
Assisted Reproduction Techniques to Improve Reproduction in a Non-Model Species: The Case of the Arabian Bustard (Ardeotis arabs) Conservation Breeding Program

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
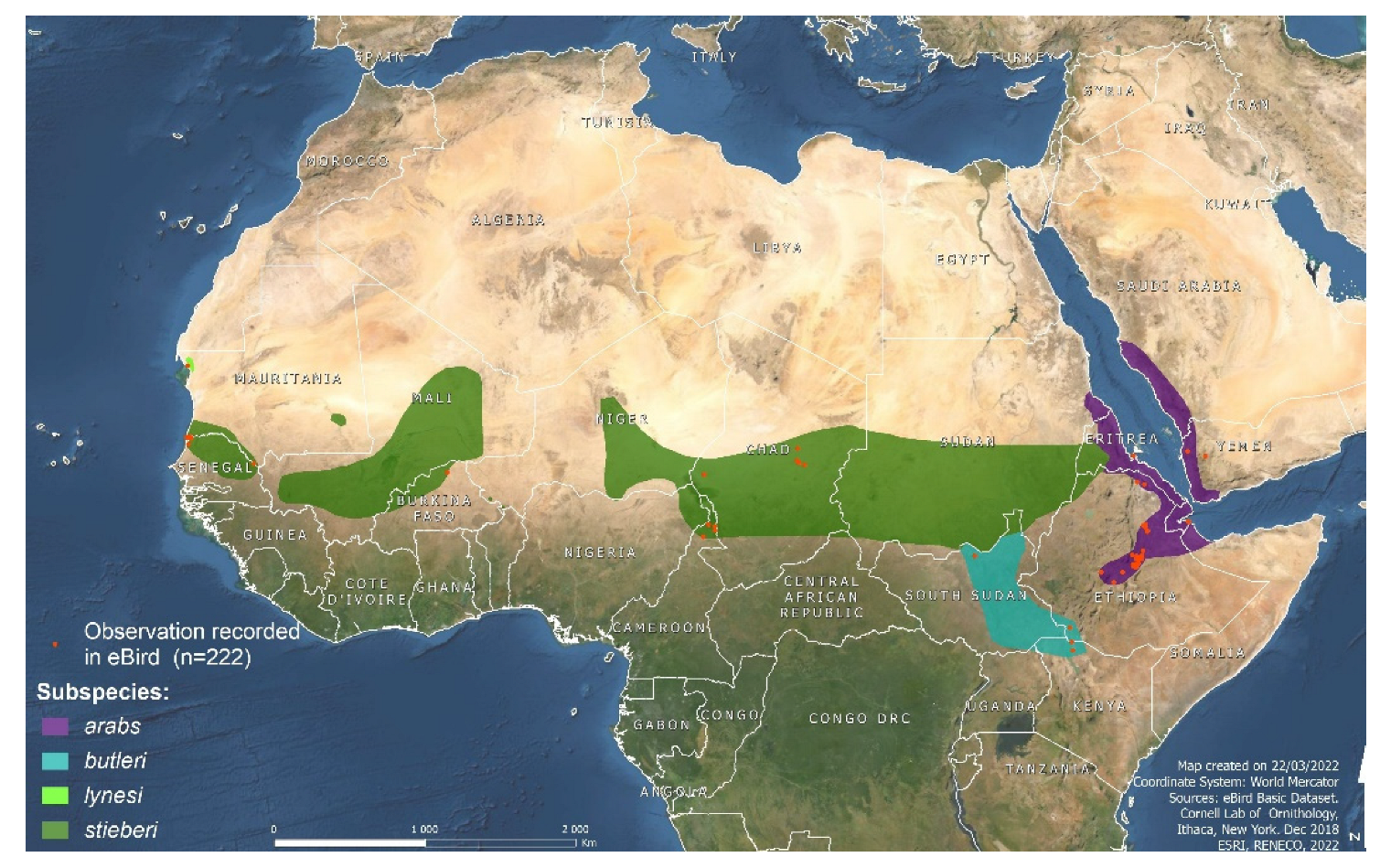
2.1. Experimental Birds and Breeding Program Management
2.2. Artificial Reproduction
2.2.1. Semen Collection
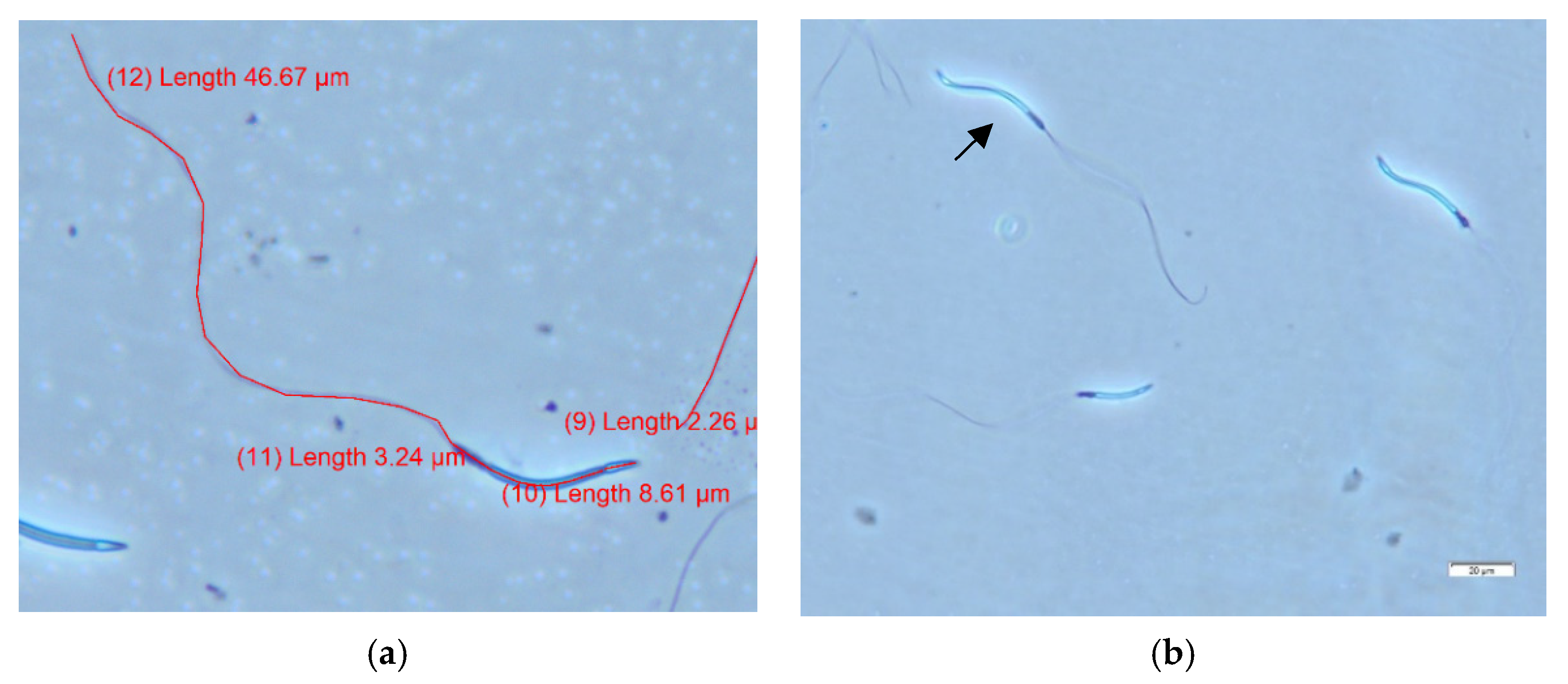
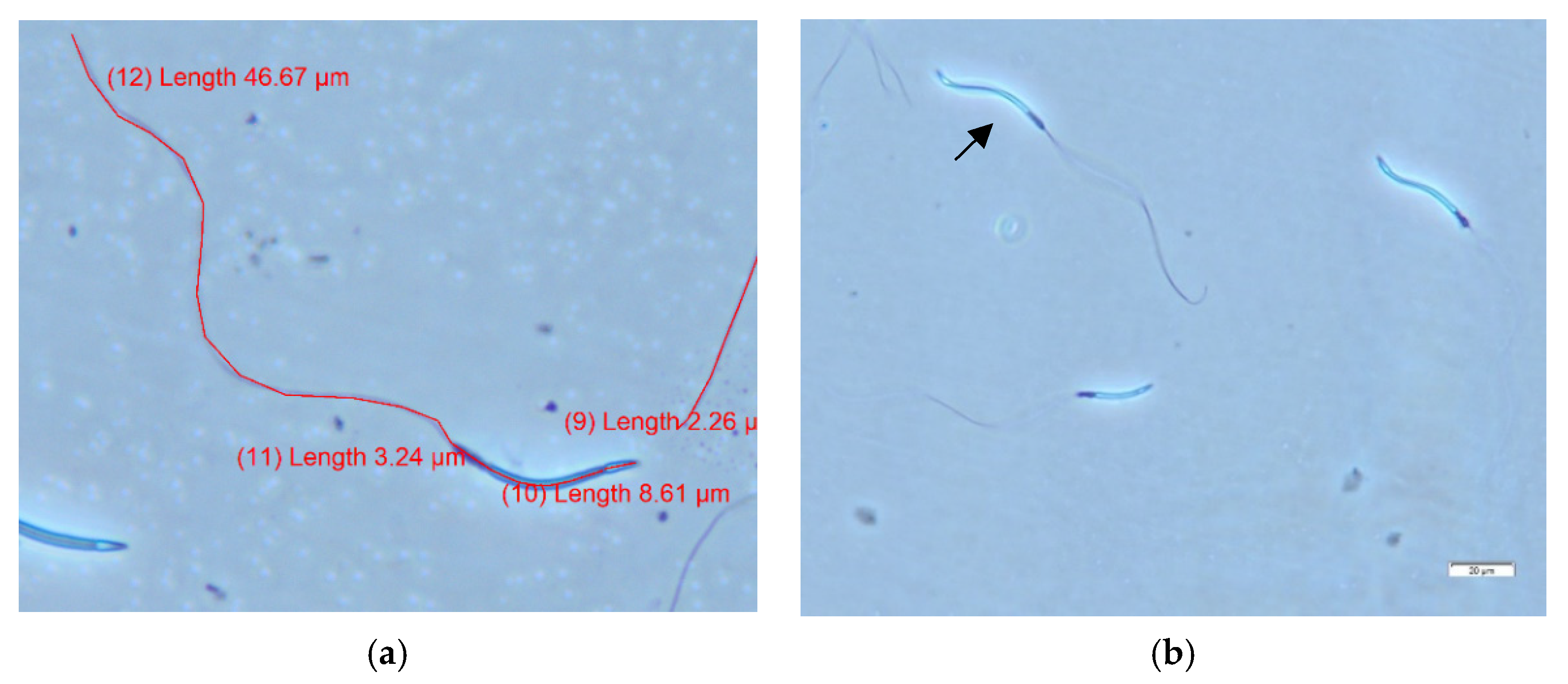
2.2.2. Semen Analyses
2.2.3. Semen Refrigeration for 24 h
2.2.4. Semen Cryopreservation and Thawing
2.2.5. Artificial Insemination
3. Results
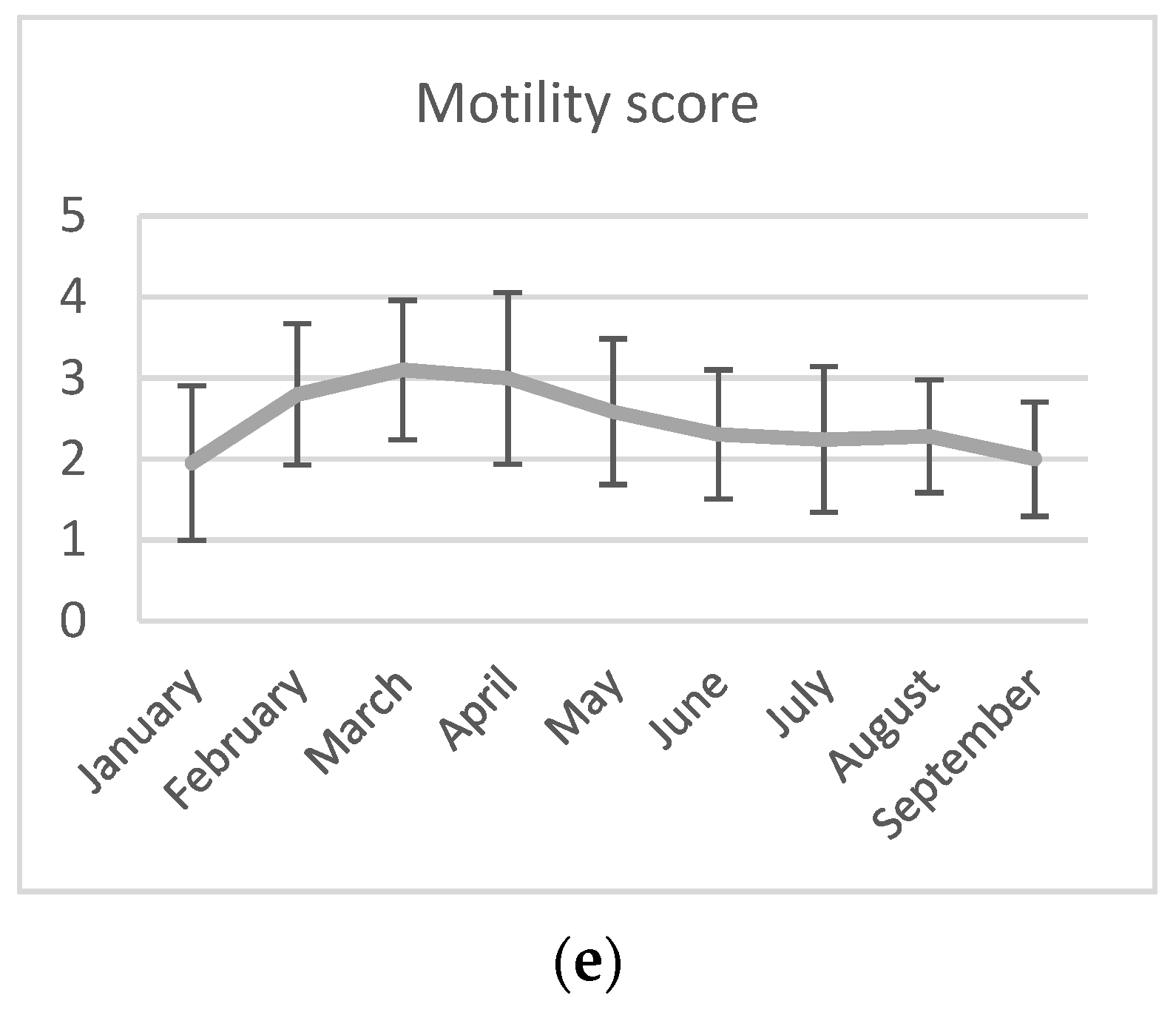
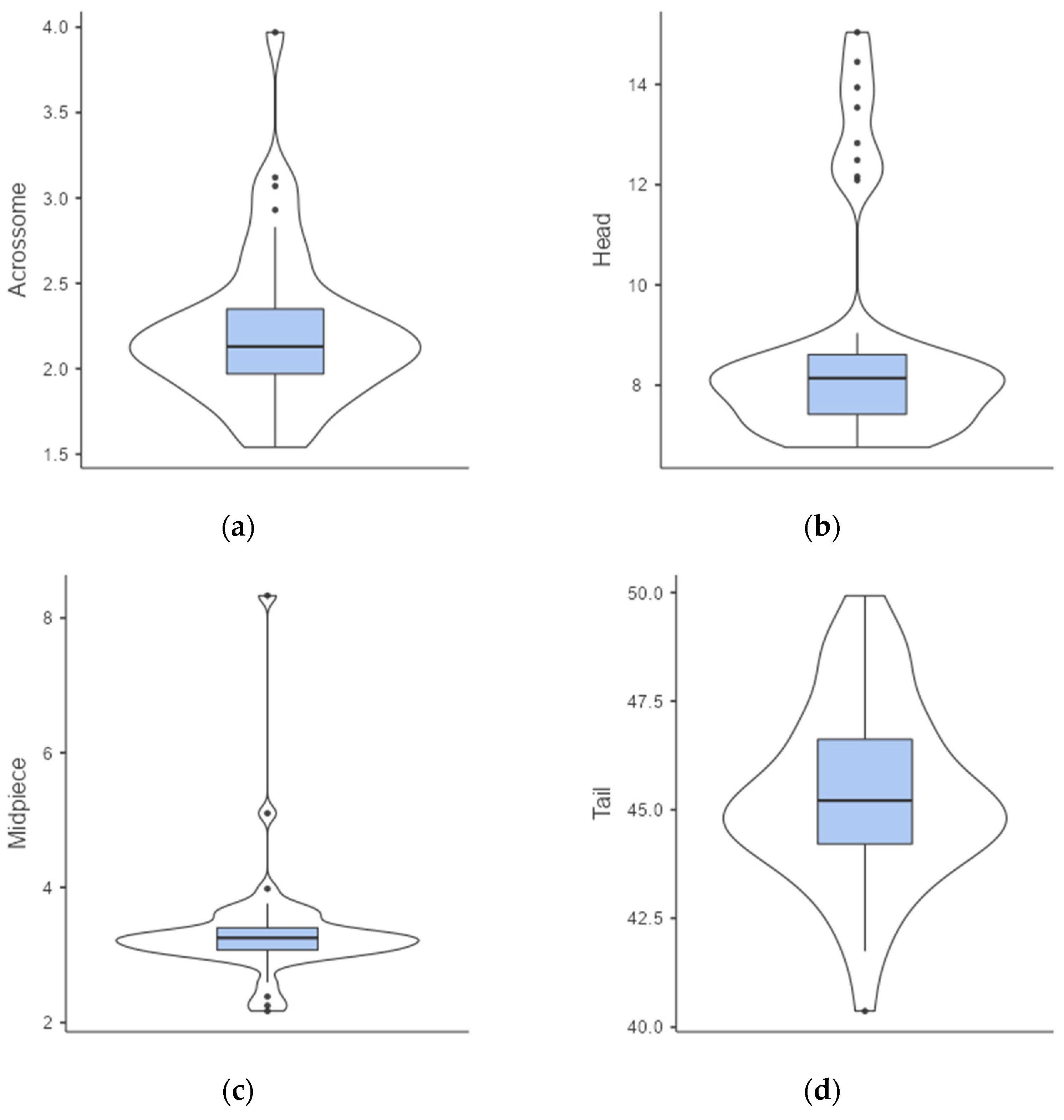
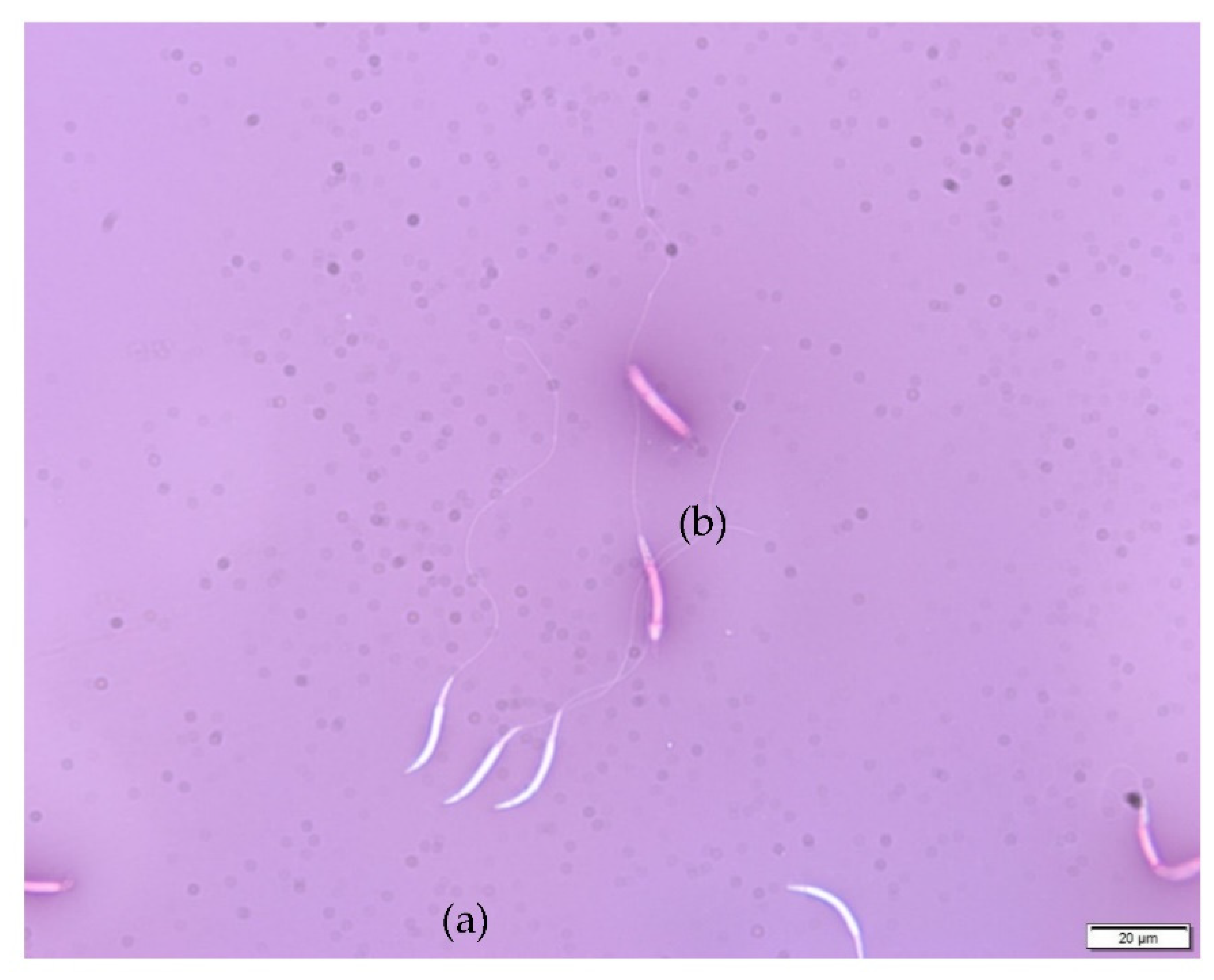
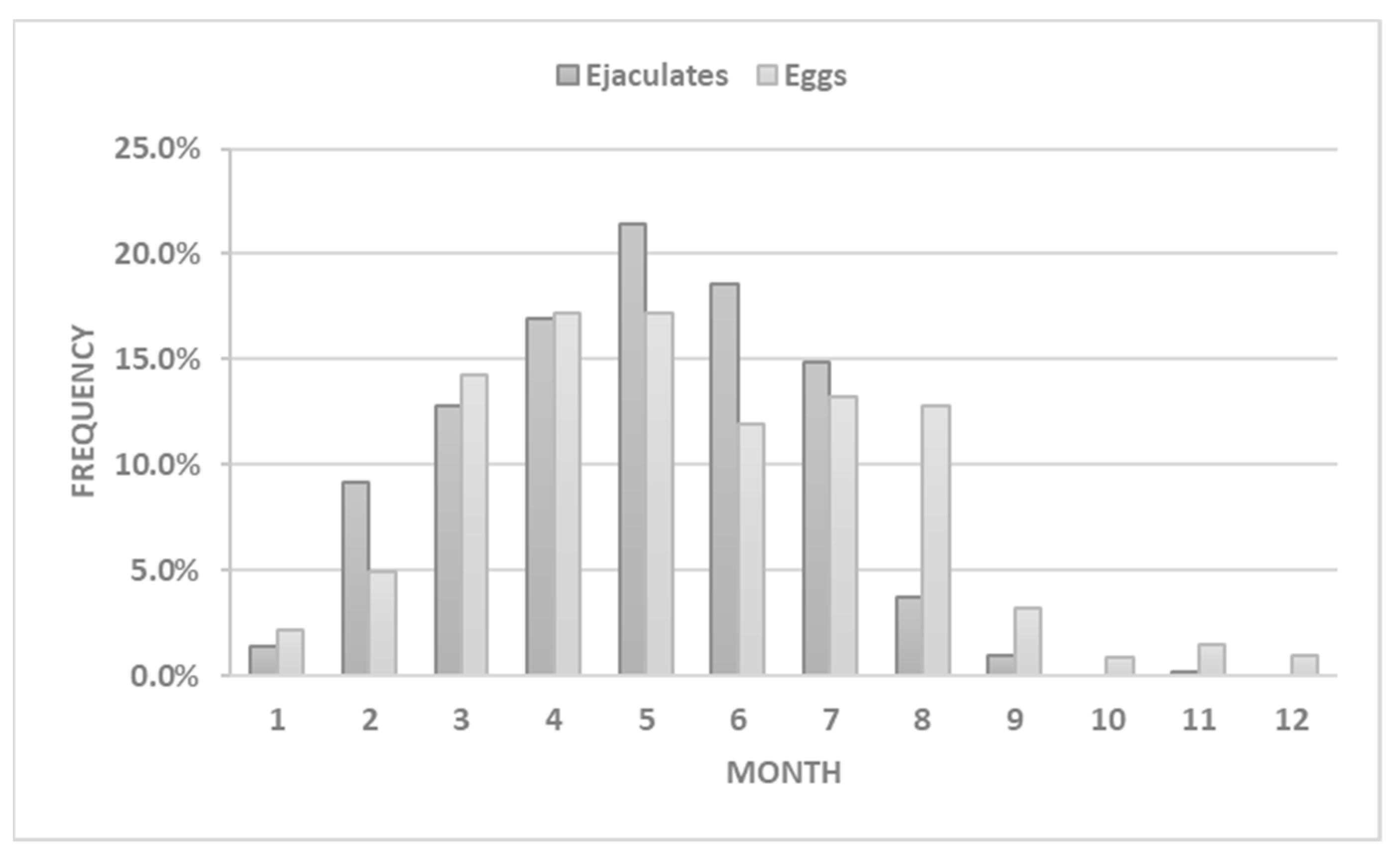
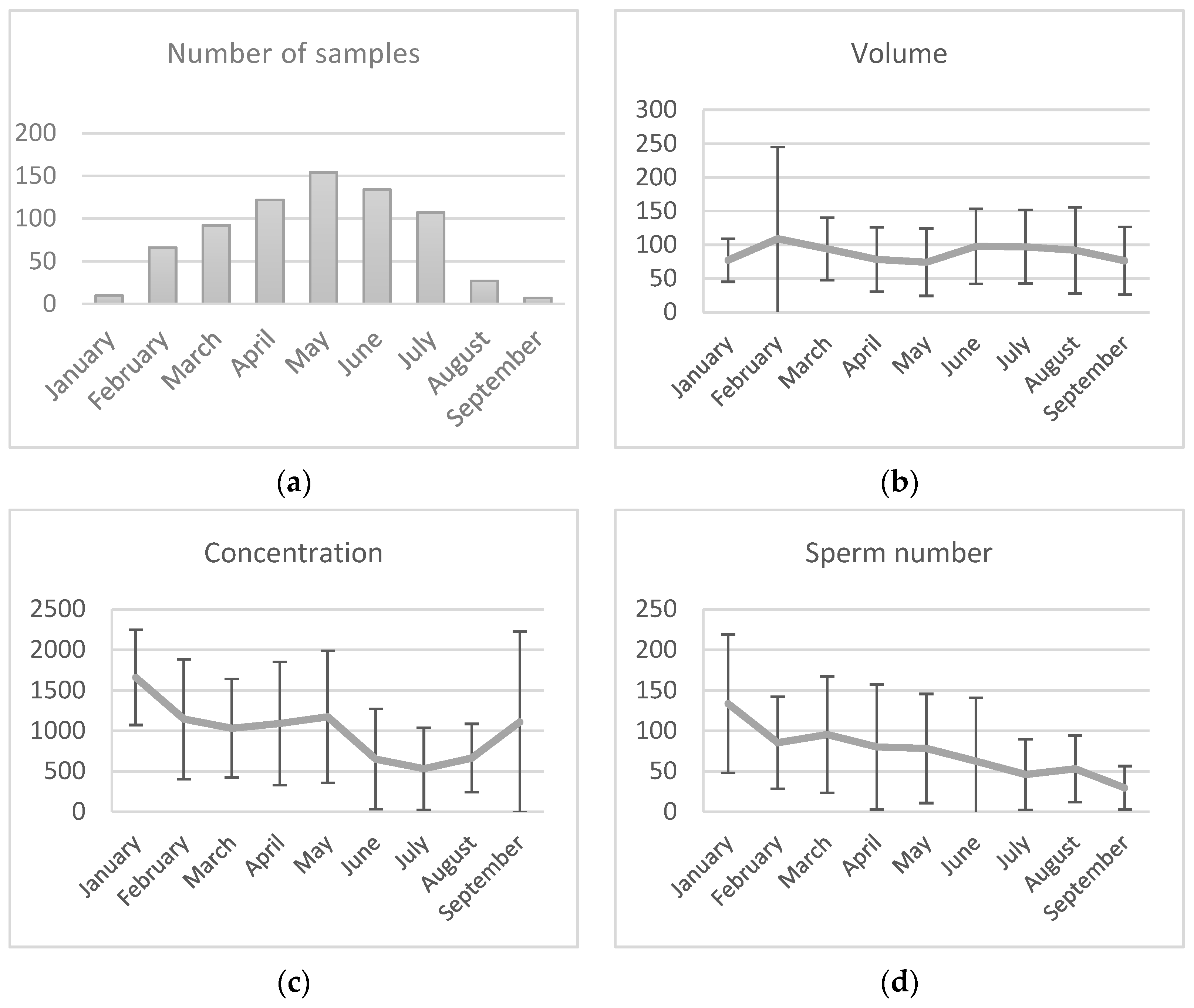
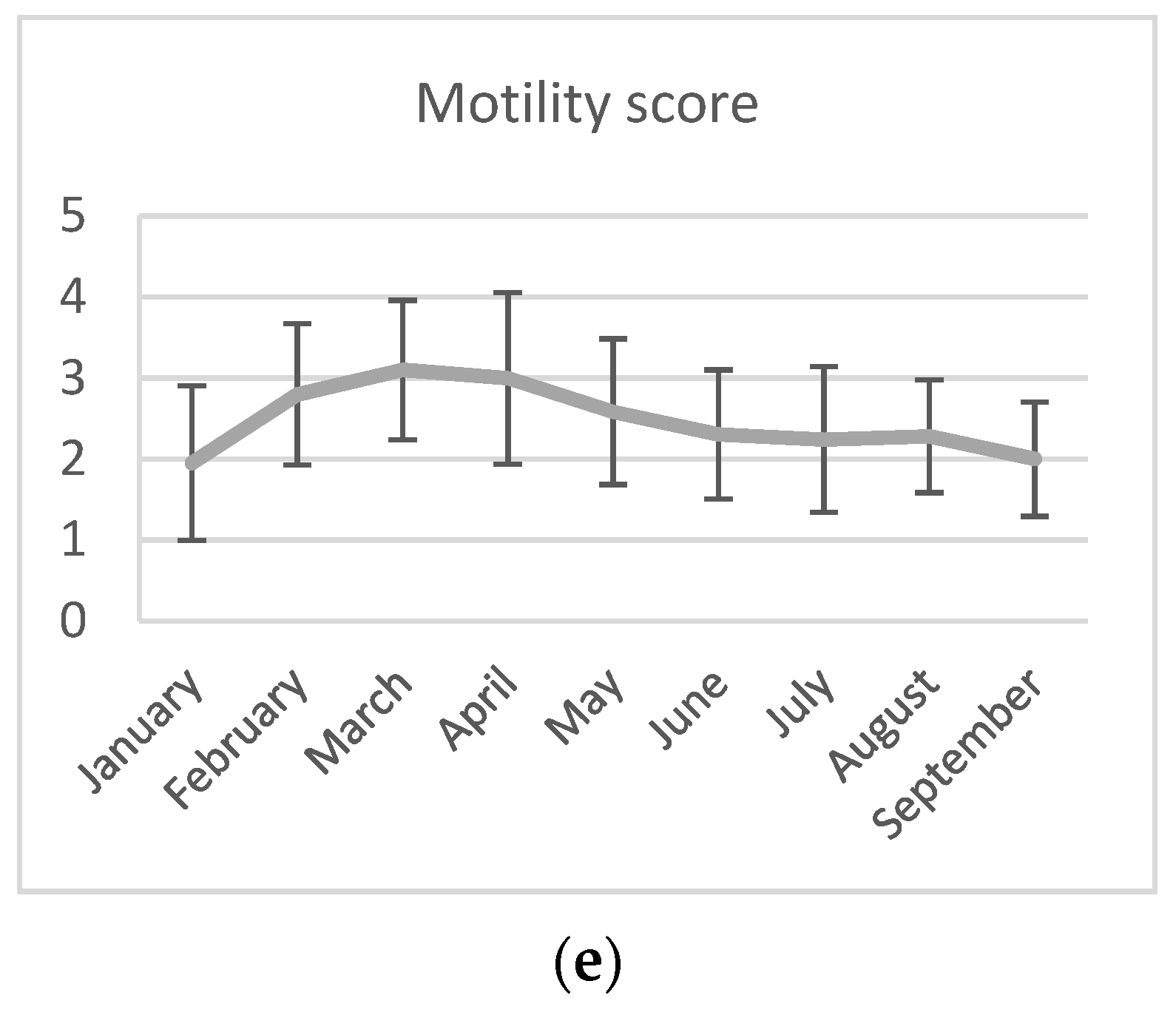
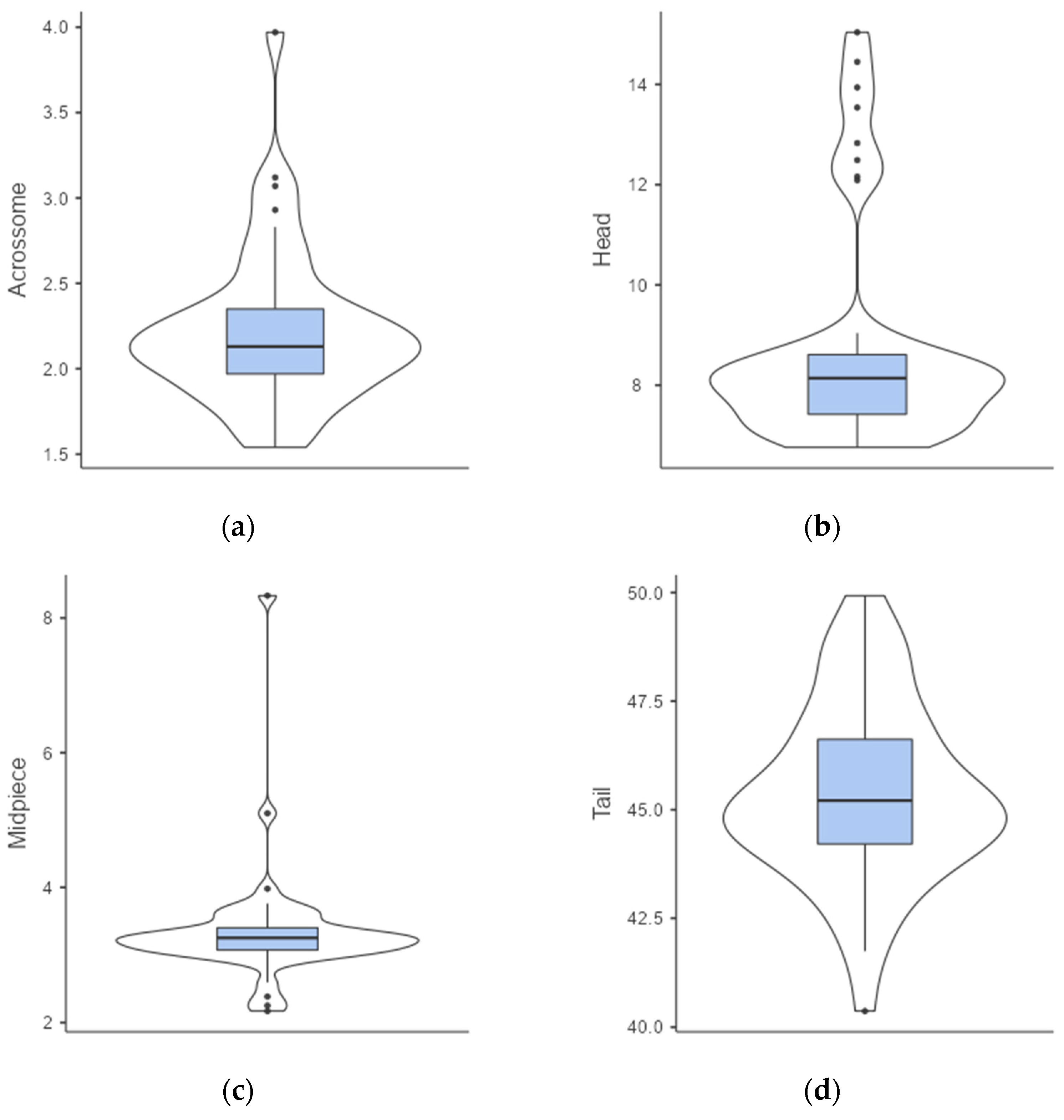
3.1. Semen Collection and Analyses
3.2. Semen Refrigeration for 24 h
3.3. Semen Cryopreservation
3.4. Artificial Insemination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Breeding Conditions










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2018 | 2019 | 2020 | 2021 | All Samples | |
|---|---|---|---|---|---|
| Number of males with collection attempts | 13 | 31 | 26 | 12 | 34 |
| Number of donors | 2 | 7 | 13 | 6 | 13 |
| Number of ejaculates | 27 | 83 | 527 | 83 | 720 |
| Number of ejaculates per donor | 13.5 | 11.9 | 40.5 | 13.8 | 25.7 |
| Non-contaminated ejaculates | 17 | 37 | 232 | 57 | 343 |
| Volume (µL) | 78.5 ± 73.1 | 55.4 ± 43.8 | 98.7 ± 53.8 | 65.5 ± 38.6 | 87.5 ± 54.1 |
| Volume range (µL) | 4–230 | 5–170 | 9–290 | 8–174 | 4–290 |
| Motility | 2.3 ± 0.8 | 2.7 ± 1.0 | 2.7 ± 0.8 | 3.7 ± 0.8 | 2.9 ± 0.9 |
| Motility range | 1.5–4 | 1–4 | 0–4 | 1.5–4.5 | 0–4.5 |
| Concentration (×106/mL) | 1204.7 ± 1070.5 | 946.7 ± 532.6 | 907.7 ± 671.8 | 1902.2 ± 863.4 | 1091.9 ± 803.6 |
| Concentration range (×106/mL) | 415.0–4103.8 | 76.6–3015.3 | 23.9–4315.3 | 194.5–3872.1 | 23.9–4315.3 |
| Sperm number (×106) | 44.3 ± 41.6 | 48.3 ± 79.3 | 78.7 ± 62.7 | 135.6 ± 124.2 | 83.2 ± 78.7 |
| Sperm number range (×106) | 8.0–150.2 | 2.8–271.4 | 5.1–335.7 | 1.6–672.8 | 1.6–672.8 |
| R14N00001 | R14N00005 | R14N00006 | R14N00009 | R15N00002 | R15N00005 | R16N00001 | R17N00010 | R18N00012 | R18N00015 | R18N00016 | R18N00017 | R18N00030 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number of collections attempts | 323 | 214 | 310 | 346 | 306 | 344 | 102 | 295 | 262 | 264 | 264 | 269 | 29 |
| Age at first collection (in months) | 61 | 56 | 47 | 55 | 40 | 57 | 46 | 16 | 22 | 22 | 10 | 21 | 19 |
| Number of ejaculates | 83 | 67 | 129 | 116 | 68 | 62 | 1 | 84 | 17 | 13 | 21 | 31 | 28 |
| Number of non-contaminated ejaculates | 43 | 20 | 53 | 56 | 36 | 31 | 0 | 40 | 9 | 9 | 15 | 18 | 13 |
| Volume (µL) | 116.4 ± 41.3 | 73.2 ± 43.3 | 104.6 ± 60.6 | 90.9 ± 54.3 | 84.3 ± 51 | 119.7 ± 70.6 | 69.5 ± 38.1 | 50.2 ± 27.6 | 41.7 ± 20.3 | 75.6 ± 44.3 | 50.3 ± 25 | 39.1 ± 22 | |
| Range Volume (min–max) | 47–224 | 24–180 | 4–237 | 6–184 | 9–210 | 9–290 | 5–174 | 17–100 | 15–73 | 15–140 | 8–121 | 12–98 | |
| Motility score (0–5) | 2.7 ± 0.8 | 2.6 ± 0.7 | 2.4 ± 0.8 | 2.9 ± 0.8 | 2.8 ± 0.7 | 2.7 ± 0.9 | 2.8 ± 1.1 | 3.6 ± 0.8 | 3.6 ± 0.7 | 3.7 ± 0.6 | 3.9 ± 0.7 | 2.6 ± 0.8 | |
| Range Motility score (min–max) | 1–3.5 | 1–3.5 | 1–4 | 1–4 | 1–4 | 0–4 | 1–4 | 2.5–4.5 | 2.5–4.5 | 3–4.5 | 2.5–4.5 | 1–3.5 | |
| Concentration (×106 spz/mL) | 835 ± 427.9 | 1113.9 ± 562.3 | 530.2 ± 559.6 | 795.6 ± 322.2 | 921.1 ± 824 | 918.1 ± 760.3 | 1633.7 ± 893.2 | 1163.7 ± 367.8 | 2257.7 ± 682.7 | 2338.5 ± 665.5 | 1319.8 ± 655.6 | 2083.4 ± 769.4 | |
| Range Concentration (min–max) | 76.6–2424.3 | 23.9–2157.4 | 67.3–3383.1 | 49.1–1674.3 | 180.9–4103.8 | 162.8–4315.3 | 307.6–3866.5 | 765–1743.5 | 1485.6–3067.7 | 1649.9–3872.1 | 194.5–3025.2 | 770.3–3618.6 | |
| Sperm number (×106 spz) | 94.9 ± 54.9 | 85.1 ± 73.9 | 40.6 ± 38.2 | 71.2 ± 51.4 | 63.8 ± 64.3 | 95.3 ± 69.3 | 125 ± 130.4 | 62.1 ± 48.1 | 89.1 ± 42.3 | 182.4 ± 123.2 | 74.2 ± 78.3 | 71.3 ± 31.8 | |
| Range Sperm number (min–max) | 10.6–233.7 | 18.8–248.4 | 5.1–179.5 | 2.8–184.2 | 7.1–271.4 | 9.5–249.4 | 6.8–672.8 | 16.5–155.1 | 40.1–156.5 | 28.9–443.2 | 1.6–366 | 41.1–162.1 |
References
- Holt, W.V.; Brown, J.L.; Comizzoli, P. Reproductive Science as an Essential Component of Conservation Biology. In Reproductive Sciences in Animal Conservation: Progress and Prospects; Holt, W.V., Brown, J.L., Comizzoli, P., Eds.; Springer: New York, NY, USA, 2014; pp. 3–14. ISBN 978-1-4939-0819-6. [Google Scholar]
- Holt, W.V.; Comizzoli, P. Opportunities and Limitations for Reproductive Science in Species Conservation. Annu. Rev. Anim. Biosci. 2021, 10, 491–511. [Google Scholar] [CrossRef] [PubMed]
- Gee, G.F.; Bertschinger, H.; Donoghue, A.M.; Blanco, J.; Soley, J. Reproduction in Nondomestic Birds: Physiology, Semen Collection, Artificial Insemination and Cryopreservation. Avian Poult. Biol. Rev. 2004, 15, 47–101. [Google Scholar] [CrossRef]
- Saint Jalme, M.; Gaucher, P.; Paillat, P. Artificial insemination in Houbara bustards (Chlamydotis undulata): Influence of the number of spermatozoa and insemination frequency on fertility and ability to hatch. J. Reprod. Fertil. 1994, 100, 93–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saint Jalme, M.; Lecoq, R.; Seigneurin, F.; Blesbois, E.; Plouzeau, E. Cryopreservation of semen from endangered pheasants: The first step towards a cryobank for endangered avian species. Theriogenology 2003, 59, 875–888. [Google Scholar] [CrossRef]
- Holt, W.V. The black-footed ferret recovery program: A strong advocate for establishing semen banking programs as support tools for small population welfare. Anim. Conserv. 2016, 19, 116–117. [Google Scholar] [CrossRef] [Green Version]
- Johnston, L.A.; Lacy, R.C. Genome resource banking for species conservation: Selection of sperm donors. Cryobiology 1995, 32, 68–77. [Google Scholar] [CrossRef]
- Rakha, B.A.; Ansari, M.S.; Akhter, S.; Hussain, I.; Blesbois, E. Cryopreservation of Indian red jungle fowl (Gallus gallus murghi) semen. Anim. Reprod. Sci. 2016, 174, 45–55. [Google Scholar] [CrossRef]
- Blesbois, E. Freezing avian semen. Avian Biol. Res. 2011, 4, 52–58. [Google Scholar] [CrossRef]
- Rabier, R.; Robert, A.; Lacroix, F.; Lesobre, L. Genetic assessment of a conservation breeding program of the Houbara bustard (Chlamydotis undulata undulata) in Morocco, based on pedigree and molecular analyses. Zoo Biol. 2020, 39, 422–435. [Google Scholar] [CrossRef]
- Holt, W.V. Crybiology, Wildlife Conservation & Reality. CryoLetters 2008, 29, 43–52. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org (accessed on 5 February 2022).
- Dutta, S.; Jhala, Y. Devil is in the detail: Behaviorally explicit habitat selection by the Critically Endangered great Indian bustard. Endanger. Species Res. 2021, 45, 55–69. [Google Scholar] [CrossRef]
- Dutta, S.; Rahmani, A.R.; Jhala, Y.V. Running out of time? The great Indian bustard Ardeotis nigriceps—Status, viability, and conservation strategies. Eur. J. Wildl. Res. 2011, 57, 615–625. [Google Scholar] [CrossRef]
- BirdLife International. Ardeotis Arabs: The IUCN Red List of Threatened Species. Available online: http://datazone.birdlife.org/species/factsheet/22691924 (accessed on 1 February 2022).
- Thiollay, J.-M. Severe decline of large birds in the Northern Sahel of West Africa: A long-term assessment. Bird Conserv. Int. 2006, 16, 353. [Google Scholar] [CrossRef] [Green Version]
- del Hoyo, J.; Elliott, A.; Sargatal, J.; Cabot, J. Handbook of the Birds of the World: Volume 3: Hoatzin to Auks; Lynx Edicions: Barcelona, Spain, 1996; p. 263. ISBN 8487334202. [Google Scholar]
- Boland, C.R.J.; Burwell, B.O. Ranking and mapping the high conservation priority bird species of Saudi Arabia. Avian Conserv. Ecol. 2020, 15, 15. [Google Scholar] [CrossRef]
- eBird. An Online Database of Bird Distribution and Abundance: eBird Basic Dataset. Version: EBD_relMar-2019. Available online: http://www.ebird.org (accessed on 20 March 2019).
- Chalah, T. Assessment of pH, Osmolarity, Motility and Viability of Houbara Bustard Semen (Chlamydotis undulata undulata). Avian Poult. Biol. Rev. 2001, 12, 176–177. [Google Scholar]
- Chalah, T. Viability, Motility and Ultra-structure of Houbara Bustard Sperm(Chlamydotis undulata undulata) Before and After the Freezing Process. Avian Poult. Biol. Rev. 2001, 12, 177–178. [Google Scholar]
- Chalah, T.; Lacroix, F. Assessment of Houbara Bustard (Chlamydotis undulata undulata) semen in captivity, stored at 4–5 °C up to 72 h, by in-vitro and in-vivo tests. In Proceedings of the 9th International Symposium on Spermatology, Cape Town, South Africa, 6–11 October 2002. [Google Scholar]
- Lacy, R.C. Managing genetic diversity in captive populations of animals. In Restoration of Endangered Species: Conceptual Issues, Planning, and Implementation; Bowles, M.L., Whelan, C.J., Eds.; Cambridge University Press: Cambridge, UK, 1994; pp. 63–89. ISBN 9780511623325. [Google Scholar]
- Ballou, J.D.; Lacy, R.C. Identifying genetically important individuals for management of genetic variation in pedigreed populations. In Population Management for Survival and Recovery: Analytical Methods and Strategies in Small Population Conservation; Ballou, J.D., Foose, T.J., Gilpin, M.E., Eds.; Columbia University Press: New York, NY, USA; Chichester, UK, 1995; pp. 76–111. ISBN 0231101775. [Google Scholar]
- Ballou, J.D.; Lees, C.M.; Faust, L.J.; Long, S.; Lynch, C.; Bingaman-Lackey, L.; Foose, T.J. Demographic and Genetic Management of Captive Populations. In Wild Mammals in Captivity: Principles and Techniques for Zoo Management, 2nd ed.; Kleiman, D.G., Baer, C.K., Thompson, K.V., Eds.; University of Chicago Press: Chicago, IL, USA; University Presses Marketing [Distributor]: Bristol, UK, 2012; pp. 219–267. ISBN 0226440117. [Google Scholar]
- Greggor, A.L.; Vicino, G.A.; Swaisgood, R.R.; Fidgett, A.; Brenner, D.; Kinney, M.E.; Farabaugh, S.; Masuda, B.; Lamberski, N. Animal Welfare in Conservation Breeding: Applications and Challenges. Front. Vet. Sci. 2018, 5, 323. [Google Scholar] [CrossRef]
- Blesbois, E.; Grasseau, I.; Seigneurin, F.; Mignon-Grasteau, S.; Saint Jalme, M.; Mialon-Richard, M.M. Predictors of success of semen cryopreservation in chickens. Theriogenology 2008, 69, 252–261. [Google Scholar] [CrossRef]
- David, I.; Kohnke, P.; Lagriffoul, G.; Praud, O.; Plouarboué, F.; Degond, P.; Druart, X. Mass sperm motility is associated with fertility in sheep. Anim. Reprod. Sci. 2015, 161, 75–81. [Google Scholar] [CrossRef]
- Lemoine, M.; Mignon-Grasteau, S.; Grasseau, I.; Magistrini, M.; Blesbois, E. Ability of chicken spermatozoa to undergo acrosome reaction after liquid storage or cryopreservation. Theriogenology 2011, 75, 122–130. [Google Scholar] [CrossRef]
- Lake, P.E.; Ravie, O. An exploration of cryoprotective compounds for fowl spermatozoa. Br. Poult. Sci. 1984, 25, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Chargé, R.; Saint Jalme, M.; Lacroix, F.; Cadet, A.; Sorci, G. Male health status, signalled by courtship display, reveals ejaculate quality and hatching success in a lekking species. J. Anim. Ecol. 2010, 79, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, C.; Staines, H.J.; McCormick, P.; McCallum, C.; Choulani, F.; Wishart, G.J. Variability in the size of the nucleus in spermatozoa from Houbara bustards, Chlamydotis undulata undulata. J. Reprod. Fertil. 1999, 117, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Hancock, J.L. The Morphology of Bull Spermatozoa. J. Exp. Biol. 1952, 29, 445–453. [Google Scholar] [CrossRef]
- Tselutin, K.; Seigneurin, F.; Blesbois, E. Comparison of cryoprotectants and methods of cryopreservation of fowl spermatozoa. Poult. Sci. 1999, 78, 586–590. [Google Scholar] [CrossRef]
- Dutta, S.; Rahmani, A.R.; Gautam, P.; Kasambe, R.; Narwade, S.; Narayan, J.; Jhala, Y.V. Guidelines for Preparation of State Action Plan for Resident Bustards’ Recovery Programme; Ministry of Environment and Forests, Government of India: New Delhi, India, 2013.
- Hallager, S. International Studbook for the Kori Bustard Ardeotis Kori; Smithsonian National Zoological Park: Washington, DC, USA, 2017. [Google Scholar]
- Ziembicki, M. Australian Bustard; CSIRO Publishing: Collingwood, Australia, 2010; ISBN 9780643096110. [Google Scholar]
- Gee, G.F. Avian artificial insemination and semen preservation. In Proceedings of the 1983 Jean Delacour/IFCB Symposium on Breeding Birds in Captivity, Los angeles, CA, USA, 24–27 February 1983; Risser, A.C., Todd, F.S., Eds.; International Foundation for the Conservation of Birds: North Hollywood, CA, USA, 1983; pp. 375–398. [Google Scholar]
- BirdLife International. Threatened Birds of Asia: The Birdlife International Red Data Book; Birdlife International: Cambridge, UK, 2001; ISBN 0946888442. [Google Scholar]
- Tsend-Ayush, E.; Dodge, N.; Mohr, J.; Casey, A.; Himmelbauer, H.; Kremitzki, C.L.; Schatzkamer, K.; Graves, T.; Warren, W.C.; Grützner, F. Higher-order genome organization in platypus and chicken sperm and repositioning of sex chromosomes during mammalian evolution. Chromosoma 2009, 118, 53–69. [Google Scholar] [CrossRef] [Green Version]
- Briskie, J.V.; Montgomerie, R.; Birkhead, T.R. The evolution of sperm size in birds. Evolution 1997, 51, 937–945. [Google Scholar] [CrossRef]
- Birkhead, T.R.; Montgomerie, R. Three decades of sperm competition in birds. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20200208. [Google Scholar] [CrossRef]
- Immler, S.; Saint Jalme, M.; Lesobre, L.; Sorci, G.; Roman, Y.; Birkhead, T.R. The evolution of sperm morphometry in pheasants. J. Evol. Biol. 2007, 20, 1008–1014. [Google Scholar] [CrossRef]
- Birkhead, T.R.; Møller, A.P. Numbers and size of sperm storage tubules and the duration of sperm storage in birds: A comparative study. Biol. J. Linn. Soc. 1992, 45, 363–372. [Google Scholar] [CrossRef]
- Briskie, J.V.; Montgomerie, R. Patterns of Sperm Storage in Relation to Sperm Competition in Passerine Birds. Condor 1993, 95, 442. [Google Scholar] [CrossRef]
- Azar, J.F.; Chalah, T.; Rautureau, P.; Lawrence, M.; Hingrat, Y. Breeding success and juvenile survival in a reintroduced captive-bred population of Asian houbara bustards in the United Arab Emirates. EndangER. Species Res. 2018, 35, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Combreau, O.; Qiao, J.; Lawrence, M.; Gao, X.; Yao, J.; Yang, W.; Launay, F. Breeding success in a Houbara Bustard Chlamydotis [undulata] macqueenii population on the eastern fringe of the Jungar Basin, People’s Republic of China. Ibis 2002, 144, E45–E56. [Google Scholar] [CrossRef]
- Raihani, G.; Székely, T.; Serrano-Meneses, M.A.; Pitra, C.; Goriup, P. The influence of sexual selection and male agility on sexual size dimorphism in bustards (Otididae). Anim. Behav. 2006, 71, 833–838. [Google Scholar] [CrossRef]
- Breed, W.G.; Bauer, M.; Wade, R.; Thitipramote, N.; Suwajarat, J.; Yelland, L. Intra-individual variation in sperm tail length in murine rodents. J. Zool. 2007, 272, 299–304. [Google Scholar] [CrossRef]
- Kleven, O.; Laskemoen, T.; Fossøy, F.; Robertson, R.J.; Lifjeld, J.T. Intraspecific variation in sperm length is negatively related to sperm competition in passerine birds. Evolution 2008, 62, 494–499. [Google Scholar] [CrossRef]
- Cramer, E.R.A.; Garcia-Del-Rey, E.; Johannessen, L.E.; Laskemoen, T.; Marthinsen, G.; Johnsen, A.; Lifjeld, J.T. Longer Sperm Swim More Slowly in the Canary Islands Chiffchaff. Cells 2021, 10, 1358. [Google Scholar] [CrossRef]
- Møller, A.P. Testes size, ejaculate quality and sperm competition in birds. Biol. J. Linn. Soc. 1988, 33, 273–283. [Google Scholar] [CrossRef]
- Vuarin, P.; Hingrat, Y.; Lesobre, L.; Jalme, M.S.; Lacroix, F.; Sorci, G. Sperm competition accentuates selection on ejaculate attributes. Biol. Lett. 2019, 15, 20180889. [Google Scholar] [CrossRef] [Green Version]
- Mizushima, S. Fertilization 2: Polyspermic Fertilization. Adv. Exp. Med. Biol. 2017, 1001, 105–123. [Google Scholar] [CrossRef]
- Hemmings, N.; Birkhead, T.R. Polyspermy in birds: Sperm numbers and embryo survival. Proc. Biol. Sci. 2015, 282, 20151682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amann, R.P.; Waberski, D. Computer-assisted sperm analysis (CASA): Capabilities and potential developments. Theriogenology 2014, 81, 5–17.e3. [Google Scholar] [CrossRef] [PubMed]
- Johnston, S.D.; López-Fernández, C.; Pappin, E.; Hampe, A.; Doneley, R.; Lierz, M.; Gosálvez, J. Assessment of avian sperm DNA fragmentation using the sperm chromatin dispersion assay. Reprod. Fertil. Dev. 2020, 32, 948–952. [Google Scholar] [CrossRef]
- Partyka, A.; Nizański, W.; Łukaszewicz, E. Evaluation of fresh and frozen-thawed fowl semen by flow cytometry. Theriogenology 2010, 74, 1019–1027. [Google Scholar] [CrossRef]
- Chalah, T.; Brillard, J.P. Comparison of assessment of fowl sperm viability by eosin-nigrosin and dual fluorescence (SYBR-14/PI). Theriogenology 1998, 50, 487–493. [Google Scholar] [CrossRef]
- Slowinska, M.; Liszewska, E.; Judycka, S.; Konopka, M.; Ciereszko, A. Mitochondrial membrane potential and reactive oxygen species in liquid stored and cryopreserved turkey (Meleagris gallopavo) spermatozoa. Poultry Science 2018, 97, 3709–3717. [Google Scholar] [CrossRef] [PubMed]
- Rakha, B.A.; Ansari, M.S.; Hussain, I.; Anwar, M.; Akhter, S.; Blesbois, E. Comparison of extenders for liquid storage of Indian Red Jungle Fowl (Gallus gallus murghi) spermatozoa. Avian Biol Res 2016, 9, 207–212. [Google Scholar] [CrossRef]

| Mean | Range | |
|---|---|---|
| Number of males with collection attempts | 34 | |
| Number of donors | 13 | |
| Age at first collection (in months) | 36.3 ± 18.4 | 10–61 |
| Number of ejaculates | 720 | |
| Number of ejaculates per donor | 55.4 ± 40.6 | 1–129 |
| Number of ejaculates per donor and per year | 25.7 ± 26.7 | 1–84 |
| Non-contaminated ejaculates | 343 | |
| Volume (µL) | 87.5 ± 54.1 | 4–290 |
| Motility | 2.9 ± 0.9 | 0–4.5 |
| Concentration (106/mL) | 1091.9 ± 803.6 | 23.9–4315.3 |
| Sperm number (×106) | 83.18 ± 78.7 | 1.6–672.8 |
| Fresh | Frozen/Thawed | p-Value | ||
|---|---|---|---|---|
| Normal live | Mean | 90.3 ± 8.2 | 52.9 ± 11 | |
| Range (min/max) | 76.5–99.9 | 31–71 | p ≤ 0.001 | |
| Normal dead | Mean | 3.4 ± 2.4 | 38.6 ± 10.2 | |
| Range (min/max) | 0–8 | 23–57 | p ≤ 0.001 | |
| Abnormal live | Mean | 8.3 ± 5 | 4.2 ± 3.5 | |
| Range (min/max) | 3–17 | 0–11 | p ≤ 0.05 | |
| Abnormal dead | Mean | 1.2 ± 1.5 | 4.2 ± 3.7 | |
| Range (min/max) | 0–4.2 | 0–10 | p ≤ 0.01 |
| Fresh | 24 h Refrigerated | Cryopreserved | |
|---|---|---|---|
| Artificial Inseminations | 41 | 5 | 19 |
| Volume (µL) | 209.4 ± 62.0 µL | 211.0 ± 72.5 | 183.6 ± 72.5 |
| Number of sperm (×106) | 73.6 ± 34.9 | 78.3 ± 21.8 | 66.6 ± 33.5 |
| Motility | 3.3 ± 0.7 | 3.4 ± 0.5 | 1.9 ± 0.4 |
| Eggs | 27 | 0 | 6 |
| Fertile Eggs (9th day of incubation) | 23 | 0 | 5 |
| Fertility rate | 85.2% | 83.3% |
| Arabian Bustard (This Study) | North African Houbara | Asian Houbara | |
|---|---|---|---|
| Ejaculates | 719 | 852,714 | 397,877 |
| Volume (µL) | 89.2 | 82.1 | 63.1 |
| Motility | 2.6 | 2.9 | 2.8 |
| Concentration | 928.7 | 401.4 | 423.5 |
| Number of sperm | 72.5 | 31.1 | 26.3 |
| Proportion of normal sperm | 0.89 | 0.80 [53] | n/a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carreira, J.T.; Lesobre, L.; Boullenger, S.; Chalah, T.; Lacroix, F.; Hingrat, Y. Assisted Reproduction Techniques to Improve Reproduction in a Non-Model Species: The Case of the Arabian Bustard (Ardeotis arabs) Conservation Breeding Program. Animals 2022, 12, 851. https://doi.org/10.3390/ani12070851
Carreira JT, Lesobre L, Boullenger S, Chalah T, Lacroix F, Hingrat Y. Assisted Reproduction Techniques to Improve Reproduction in a Non-Model Species: The Case of the Arabian Bustard (Ardeotis arabs) Conservation Breeding Program. Animals. 2022; 12(7):851. https://doi.org/10.3390/ani12070851
Chicago/Turabian StyleCarreira, Janaina Torres, Loïc Lesobre, Sylvain Boullenger, Toni Chalah, Frédéric Lacroix, and Yves Hingrat. 2022. "Assisted Reproduction Techniques to Improve Reproduction in a Non-Model Species: The Case of the Arabian Bustard (Ardeotis arabs) Conservation Breeding Program" Animals 12, no. 7: 851. https://doi.org/10.3390/ani12070851
APA StyleCarreira, J. T., Lesobre, L., Boullenger, S., Chalah, T., Lacroix, F., & Hingrat, Y. (2022). Assisted Reproduction Techniques to Improve Reproduction in a Non-Model Species: The Case of the Arabian Bustard (Ardeotis arabs) Conservation Breeding Program. Animals, 12(7), 851. https://doi.org/10.3390/ani12070851