
Detection of a Novel Chlamydia Species in Invasive Turtles

 ,
,  ,
,  ,
,  and
and 
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Molecular Analysis
2.2.1. Nucleic Acid Extraction
2.2.2. Real-Time PCR
2.2.3. Species Designation
2.3. Phylogenetic Analysis
3. Results
3.1. Sampling
3.2. Molecular Analysis
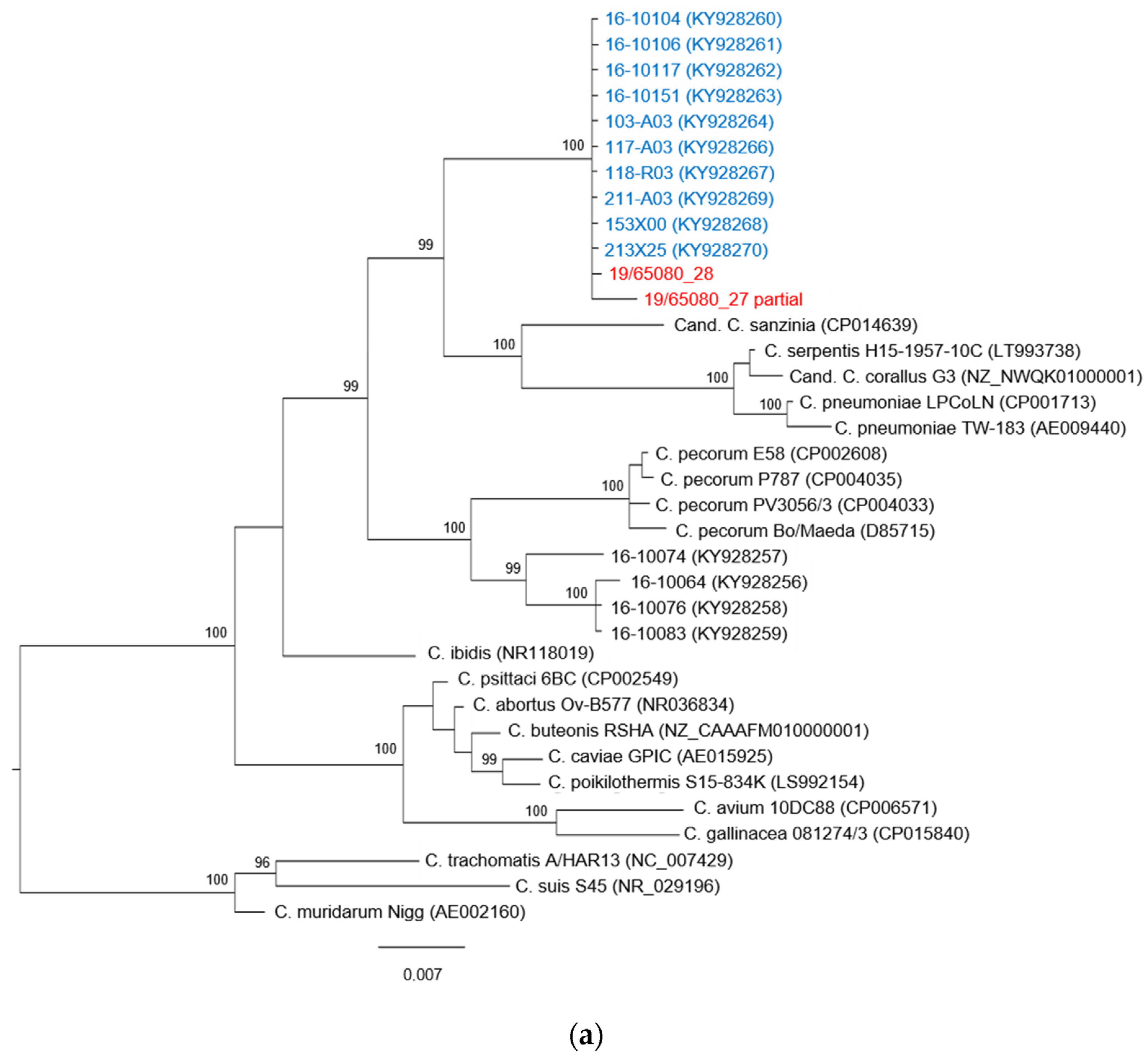
3.3. Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Warwick, C. Red-eared terrapin farms and conservation. Oryx 1986, 20, 37–40. [Google Scholar] [CrossRef]
- Yasukawa, Y. Alien freshwater turtles which may become established in Japan in the near future. Bull. Herpetol. Soc. Jpn. 2005, 2, 155–163. [Google Scholar]
- Warwick, C. Conservation of red-eared terrapins Trachemys scripta elegans: Threats from international pet and culinary markets. BCG Testudo 1991, 3, 34–44. [Google Scholar]
- Pleguezuelos, J.M. Las especies introducidas de Amphibios y Reptiles. In Atlas y Libro Rojo de los Anfibios y Reptiles de España; Organismo Autónomo de Parques Nacionales Gran Vía de San Francisco: Madrid, Spain, 2002; pp. 501–532. [Google Scholar]
- Cadi, A.; Teillac, P.; Delmas, V.; Girondot, M.; Servais, V.; Prevot-Julliard, A. Slider turtles (Trachemys scripta elegans) released in France: A case of integrated research and conservation program. Rev. Española Herpetol. 2008, 22, 111–114. [Google Scholar]
- Cadi, A.; Delmas, V.; Prévot-Julliard, A.-C.; Joly, P.; Pieau, C.; Girondot, M. Successful reproduction of the introduced slider turtle (Trachemys scripta elegans) in the South of France. Aquat. Conserv. Mar. Freshw. Ecosyst. 2004, 14, 237–246. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Thuiller, W.; Padoa-Schioppa, E. From introduction to the establishment of alien species: Bioclimatic differences between presence and reproduction localities in the slider turtle. Divers. Distrib. 2009, 15, 108–116. [Google Scholar] [CrossRef]
- Ferri, V.; Soccini, C. Case Study 11. Management of Abandoned North American Pond Turtles (Trachemys scripta) in Italy. In Urban Herpetology; Society for the Study of Amphibians and Reptiles: Salt Lake Cit, UT, USA, 2008; pp. 529–534. [Google Scholar]
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien species as a driver of recent extinctions. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; The Invasive Species Specialist Group (ISSG) a Specialist Group of the Species Survival Commission (SSC) of the World Conservation Union (IUCN)—Hollands Printing Ltd.: Auckland, New Zealand, 2000; pp. 1–12. [Google Scholar]
- Cadi, A.; Joly, P. Competition for basking places between the endangered European pond turtle (Emys orbicularis galloitalica) and the introduced red-eared slider (Trachemys scripta elegans). Can. J. Zool. 2003, 81, 1392–1398. [Google Scholar] [CrossRef] [Green Version]
- Prévot-Julliard, A.-C.; Gousset, E.; Archinard, C.; Cadi, A.; Girondot, M. Pets and invasion risks: Is the Slider turtle strictly carnivorous? Amphibia-Reptilia 2007, 28, 139–143. [Google Scholar] [CrossRef]
- Lindsay, M.K.; Zhang, Y.; Forstner, M.R.J.; Hahn, D. Effects of the freshwater turtle Trachemys scripta elegans on ecosystem functioning: An approach in experimental ponds. Amphibia-Reptilia 2013, 34, 75–84. [Google Scholar] [CrossRef]
- European Parliament. Regulation (EU) No 1143/2014 of the European Parliament and the Council of 22 October 2014 on the Prevention and Management of the Introduction and Spread of Invasive Alien Species; European Parliament: Strasbourg, France, 2014; Volume 317, pp. 35–55. [Google Scholar]
- European Commission. Commission Implementing Regulation (EU) 2016/1141 of 13 July 2016 adopting a list of invasive alien species of Union concern pursuant to Regulation (EU) No 1143/2014 of the European Parliament and of the Council. Off. J. Eur. Union 2016, 189, 4–8. [Google Scholar]
- Roy, H.E.; Hesketh, H.; Purse, B.V.; Eilenberg, J.; Santini, A.; Scalera, R.; Stentiford, G.D.; Adriaens, T.; Bacela-Spychalska, K.; Bass, D.; et al. Alien Pathogens on the Horizon: Opportunities for Predicting their Threat to Wildlife. Conserv. Lett. 2017, 10, 476–483. [Google Scholar] [CrossRef]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Anthropogenic environmental change and the emergence of infectious diseases in wildlife. Acta Trop. 2001, 78, 103–116. [Google Scholar] [CrossRef]
- Oi, M.; Araki, J.; Matsumoto, J.; Nogami, S. Helminth fauna of a turtle species introduced in Japan, the red-eared slider turtle (Trachemys scripta elegans). Res. Vet. Sci. 2012, 93, 826–830. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Silvestre, A.; Guinea, D.; Ferrer, D.; Pantchev, N. Parasitic Enteritis Associated with the Camallanid Nematode Serpinema microcephalus in Wild Invasive Turtles (Trachemys, Pseudemys, Graptemys, and Ocadia) in Spain. J. Herpetol. Med. Surg. 2015, 25, 48. [Google Scholar] [CrossRef]
- Martínez-Ríos, M.; Martín-Torrijos, L.; Diéguez-Uribeondo, J. The invasive alien red-eared slider turtle, Trachemys scripta, as a carrier of STEF-disease pathogens. Fungal Biol. 2022, 126, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Kolesnik, E.; Obiegala, A.; Marschang, R.E. Detection of Mycoplasma spp., herpesviruses, topiviruses, and ferlaviruses in samples from chelonians in Europe. J. Vet. Diagn. Investig. 2017, 29, 820–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebani, V.V.; Fratini, F.; Bertelloni, F.; Cerri, D.; Tortoli, E. Isolation and identification of mycobacteria from captive reptiles. Res. Vet. Sci. 2012, 93, 1136–1138. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo-Vila, J.; Díaz-Paniagua, C.; Pérez-Santigosa, N.; de Frutos-Escobar, C.; Herrero-Herrero, A. Salmonella in free-living exotic and native turtles and in pet exotic turtles from SW Spain. Res. Vet. Sci. 2008, 85, 449–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, J.R.; Neil, K.P.; Behravesh, C.B.; Sotir, M.J.; Angulo, F.J. Recent multistate outbreaks of human Salmonella infections acquired from turtles: A continuing public health challenge. Clin. Infect. Dis. 2010, 50, 554–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gioia-Di Chiacchio, R.; Penido Júnior, G.N.; De Souza, C.A.I.; Prioste, F.E.S.; Prado, M.S.; Knöbl, T.; Menão, M.C.; Matushima, E.R. Enterobacterial colonization in captive red-eared sliders (Trachemys scripta elegans). J. Zoo Wildl. Med. 2014, 45, 919–921. [Google Scholar] [CrossRef] [PubMed]
- Hotzel, H.; Blahak, S.; Diller, R.; Sachse, K. Evidence of infection in tortoises by Chlamydia-like organisms that are genetically distinct from known Chlamydiaceae species. Vet. Res. Commun. 2005, 29, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Mitura, A.; Niemczuk, K.; Zaręba, K.; Zając, M.; Laroucau, K.; Szymańska-Czerwińska, M. Free-living and captive turtles and tortoises as carriers of new Chlamydia spp. PLoS ONE 2017, 12, e0185407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laroucau, K.; Ortega, N.; Vorimore, F.; Aaziz, R.; Mitura, A.; Szymanska-Czerwinska, M.; Cicerol, M.; Salinas, J.; Sachse, K.; Caro, M.R. Detection of a novel Chlamydia species in captive spur-thighed tortoises (Testudo graeca) in southeastern Spain and proposal of Candidatus Chlamydia testudinis. Syst. Appl. Microbiol. 2020, 43, 126071. [Google Scholar] [CrossRef] [PubMed]
- Zarȩba-Marchewka, K.; Szymańska-Czerwińska, M.; Niemczuk, K. Chlamydiae—What’s new? J. Vet. Res. 2020, 64, 461. [Google Scholar] [CrossRef] [PubMed]
- Vorimore, F.; Hölzer, M.; Liebler-Tenorio, E.M.; Barf, L.M.; Delannoy, S.; Vittecoq, M.; Wedlarski, R.; Lécu, A.; Scharf, S.; Blanchard, Y.; et al. Evidence for the existence of a new genus Chlamydiifrater gen. nov. inside the family Chlamydiaceae with two new species isolated from flamingo (Phoenicopterus roseus): Chlamydiifrater phoenicopteri sp. nov. and Chlamydiifrater volucris sp. nov. Syst. Appl. Microbiol. 2021, 44, 126200. [Google Scholar] [CrossRef] [PubMed]
- Borel, N.; Greub, G. International Committee on Systematics of Prokaryotes (ICSP) Subcommittee on the taxonomy of Chlamydiae, minutes of the closed meeting, 10 September 2020, via Zoom. Int. J. Syst. Evol. Microbiol. 2021, 71, 004620. [Google Scholar] [CrossRef] [PubMed]
- Staub, E.; Marti, H.; Biondi, R.; Levi, A.; Donati, M.; Leonard, C.A.; Ley, S.D.; Pillonel, T.; Greub, G.; Seth-Smith, H.M.B.; et al. Novel Chlamydia species isolated from snakes are temperature-sensitive and exhibit decreased susceptibility to azithromycin. Sci. Rep. 2018, 8, 5660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huchzermeyer, F.W.; Gerdes, G.H.; Foggin, C.M.; Huchzermeyer, K.D.; Limper, L.C. Hepatitis in farmed hatchling Nile crocodiles (Crocodylus niloticus) due to chlamydial infection. J. S. Afr. Vet. Assoc. 1994, 65, 20–22. [Google Scholar] [PubMed]
- Homer, B.L.; Jacobson, E.R.; Schumacher, J.; Scherba, G. Chlamydiosis in mariculture-reared green sea turtles (Chelonia mydas). Vet. Pathol. 1994, 31, 1–7. [Google Scholar] [CrossRef]
- Bodetti, T.J.; Jacobson, E.; Wan, C.; Hafner, L.; Pospischil, A.; Rose, K.; Timms, P. Molecular Evidence to Support the Expansion of the Hostrange of Chlamydophila pneumoniae to Include Reptiles as Well as Humans, Horses, Koalas and Amphibians. Syst. Appl. Microbiol. 2002, 25, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Brown, A.; Rüegg, S.; Polkinghorne, A.; Borel, N. Characterisation of Chlamydia pneumoniae and other novel chlamydial infections in captive snakes. Vet. Microbiol. 2015, 178, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Brown, A.; Spang, L.; Borel, N.; Polkinghorne, A. Culture-independent metagenomics supports discovery of uncultivable bacteria within the genus Chlamydia. Sci. Rep. 2017, 7, 10661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor-Brown, A.; Bachmann, N.L.; Borel, N.; Polkinghorne, A. Culture-independent genomic characterisation of Candidatus Chlamydia sanzinia, a novel uncultivated bacterium infecting snakes. BMC Genom. 2016, 17, 710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebani, V.V. Domestic reptiles as source of zoonotic bacteria: A mini review. Asian Pac. J. Trop. Med. 2017, 10, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Laroucau, K.; Aaziz, R.; Lécu, A.; Laidebeure, S.; Marquis, O.; Vorimore, F.; Thierry, S.; Briend-Marchal, A.; Miclard, J.; Izembart, A.; et al. A cluster of Chlamydia serpentis cases in captive snakes. Vet. Microbiol. 2020, 240, 108499. [Google Scholar] [CrossRef] [PubMed]
- Ehricht, R.; Slickers, P.; Goellner, S.; Hotzel, H.; Sachse, K. Optimized DNA microarray assay allows detection and genotyping of single PCR-amplifiable target copies. Mol. Cell. Probes 2006, 20, 60–63. [Google Scholar] [CrossRef]
- OIE Section 3.3. Aves—Chapter 3.3.1 Avian Chlamydiosis. OIE Terrestrial Manual. 2018, pp. 783–795. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.03.01_AVIAN_CHLAMYD.pdf (accessed on 16 March 2022).
- Vicari, N.; Santoni, R.; Vigo, P.; Magnino, S. A PCR-RFLP Assay Targeting the 16S Ribosomal Gene for the Diagnosis of Animal Chlamydioses. In Proceedings of the 5th Meeting of the European Society for Chlamydia Research, Budapest, Hungary, 1–4 September 2004; p. 297. [Google Scholar]
- Denamur, E.; Sayada, C.; Souriau, A.; Orfila, J.; Rodolakis, A.; Elion, J. Restriction pattern of the major outer-membrane protein gene provides evidence for a homogeneous invasive group among ruminant isolates of Chlamydia psittaci. J. Gen. Microbiol. 1991, 137, 2525–2530. [Google Scholar] [CrossRef] [Green Version]
- Laroucau, K.; Vorimore, F.; Aaziz, R.; Berndt, A.; Schubert, E.; Sachse, K. Isolation of a new chlamydial agent from infected domestic poultry coincided with cases of atypical pneumonia among slaughterhouse workers in France. Infect. Genet. Evol. 2009, 9, 1240–1247. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huchzermeyer, F.W.; Langelet, E.; Putterill, J.F. An outbreak of chlamydiosis in farmed Indopacific crocodiles (Crocodylus porosus). J. S. Afr. Vet. Assoc. 2008, 79, 99–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabeya, H.; Sato, S.; Maruyama, S. Prevalence and characterization of Chlamydia DNA in zoo animals in Japan. Microbiol. Immunol. 2015, 59, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, L.M.; Vogel, L.A.; Bowden, R.M. Understanding the vertebrate immune system: Insights from the reptilian perspective. J. Exp. Biol. 2010, 213, 661–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmerman, L.M.; Clairardin, S.G.; Paitz, R.T.; Hicke, J.W.; LaMagdeleine, K.A.; Vogel, L.A.; Bowden, R.M. Humoral immune responses are maintained with age in a long-lived ectotherm, the red-eared slider turtle. J. Exp. Biol. 2013, 216, 633–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagano, N.; Oana, S.; Nagano, Y.; Arakawa, Y. A severe Salmonella enterica serotype Paratyphi B infection in a child related to a pet turtle, Trachemys scripta elegans. Jpn. J. Infect. Dis. 2006, 59, 132–134. [Google Scholar] [PubMed]
- Pace, A.; Vicari, N.; Rigamonti, S.; Magnino, S.; Borrelli, L.; Dipineto, L.; Fioretti, A.; Hochscheid, S.; Tavares, L.; Duarte, A. Detection of Chlamydial DNA from Mediterranean Loggerhead Sea Turtles in Southern Italy. Animals 2022, 12, 715. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellinati, L.; Pesaro, S.; Marcer, F.; Danesi, P.; Natale, A.; Ceglie, L. Detection of a Novel Chlamydia Species in Invasive Turtles. Animals 2022, 12, 784. https://doi.org/10.3390/ani12060784
Bellinati L, Pesaro S, Marcer F, Danesi P, Natale A, Ceglie L. Detection of a Novel Chlamydia Species in Invasive Turtles. Animals. 2022; 12(6):784. https://doi.org/10.3390/ani12060784
Chicago/Turabian StyleBellinati, Laura, Stefano Pesaro, Federica Marcer, Patrizia Danesi, Alda Natale, and Letizia Ceglie. 2022. "Detection of a Novel Chlamydia Species in Invasive Turtles" Animals 12, no. 6: 784. https://doi.org/10.3390/ani12060784
APA StyleBellinati, L., Pesaro, S., Marcer, F., Danesi, P., Natale, A., & Ceglie, L. (2022). Detection of a Novel Chlamydia Species in Invasive Turtles. Animals, 12(6), 784. https://doi.org/10.3390/ani12060784