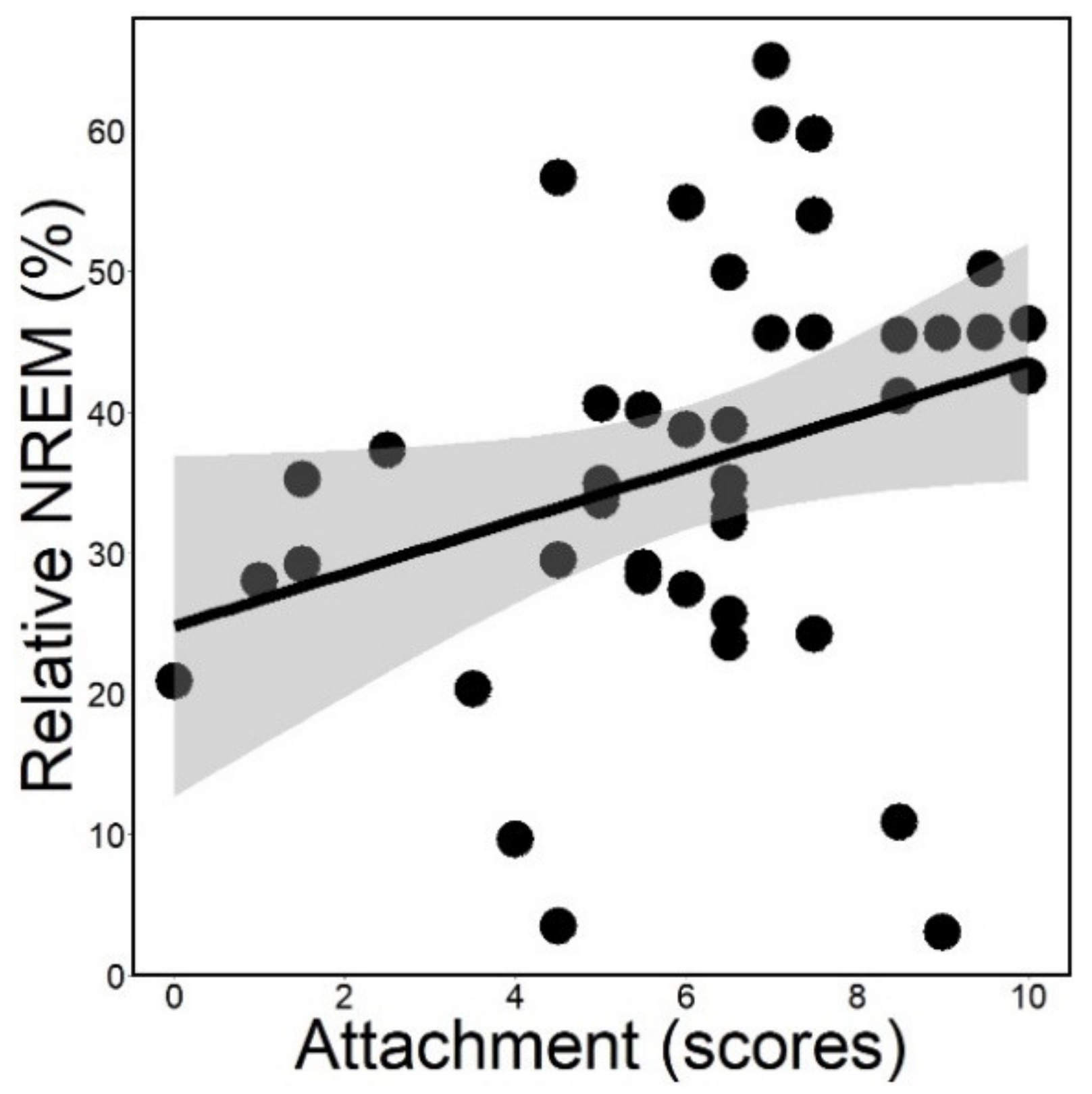
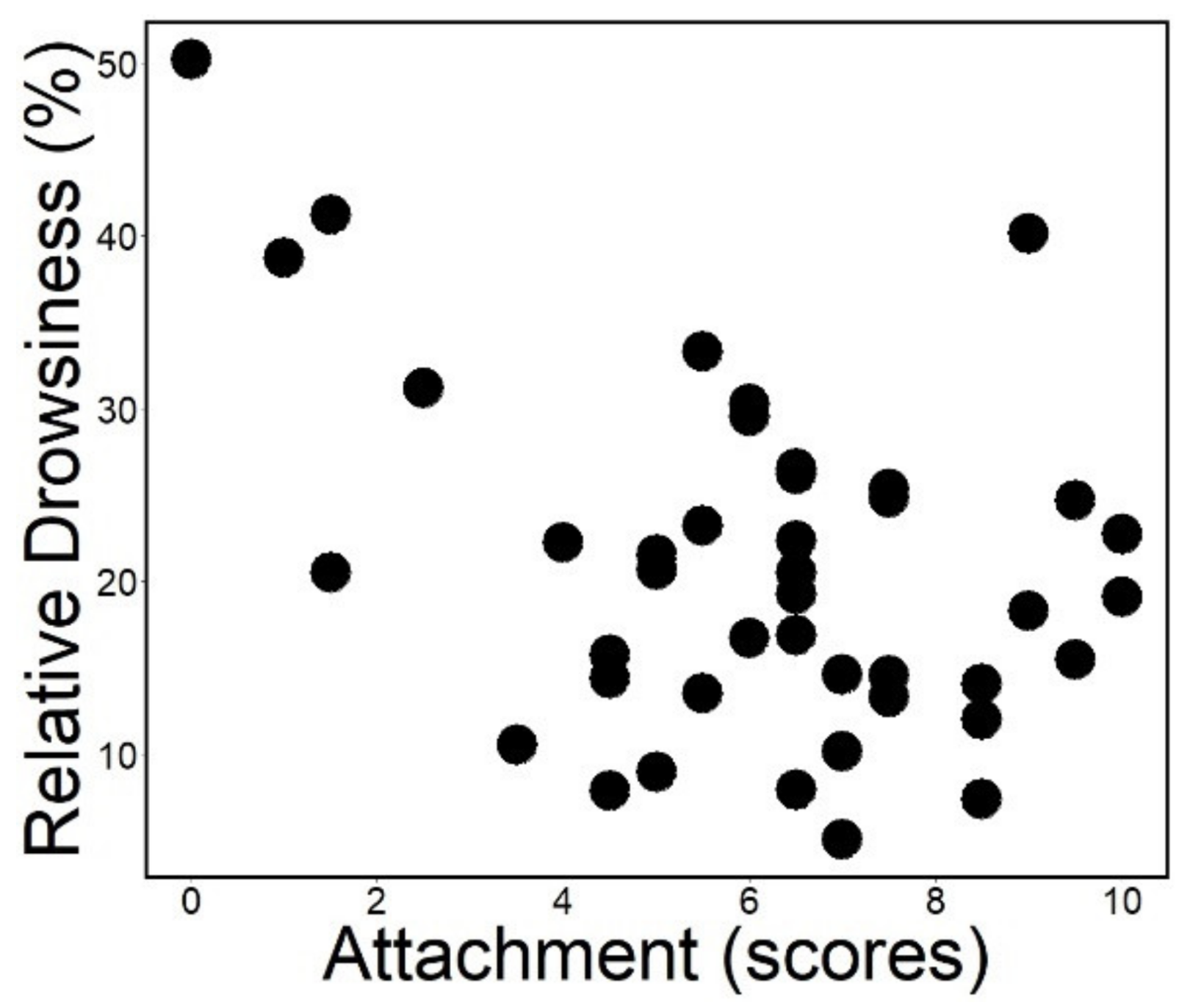
3.1. Sleep Macrostructure
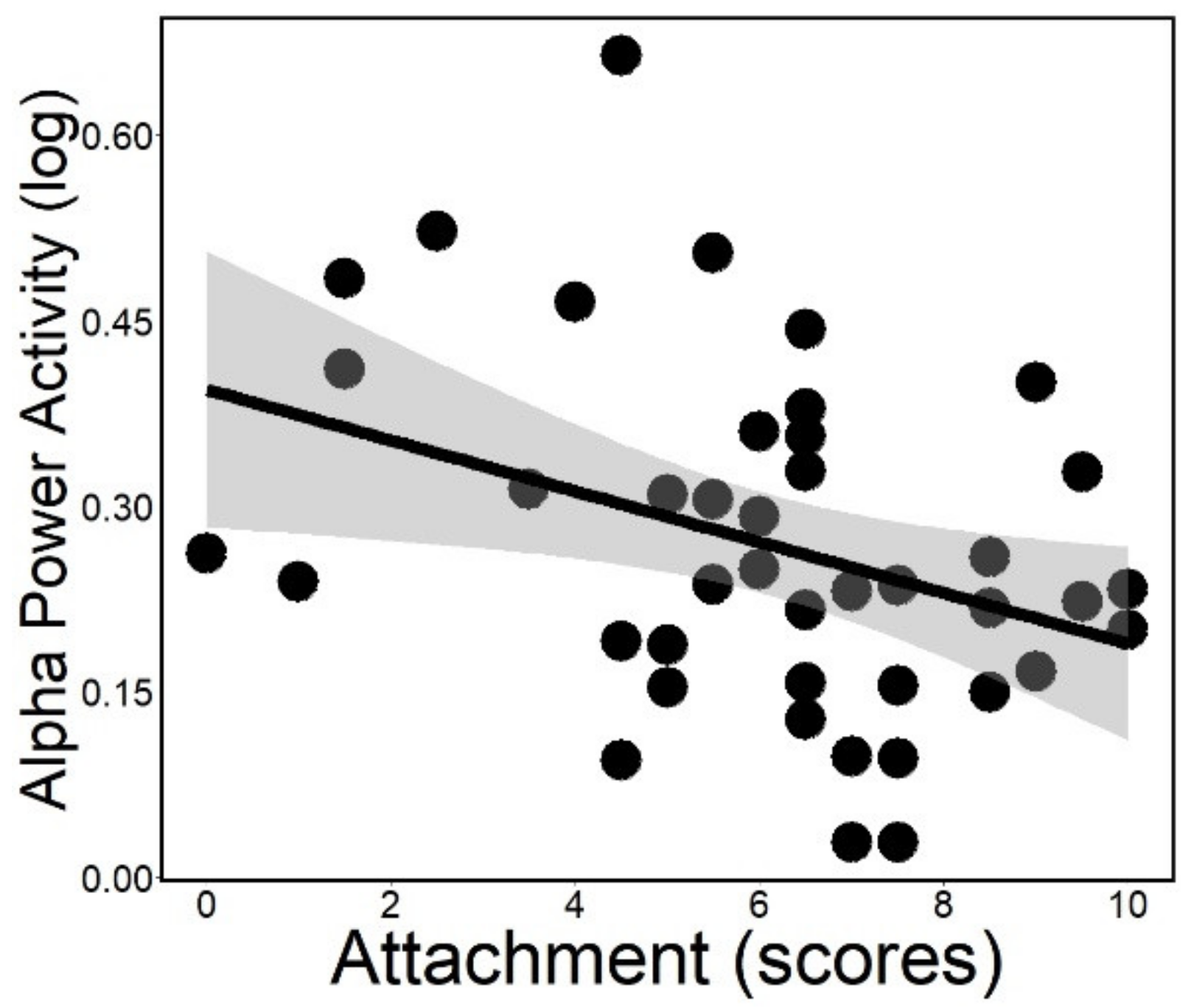
3.2. Spectral Analysis (in NREM Sleep)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bowlby, J. Attachment and Loss: Volume 1 Attachment; Basic Books: New York, NY, USA, 1969. [Google Scholar]
- Ainsworth, M.S. Attachments beyond Infancy. Am. Psychol. 1989, 44, 709. [Google Scholar] [CrossRef] [PubMed]
- Vrtička, P.; Vuilleumier, P. Neuroscience of Human Social Interactions and Adult Attachment Style. Front. Hum. Neurosci. 2012, 6, 212. [Google Scholar] [CrossRef] [PubMed]
- Collins, N.L.; Feeney, B.C. Working Models of Attachment Shape Perceptions of Social Support: Evidence from Experimental and Observational Studies. J. Personal. Soc. Psychol. 2004, 87, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Noftle, E.E.; Shaver, P.R. Attachment Dimensions and the Big Five Personality Traits: Associations and Comparative Ability to Predict Relationship Quality. J. Res. Personal. 2006, 40, 179–208. [Google Scholar] [CrossRef]
- Muris, P.; Meesters, C.; van Melick, M.; Zwambag, L. Self-Reported Attachment Style, Attachment Quality, and Symptoms of Anxiety and Depression in Young Adolescents. Personal. Individ. Differ. 2001, 30, 809–818. [Google Scholar] [CrossRef]
- Adams, G.C.; Stoops, M.A.; Skomro, R.P. Sleep Tight: Exploring the Relationship between Sleep and Attachment Style across the Life Span. Sleep Med. Rev. 2014, 18, 495–507. [Google Scholar] [CrossRef]
- Sloan, E.P.; Maunder, R.G.; Hunter, J.J.; Moldofsky, H. Insecure Attachment Is Associated with the α-EEG Anomaly during Sleep. BioPsychoSoc. Med. 2007, 1, 20. [Google Scholar] [CrossRef]
- Troxel, W.M.; Cyranowski, J.M.; Hall, M.; Frank, E.; Buysse, D.J. Attachment Anxiety, Relationship Context, and Sleep in Women with Recurrent Major Depression. Psychosom. Med. 2007, 69, 692–699. [Google Scholar] [CrossRef]
- Field, T.; Reite, M. Children’s Responses to Separation from Mother during the Birth of Another Child. Child Dev. 1984, 55, 1308–1316. [Google Scholar] [CrossRef]
- Ramesh, V.; Kaushal, N.; Gozal, D. Sleep Fragmentation Differentially Modifies EEG Delta Power during Slow Wave Sleep in Socially Isolated and Paired Mice. Sleep Sci. 2009, 2, 64–75. [Google Scholar]
- Knyazev, G.G.; Schutter, D.J.L.G.; van Honk, J. Anxious Apprehension Increases Coupling of Delta and Beta Oscillations. Int. J. Psychophysiol. 2006, 61, 283–287. [Google Scholar] [CrossRef]
- Knyazev, G.G.; Slobodskaya, H.R. Personality Trait of Behavioral Inhibition Is Associated with Oscillatory Systems Reciprocal Relationships. Int. J. Psychophysiol. 2003, 48, 247–261. [Google Scholar] [CrossRef]
- Butt, M.; Quan, S.F.; Pentland, A.S.; Khayal, I. Gender Differences in Real-Home Sleep of Young and Older Couples. Southwest J. Pulm. Crit. Care 2015, 10, 289. [Google Scholar] [CrossRef][Green Version]
- Monroe, L.J. Transient Changes in EEG Sleep Patterns of Married Good Sleepers: The Effects of Altering Sleeping Arrangement. Psychophysiology 1969, 6, 330–337. [Google Scholar] [CrossRef]
- Bunford, N.; Reicher, V.; Kis, A.; Pogány, Á.; Gombos, F.; Bódizs, R.; Gácsi, M. Differences in Pre-Sleep Activity and Sleep Location Are Associated with Variability in Daytime/Nighttime Sleep Electrophysiology in the Domestic Dog. Sci. Rep. 2018, 8, 7109. [Google Scholar] [CrossRef]
- Andics, A.; Miklósi, Á. Neural Processes of Vocal Social Perception: Dog-Human Comparative FMRI Studies. Neurosci. Biobehav. Rev. 2018, 85, 54–64. [Google Scholar] [CrossRef]
- Bunford, N.; Andics, A.; Kis, A.; Miklósi, Á.; Gácsi, M. Canis Familiaris as a Model for Non-Invasive Comparative Neuroscience. Trends Neurosci. 2017, 40, 438–452. [Google Scholar] [CrossRef]
- Miklósi, Á. Dog Behaviour Evolution and Cognition; Oxford University Press: New York, NY, USA, 2014. [Google Scholar]
- Topál, J.; Gácsi, M.; Miklósi, Á.; Virányi, Z.; Kubinyi, E.; Csányi, V. Attachment to Humans: A Comparative Study on Hand-Reared Wolves and Differently Socialized Dog Puppies. Anim. Behav. 2005, 70, 1367–1375. [Google Scholar] [CrossRef]
- Pettijohn, T.F. Attachment and Separation Distress in the Infant Guinea Pig. Dev. Psychobiol. 1979, 12, 73–81. [Google Scholar] [CrossRef]
- Coe, C.L.; Mendoza, S.P.; Smotherman, W.P.; Levine, S. Mother-Infant Attachment in the Squirrel Monkey: Adrenal Response to Separation. Behav. Biol. 1978, 22, 256–263. [Google Scholar] [CrossRef]
- Topál, J.; Miklósi, Á.; Csányi, V.; Dóka, A. Attachment Behavior in Dogs (Canis familiaris): A New Application of Ainsworth’s (1969) Strange Situation Test. J. Comp. Psychol. 1998, 112, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Gácsi, M.; Topál, J.; Miklósi, Á.; Dóka, A.; Csányi, V. Attachment Behavior of Adult Dogs (Canis familiaris) Living at Rescue Centers: Forming New Bonds. J. Comp. Psychol. 2001, 115, 423. [Google Scholar] [CrossRef] [PubMed]
- Topál, J.; Gácsi, M. Lessons We Should Learn from Our Unique Relationship with Dogs: An Ethological Approach. In Crossing Boundaries; Birke, L., Hockenhull, J., Eds.; Brill Academic Press: Leiden, The Netherlands, 2012; pp. 161–186. [Google Scholar] [CrossRef]
- Palmer, R.; Custance, D. A Counterbalanced Version of Ainsworth’s Strange Situation Procedure Reveals Secure-Base Effects in Dog–Human Relationships. Appl. Anim. Behav. Sci. 2008, 109, 306–319. [Google Scholar] [CrossRef]
- Gácsi, M.; Maros, K.; Sernkvist, S.; Faragó, T.; Miklósi, Á. Human Analogue Safe Haven Effect of the Owner: Behavioural and Heart Rate Response to Stressful Social Stimuli in Dogs. PLoS ONE 2013, 8, e58475. [Google Scholar] [CrossRef]
- Ternman, E.; Hänninen, L.; Pastell, M.; Agenäs, S.; Nielsen, P.P. Sleep in Dairy Cows Recorded with a Non-Invasive EEG Technique. Appl. Anim. Behav. Sci. 2012, 140, 25–32. [Google Scholar] [CrossRef]
- Williams, D.C.; Aleman, M.; Holliday, T.A.; Fletcher, D.J.; Tharp, B.; Kass, P.H.; Steffey, E.P.; LeCouteur, R.A. Qualitative and Quantitative Characteristics of the Electroencephalogram in Normal Horses during Spontaneous Drowsiness and Sleep. J. Vet. Intern. Med. 2008, 22, 630–638. [Google Scholar] [CrossRef]
- Kis, A.; Szakadát, S.; Kovács, E.; Gácsi, M.; Simor, P.; Gombos, F.; Topál, J.; Miklósi, Á.; Bódizs, R. Development of a Non-Invasive Polysomnography Technique for Dogs (Canis familiaris). Physiol. Behav. 2014, 130, 149–156. [Google Scholar] [CrossRef]
- Tóth, L.; Gácsi, M.; Miklósi, Á.; Bogner, P.; Repa, I. Awake Dog Brain Magnetic Resonance Imaging. J. Vet. Behav. 2009, 4, 50. [Google Scholar] [CrossRef]
- Karl, S.; Boch, M.; Zamansky, A.; van der Linden, D.; Wagner, I.C.; Völter, C.J.; Lamm, C.; Huber, L. Exploring the Dog–Human Relationship by Combining FMRI, Eye-Tracking and Behavioural Measures. Sci. Rep. 2020, 10, 22273. [Google Scholar] [CrossRef]
- Gábor, A.; Andics, A.; Miklósi, Á.; Czeibert, K.; Carreiro, C.; Gácsi, M. Social Relationship-Dependent Neural Response to Speech in Dogs. NeuroImage 2021, 243, 118480. [Google Scholar] [CrossRef]
- Bódizs, R.; Kis, A.; Gácsi, M.; Topál, J. Sleep in the Dog: Comparative, Behavioral and Translational Relevance. Curr. Opin. Behav. Sci. 2020, 33, 25–33. [Google Scholar] [CrossRef]
- Kis, A.; Gergely, A.; Galambos, Á.; Abdai, J.; Gombos, F.; Bódizs, R.; Topál, J. Sleep Macrostructure Is Modulated by Positive and Negative Social Experience in Adult Pet Dogs. Proc. R. Soc. B Biol. Sci. 2017, 284, 20171883. [Google Scholar] [CrossRef]
- Kurth, S.; Olini, N.; Huber, R.; LeBourgeois, M. Sleep and Early Cortical Development. Curr. Sleep Med. Rep. 2015, 1, 64–73. [Google Scholar] [CrossRef]
- Reicher, V.; Bunford, N.; Kis, A.; Carreiro, C.; Csibra, B.; Kratz, L.; Gácsi, M. Developmental Features of Sleep Electrophysiology in Family Dogs. Sci. Rep. 2021, 11, 22760. [Google Scholar] [CrossRef]
- Iotchev, I.B.; Kis, A.; Turcsán, B.; Tejeda Fernández de Lara, D.R.; Reicher, V.; Kubinyi, E. Age-Related Differences and Sexual Dimorphism in Canine Sleep Spindles. Sci. Rep. 2019, 9, 10092. [Google Scholar] [CrossRef]
- Lenkei, R.; Carreiro, C.; Gácsi, M.; Pongrácz, P. The Relationship between Functional Breed Selection and Attachment Pattern in Family Dogs (Canis familiaris). Appl. Anim. Behav. Sci. 2021, 235, 105231. [Google Scholar] [CrossRef]
- Ainsworth, M.D.S. Object Relations, Dependency, and Attachment: A Theoretical Review of the Infant-Mother Relationship. Child Dev. 1969, 40, 969–1025. [Google Scholar] [CrossRef]
- Kovács, K.; Virányi, Z.; Kis, A.; Turcsán, B.; Hudecz, Á.; Marmota, M.T.; Koller, D.; Rónai, Z.; Gácsi, M.; Topál, J. Dog-Owner Attachment Is Associated With Oxytocin Receptor Gene Polymorphisms in Both Parties. A Comparative Study on Austrian and Hungarian Border Collies. Front. Psychol. 2018, 9, 435. [Google Scholar] [CrossRef]
- Reicher, V.; Kis, A.; Simor, P.; Bódizs, R.; Gombos, F.; Gácsi, M. Repeated Afternoon Sleep Recordings Indicate First-night-effect-like Adaptation Process in Family Dogs. J. Sleep Res. 2020, 29, e12998. [Google Scholar] [CrossRef]
- Pótári, A.; Ujma, P.P.; Konrad, B.N.; Genzel, L.; Simor, P.; Körmendi, J.; Gombos, F.; Steiger, A.; Dresler, M.; Bódizs, R. Age-Related Changes in Sleep EEG Are Attenuated in Highly Intelligent Individuals. NeuroImage 2017, 146, 554–560. [Google Scholar] [CrossRef]
- Ujma, P.P.; Konrad, B.N.; Genzel, L.; Bleifuss, A.; Simor, P.; Pótári, A.; Körmendi, J.; Gombos, F.; Steiger, A.; Bódizs, R.; et al. Sleep Spindles and Intelligence: Evidence for a Sexual Dimorphism. J. Neurosci. 2014, 34, 16358–16368. [Google Scholar] [CrossRef]
- Berry, R.B.; Brooks, R.; Gamaldo, C.E.; Harding, S.M.; Marcus, C.; Vaughn, B.v. The AASM Manual for the Scoring of Sleep and Associated Events. In Rules, Terminology and Technical Specifications; American Academy of Sleep Medicine: Darien, IL, USA, 2012; Volume 176. [Google Scholar]
- Gergely, A.; Kiss, O.; Reicher, V.; Iotchev, I.; Kovács, E.; Gombos, F.; Benczúr, A.; Galambos, Á.; Topál, J.; Kis, A. Reliability of Family Dogs’ Sleep Structure Scoring Based on Manual and Automated Sleep Stage Identification. Animals 2020, 10, 927. [Google Scholar] [CrossRef]
- Knyazev, G.G.; Savostyanov, A.N.; Levin, E.A. Alpha Oscillations as a Correlate of Trait Anxiety. Int. J. Psychophysiol. 2004, 53, 147–160. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.G. Statistical Power Analyses Using G* Power 3.1: Tests for Correlation and Regression Analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef]
- Meerlo, P.; de Bruin, E.A.; Strijkstra, A.M.; Daan, S. A Social Conflict Increases EEG Slow-Wave Activity during Subsequent Sleep. Physiol. Behav. 2001, 73, 331–335. [Google Scholar] [CrossRef]
- Kalin, N.H.; Shelton, S.E.; Davidson, R.J.; Kelley, A.E. The Primate Amygdala Mediates Acute Fear but Not the Behavioral and Physiological Components of Anxious Temperament. J. Neurosci. 2001, 21, 2067–2074. [Google Scholar] [CrossRef]
- Lindsley, D.B.; Wicke, J.D. The Electroencephalogram: Autonomous Electrical Activity in Man and Animals. In Bioelectric Recording Techniques: Electroencephalography and Human Brain Potentials; Thompson, R.F., Patterson, M.M., Eds.; Academic Press: New York, NY, USA, 2013; Volume 1-B, pp. 3–84. [Google Scholar]
- Gander, M.; Buchheim, A. Attachment Classification, Psychophysiology and Frontal EEG Asymmetry across the Lifespan: A Review. Front. Hum. Neurosci. 2015, 9, 79. [Google Scholar] [CrossRef]
- Stanley, S.M. Attachment and Prefrontal Emotional Reactivity: An EEG Study of Emotional Processing in the Context of Attachment. Ph.D. Thesis, University of California, Davis, CA, USA, 2005. [Google Scholar]
- Davidson, R.J. Anterior Cerebral Asymmetry and the Nature of Emotion. Brain Cogn. 1992, 20, 125–151. [Google Scholar] [CrossRef]
- Verbeke, W.J.M.I.; Pozharliev, R.; van Strien, J.W.; Belschak, F.; Bagozzi, R.P. “I Am Resting but Rest Less Well with You.” The Moderating Effect of Anxious Attachment Style on Alpha Power during EEG Resting State in a Social Context. Front. Hum. Neurosci. 2014, 8, 486. [Google Scholar] [CrossRef]
- Knyazev, G.G.; Slobodskaya, H.R.; Wilson, G.D. Personality and Brain Oscillations: Developmental Aspects. In Advances in Psychology Research; Shohov, S.P., Ed.; Nova Science Publishers: New York, NY, USA, 2004; Volume 29, pp. 3–34. [Google Scholar]
- Meerlo, P.; Pragt, B.J.; Daan, S. Social Stress Induces High Intensity Sleep in Rats. Neurosci. Lett. 1997, 225, 41–44. [Google Scholar] [CrossRef]
- Sternthal, H.S.; Webb, W.B. Sleep Deprivation of Rats by Punitive and Non Punitive Procedures. Physiol. Behav. 1986, 37, 249–252. [Google Scholar] [CrossRef]
- Bálint, A.; Eleőd, H.; Körmendi, J.; Bódizs, R.; Reicher, V.; Gácsi, M. Potential Physiological Parameters to Indicate Inner States in Dogs: The Analysis of ECG, and Respiratory Signal during Different Sleep Phases. Front. Behav. Neurosci. 2019, 13, 207. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, L.A.; Fox, N.A.; Schulkin, J.; Gold, P.W. Behavioral and Psychophysiological Correlates of Self-Presentation in Temperamentally Shy Children. Dev. Psychobiol. 1999, 35, 119–135. [Google Scholar] [CrossRef]
- Cimarelli, G.; Turcsán, B.; Bánlaki, Z.; Range, F.; Virányi, Z. Dog Owners’ Interaction Styles: Their Components and Associations with Reactions of Pet Dogs to a Social Threat. Front. Psychol. 2016, 7, 1979. [Google Scholar] [CrossRef]
- Kis, A.; Turcsán, B.; Miklósi, Á.; Gácsi, M. The Effect of the Owner’s Personality on the Behaviour of Owner-Dog Dyads. Interact. Stud. 2012, 13, 373–385. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Episode | Variable | Behavior Description | Score |
|---|---|---|---|
| 1, 2, 4, 6 | Proximity | D is close to O (closest body part is within 1 m)—in more than 75% of the time when D is not exploring or playing | 1 |
| 1 | BlockO-1 | During the first block-carrying episode, D watches or follows O for more than half of the time | 1 |
| 2 | LeaveO-1 | When O first leaves, D follows O to the door (at least within 1 m from the door) | 1 |
| 4 | EnterO-1 | When O first enters, D approaches O at once (in reaching distance) and wags the tail | 1 |
| 4 | BlockO-2 | During the second block-carrying episode, D watches or follows O for more than half of the time | 1 |
| 4 | LeaveO-2 | When O leaves the second time, D follows O to the door (at least within 1 m from the door) | 0.5 |
| 6 | EnterO-2 | When O enters the second time, D approaches O at once (in reaching distance) and wags the tail/jumps/spins | 0.5 |
| 3 | DoorS-1 | D stands by or orients at O’s door (for at least 5 s—score 0.5; almost all the time—score 1) during first separation | 1 |
| 3 | NoPlayS | D does not play with S, although D played with S for more than 10 s in Episode 2 (in O’s presence) | 1 |
| 3, 5 | VocalizeS | D vocalizes (any occurrence, except asking S for ball) | 0.5 |
| 3, 5 | Chair | D is mostly (for more than half of the time) at O’s chair if not at the door | 0.5 |
| 5 | DoorS-2 | D stands by or orients at O’s door (for at least 5 s) during second separation | 1 |
| 5 | EscapeS | When S enters, D at first tries to approach the door opening (to sneak out through the door) instead of greeting S | 0.5 |
| 6 | DoorS-3 | D stands by or orients at O’s door (for at least 5 s) during third separation | 0.5 |
| sum | 11 |
| Assessment | Variable | Measure |
|---|---|---|
| SST 1 factor | Attachment | sum of the behavior items referring to dog’s attachment towards the owner (score range from 0–11) |
| Sleep macrostructure | Relative drowsiness | time spent in drowsiness/record duration (%) |
| Relative NREM 2 | time spent in NREM/record duration (%) | |
| NREM spectrum | Alpha band | relative power of the frequency range 8–12 Hz 3 (log 4) |
| Alpha–delta anticorrelation | ratio of relative powers of alpha (8–12 Hz) and delta (1–4 Hz) (log) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carreiro, C.; Reicher, V.; Kis, A.; Gácsi, M. Attachment towards the Owner Is Associated with Spontaneous Sleep EEG Parameters in Family Dogs. Animals 2022, 12, 895. https://doi.org/10.3390/ani12070895
Carreiro C, Reicher V, Kis A, Gácsi M. Attachment towards the Owner Is Associated with Spontaneous Sleep EEG Parameters in Family Dogs. Animals. 2022; 12(7):895. https://doi.org/10.3390/ani12070895
Chicago/Turabian StyleCarreiro, Cecília, Vivien Reicher, Anna Kis, and Márta Gácsi. 2022. "Attachment towards the Owner Is Associated with Spontaneous Sleep EEG Parameters in Family Dogs" Animals 12, no. 7: 895. https://doi.org/10.3390/ani12070895
APA StyleCarreiro, C., Reicher, V., Kis, A., & Gácsi, M. (2022). Attachment towards the Owner Is Associated with Spontaneous Sleep EEG Parameters in Family Dogs. Animals, 12(7), 895. https://doi.org/10.3390/ani12070895