Integration of Long Non-Coding RNA and mRNA Profiling Reveals the Mechanisms of Different Dietary NFC/NDF Ratios Induced Rumen Development in Calves

 ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Feed Intake and Growth Performance
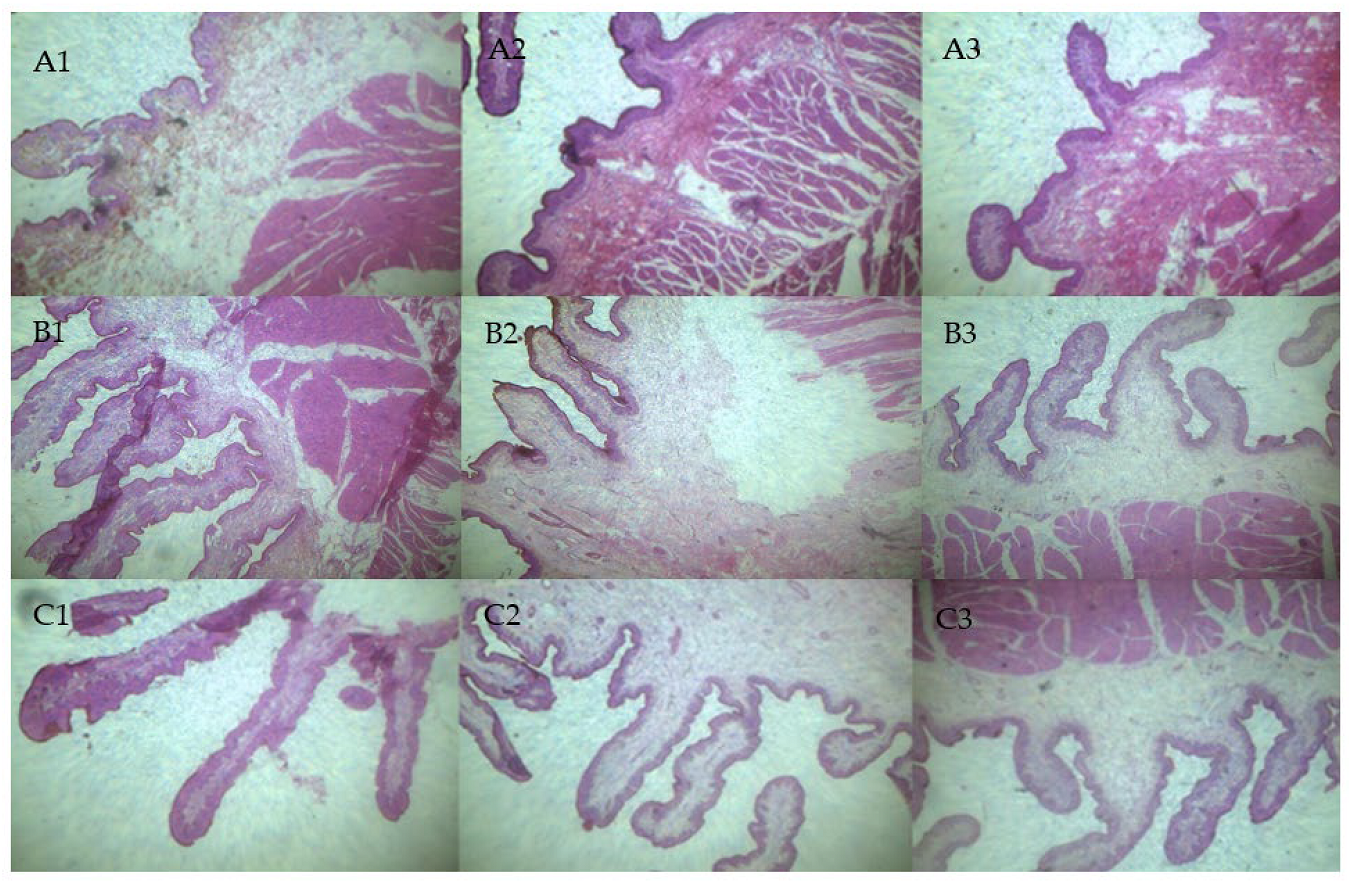
2.3. Preparation and Observation of Rumen Sections
2.4. RNA Extraction, Library Construction, and Sequencing
2.5. Data Mapping and Transcriptome Assembly
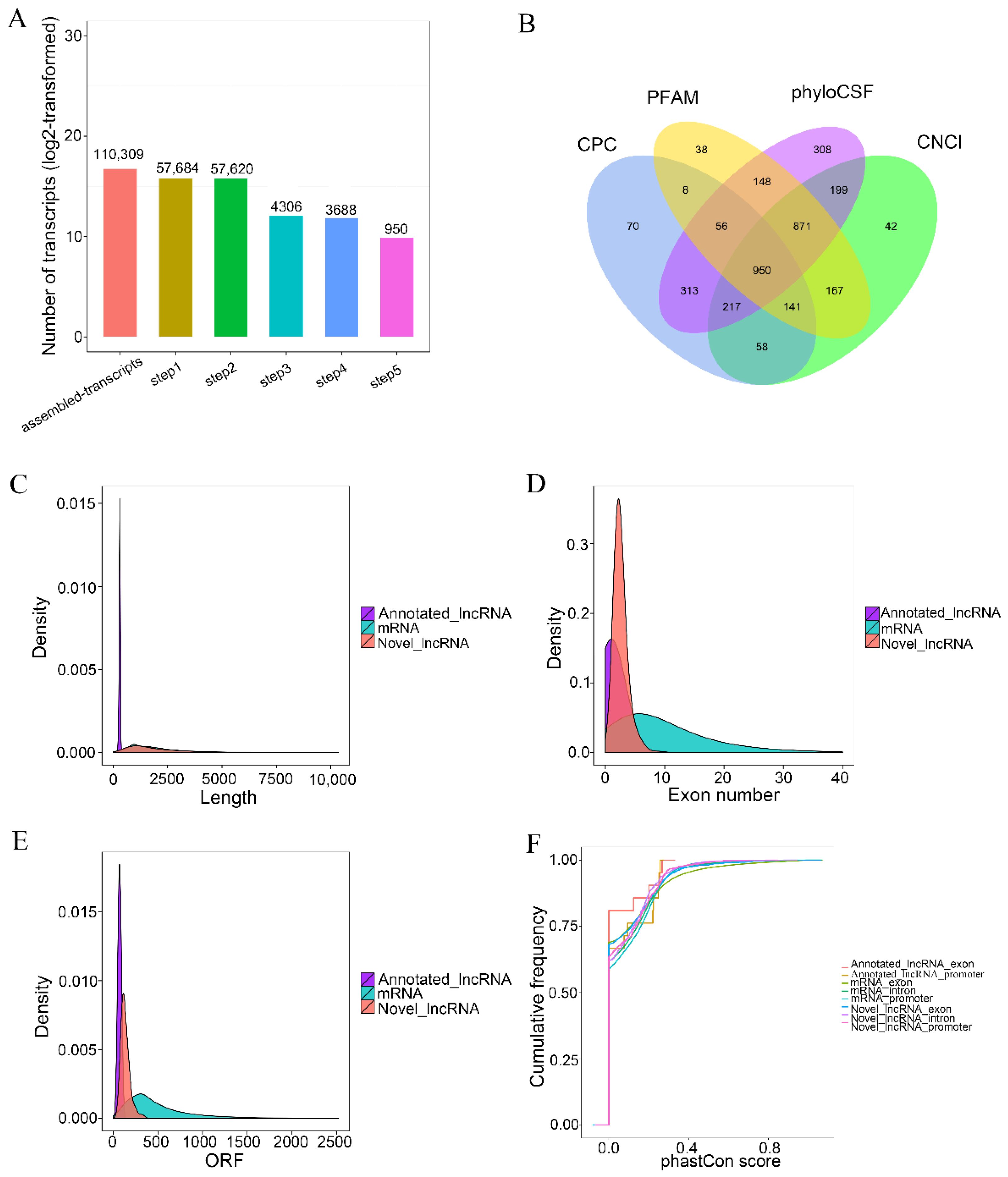
2.6. LncRNA Prediction
2.7. Structural and Conservative Analysis
2.8. Quantitation of Gene Expression
2.9. Prediction of lncRNA Targets
2.10. Gene Ontology and Kyoto Encyclopedia of Genes and Genomes Analysis
2.11. Quantitative Real-Time PCR Analysis
2.12. Statistical Analysis
3. Results
3.1. Feed Intake and Growth Performance
3.2. Rumen Fermentation Parameters and Morphological Observation of Rumen Tissue
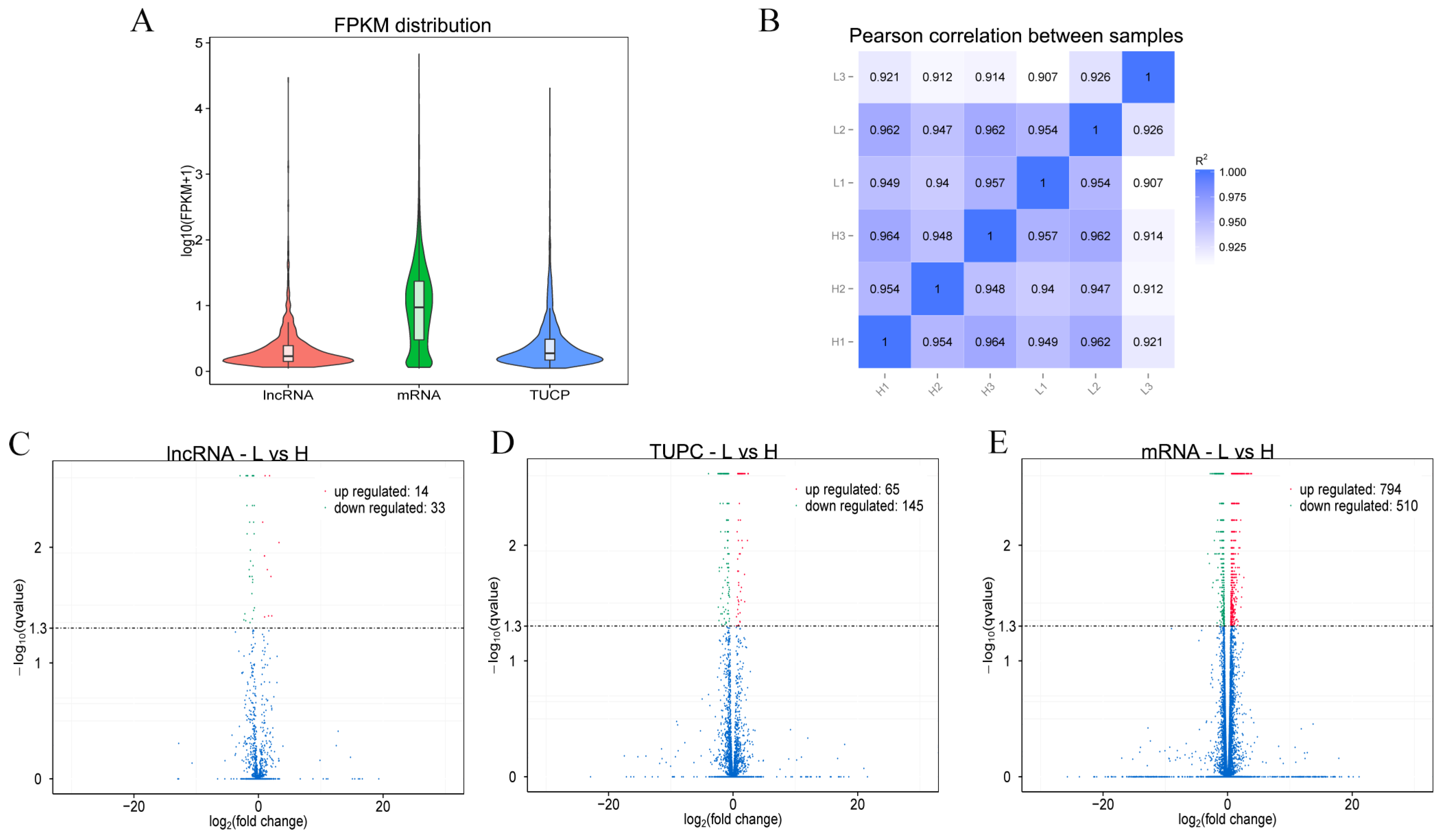
3.3. High-Throughput Sequencing and Quality Control
3.4. Identification of lncRNAs and mRNAs in Calves Rumen
3.5. Differential Expression Analysis of lncRNAs and mRNAs
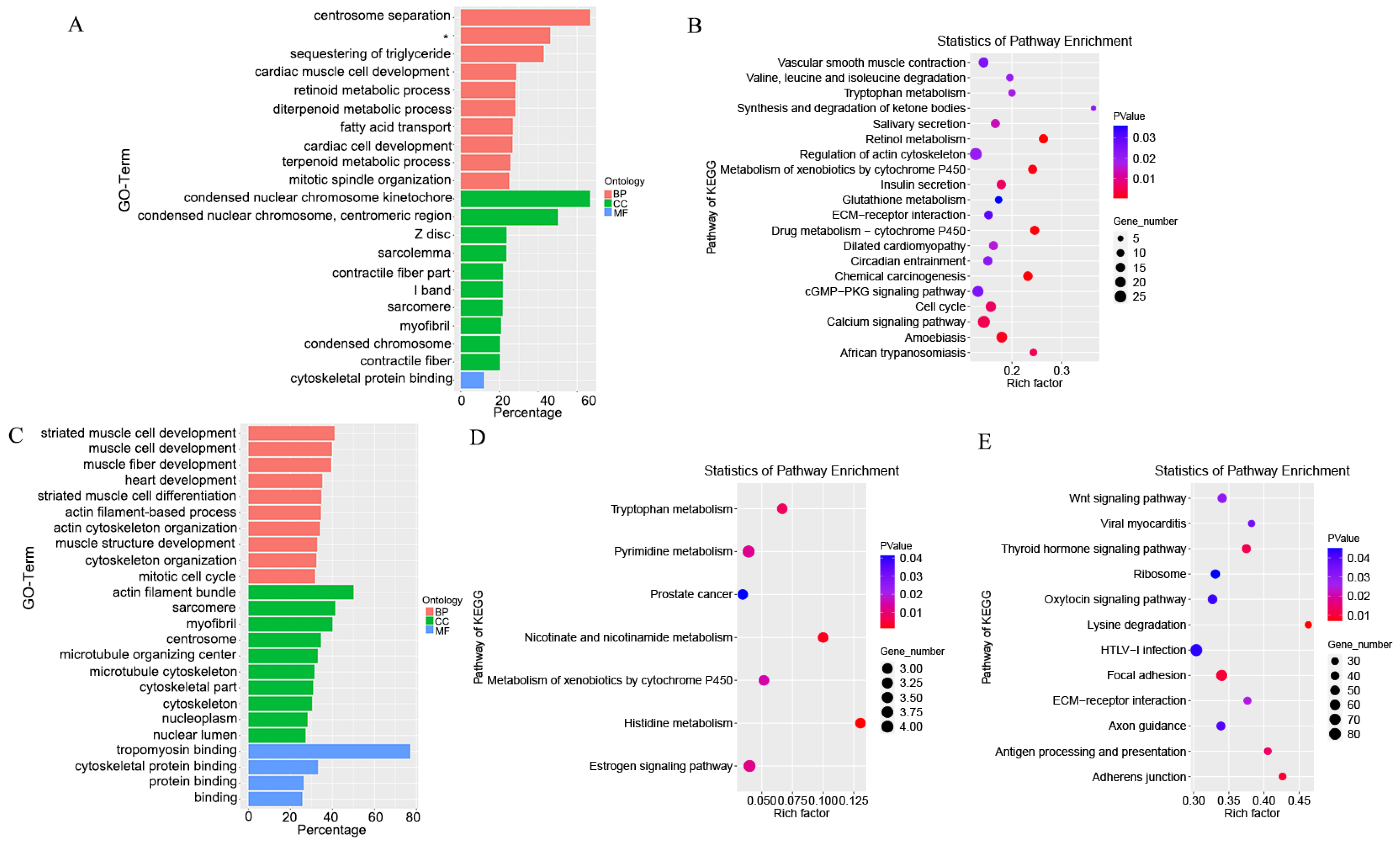
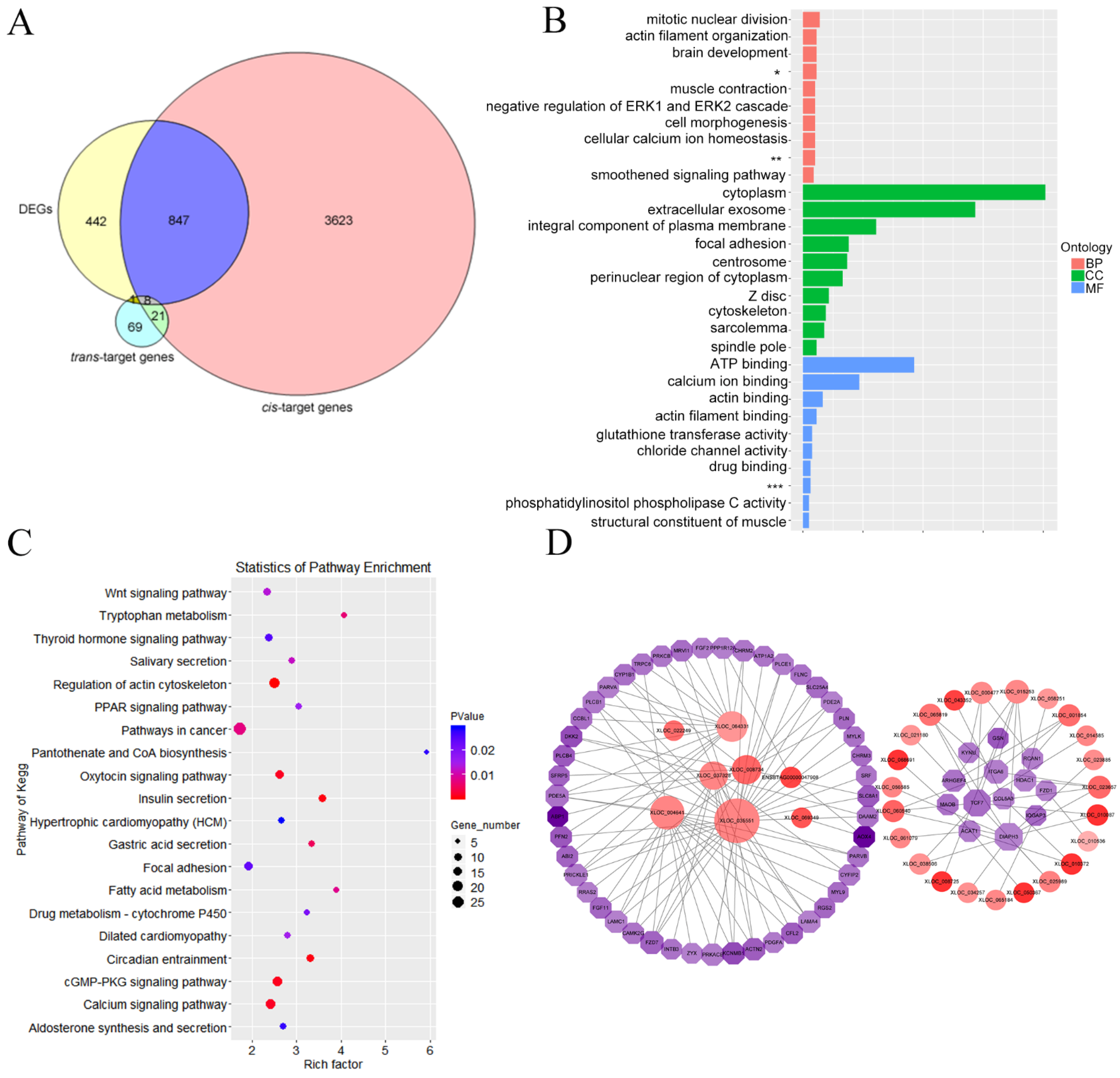
3.6. Target Gene Prediction and Functional Enrichment Analysis
3.7. Integrated Analysis of Differentially Expressed lncRNAs and mRNA
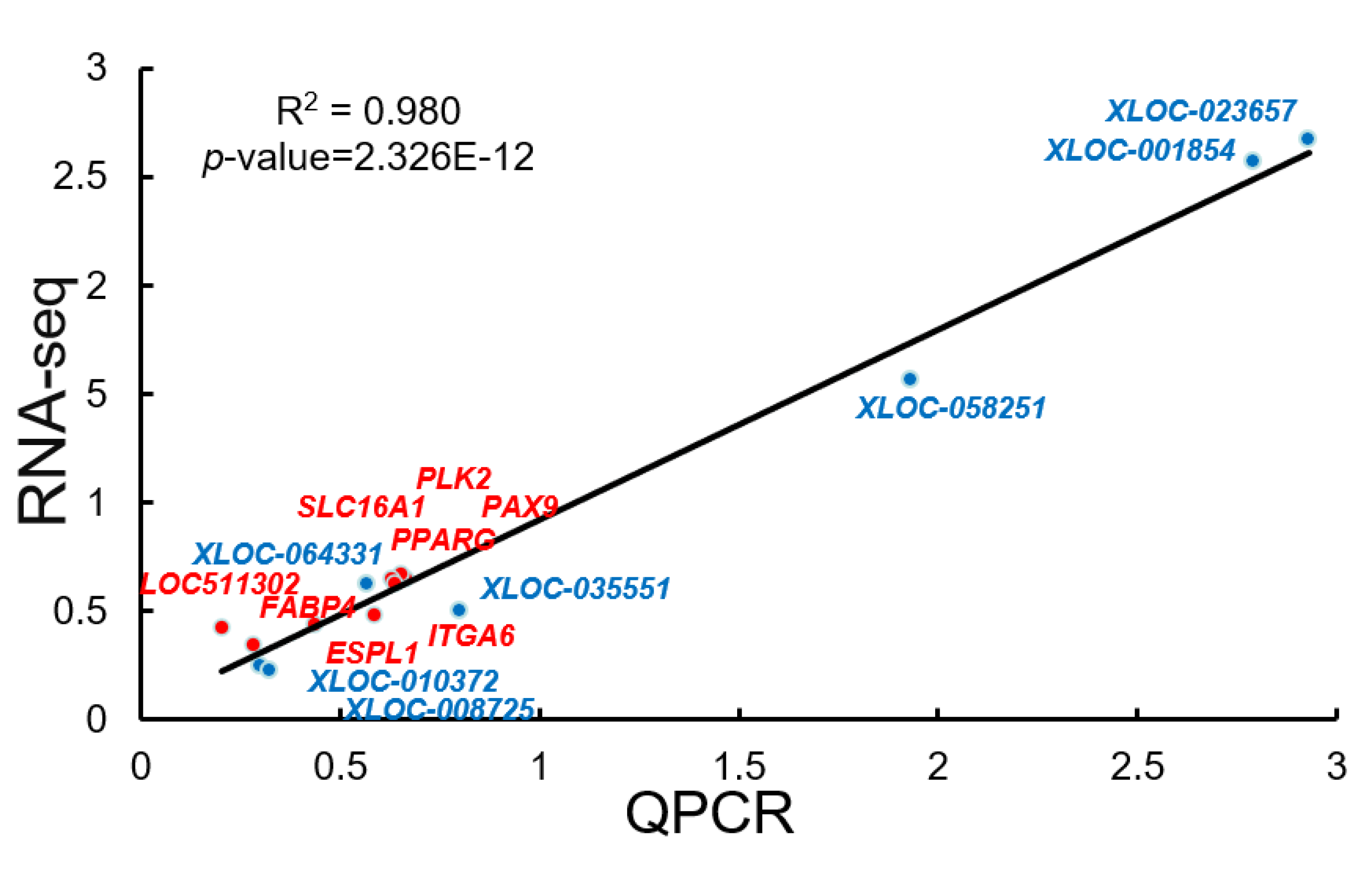
3.8. The qRT-PCR Validation of the Sequencing Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tummler, L.M.; Derno, M.; Rottgen, V.; Vernunft, A.; Tuchscherer, A.; Wolf, P.; Kuhla, B. Effects of 2 colostrum and subsequent milk replacer feeding intensities on methane production, rumen development, and performance in young calves. J. Dairy Sci. 2020, 103, 6054–6069. [Google Scholar] [CrossRef]
- Woodruff, K.; Hummel, G.; Austin, K.; Smith, T.; Cunningham, H. Influence of the maternal rumen microbiome on development of the calf meconium and rumen microbiome. J. Anim. Sci. 2020, 98, 195. [Google Scholar] [CrossRef]
- Van Niekerk, J.K.; Middeldorp, M.; Guan, L.L.; Steele, M.A. Preweaning to postweaning rumen papillae structural growth, ruminal fermentation characteristics, and acute-phase proteins in calves. J. Dairy Sci. 2021, 104, 3632–3645. [Google Scholar] [CrossRef]
- Wu, S.; Cui, Z.; Chen, X.; Zheng, L.; Ren, H.; Wang, D.; Yao, J. Diet-ruminal microbiome-host crosstalk contributes to differential effects of calf starter and alfalfa hay on rumen epithelial development and pancreatic α-amylase activity in yak calves. J. Dairy Sci. 2021, 104, 4326–4340. [Google Scholar] [CrossRef] [PubMed]
- Niwinska, B.; Hanczakowska, E.; Arciszewski, M.B.; Klebaniuk, R. Review: Exogenous butyrate: Implications for the functional development of ruminal epithelium and calf performance. Animal 2017, 11, 1522–1530. [Google Scholar] [CrossRef]
- Suarez, B.J.; Van Reenen, C.G.; Gerrits, W.J.J.; Stockhofe, N.; van Vuuren, A.M.; Dijkstra, J. Effects of supplementing concentrates differing in carbohydrate composition in veal calf diets: II. Rumen development. J. Dairy Sci. 2006, 89, 4376–4386. [Google Scholar] [CrossRef]
- Xie, B.; Huang, W.Q.; Zhang, C.X.; Diao, Q.Y.; Cui, K.; Chai, J.M.; Wang, S.Q.; Lv, X.K.; Zhang, N.F. Influences of starter NDF level on growth performance and rumen development in lambs fed isocaloric and isonitrogenous diets. J. Anim. Sci. 2020, 98, skaa093. [Google Scholar] [CrossRef] [PubMed]
- Beiranvand, H.; Ghorbani, G.R.; Khorvash, M.; Nabipour, A.; Dehghan-Banadaky, M.; Homayouni, A.; Kargar, S. Interactions of alfalfa hay and sodium propionate on dairy calf performance and rumen development. J. Dairy Sci. 2014, 97, 2270–2280. [Google Scholar] [CrossRef] [PubMed]
- Moolchand, M.; Wang, J.; Gui, H.; Shen, Z. Ruminal butyrate infusion increased papillae size and digesta weight but did not change liquid flow rate in the rumen of goats. J. Anim. Plant Sci. 2013, 23, 1516. [Google Scholar] [CrossRef]
- Engreitz, J.M.; Haines, J.E.; Perez, E.M.; Munson, G.; Chen, J.; Kane, M.; McDonel, P.E.; Guttman, M.; Lander, E.S. Local regulation of gene expression by lncRNA promoters, transcription and splicing. Nature 2016, 539, 452–455. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, B.B.; Oh, Y.M.; Zhu, C.C.; Steinmetz, L.M.; Lee, Y.Y.; Kim, W.K.; Lee, S.B.; Buratowski, S.; Kim, T. Modulation of mRNA and lncRNA expression dynamics by the Set2-Rpd3S pathway. Nat. Commun. 2016, 7, 16122. [Google Scholar] [CrossRef]
- Li, Z.W.; Hou, P.F.; Fan, D.M.; Dong, M.C.; Ma, M.S.; Li, H.Y.; Yao, R.S.; Li, Y.X.; Wang, G.N.; Geng, P.Y.; et al. The degradation of EZH2 mediated by lncRNA ANCR attenuated the invasion and metastasis of breast cancer. Cell Death Differ. 2017, 24, 59–71. [Google Scholar] [CrossRef]
- Wang, W.M.; Li, C.; Li, F.; Wang, X.J.; Zhang, X.X.; Liu, T.; Nian, F.; Yue, X.P.; Li, F.; Pan, X.Y.; et al. Effects of early feeding on the host rumen transcriptome and bacterial diversity in lambs. Sci. Rep. 2016, 6, 32479. [Google Scholar] [CrossRef]
- Ibeagha-Awemu, E.M.; Do, D.N.; Dudemaine, P.L.; Fomenky, B.E.; Bissonnette, N. Integration of lncRNA and mRNA Transcriptome Analyses Reveals Genes and Pathways Potentially Involved in Calf Intestinal Growth and Development during the Early Weeks of Life. Genes 2018, 9, 142. [Google Scholar] [CrossRef]
- Sla, B.; Lfa, C.; Xka, D.; Sla, E.; Mei, L.; Eec, A.; Rlbv, A.; Gl, A.; Cjl, A. Establishment and transcriptomic analyses of a cattle rumen epithelial primary cells (REPC) culture by bulk and single-cell RNA sequencing to elucidate interactions of butyrate and rumen development. J. Heliyon 2020, 6, e04112. [Google Scholar] [CrossRef]
- NY/T 815-2004; Beef Cattle Feeding Standards of China. 2004. Available online: https://www.chinesestandard.net/Related.aspx/NYT815-2004 (accessed on 20 December 2021).
- Lesmeister, K.E.; Tozer, P.R.; Heinrichs, A.J. Development and analysis of a rumen tissue sampling procedure. J. Dairy Sci. 2004, 87, 1336–1344. [Google Scholar] [CrossRef]
- Krueger, W.K.; Gutierrez- Bañuelos, H.; Carstens, G.E.; Min, B.R.; Pinchak, W.E.; Gomez, R.R.; Anderson, R.C.; Krueger, N.A.; Forbes, T. Effects of dietary tannin source on performance, feed efficiency, ruminal fermentation, and carcass and non-carcass traits in steers fed a high-grain diet. Anim. Feed Sci. Tech. 2010, 159, 1–9. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Sun, L.; Luo, H.T.; Bu, D.C.; Zhao, G.G.; Yu, K.T.; Zhang, C.H.; Liu, Y.N.; Chen, R.S.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef]
- Kenneth, J.L.; Thomas, D.S. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2002, 25, 402–408. [Google Scholar] [CrossRef]
- Zanin, E.; Henrique, D.S.; Fluck, A.C. Evaluation of equations for estimating the consumption of dairy cows. Rev. Brasil. Saude Pro. Anim. 2017, 18, 76–88. [Google Scholar] [CrossRef][Green Version]
- Vi, R.L.B.; McLeod, K.R.; Klotz, J.L.; Heitmann, R.N. Rumen Development, Intestinal Growth and Hepatic Metabolism In The Pre- and Postweaning Ruminant. J. Dairy Sci. 2004, 87, 55–65. [Google Scholar] [CrossRef]
- Lesmeister, K.E.; Heinrichs, A.J. Effects of adding extra molasses to a texturized calf starter on rumen development, growth characteristics, and blood parameters in neonatal dairy calves. J. Dairy Sci. 2005, 88, 411–418. [Google Scholar] [CrossRef]
- Rice, E.M.; Aragona, K.M.; Moreland, S.C.; Erickson, P.S. Supplementation of sodium butyrate to postweaned heifer diets: Effects on growth performance, nutrient digestibility, and health. J. Dairy Sci. 2019, 102, 3121–3130. [Google Scholar] [CrossRef]
- Greenwood, R.H.; Morrill, J.L.; Titgemeyer, E.C.; Kennedy, G.A. A new method of measuring diet abrasion and its effect on the development of the forestomach. J. Dairy Sci. 1997, 80, 2534. [Google Scholar] [CrossRef]
- Li, R.Y.; Li, B.J.; Jiang, A.W.; Cao, Y.; Hou, L.M.; Zhang, Z.K.; Zhang, X.Y.; Liu, H.L.; Kim, K.H.; Wu, W.J. Exploring the lncRNAs Related to Skeletal Muscle Fiber Types and Meat Quality Traits in Pigs. Genes 2020, 11, 883. [Google Scholar] [CrossRef]
- Sun, J.; Lin, Y.; Wu, J. Long Non-Coding RNA Expression Profiling of Mouse Testis during Postnatal Development. PLoS ONE 2013, 8, e75750. [Google Scholar] [CrossRef]
- Helassa, N.; Nugues, C.; Rajamanoharan, D.; Burgoyne, R.D.; Haynes, L.P. A centrosome-localized calcium signal is essential for mammalian cell mitosis. FASEB J. 2019, 33, 14602–14610. [Google Scholar] [CrossRef]
- Crncec, A.; Hochegger, H. Triggering mitosis. FEBS Lett. 2019, 593, 2868–2888. [Google Scholar] [CrossRef]
- Farshadi, E.; Yan, J.; Leclere, P.; Goldbeter, A.; Chaves, I.; van der Horst, G.T.J. The positive circadian regulators CLOCK and BMAL1 control G2/M cell cycle transition through Cyclin B1. Cell Cycle 2019, 18, 16–33. [Google Scholar] [CrossRef] [PubMed]
- Thompson, W.J.; Piazza, G.A.; Li, H. Exisulind induction of apoptosis involves guanosine 3′,5′-cyclic monophosphate phosphodiesterase inhibition, protein kinase G activation, and attenuated beta-catenin. Cancer Res. 2000, 60, 3338–3342. [Google Scholar] [PubMed]
- Xiao, H.; Cheng, O.M.; Qiu, H.M.; Yang, J.X.; Ma, X.J.; Jiang, Q.S. PDE9 inhibition promotes proliferation of neural stem cells via cGMP-PKG pathway following oxygen-glucose deprivation/reoxygenation injury in vitro. Neurochem. Int. 2020, 133, 104630. [Google Scholar] [CrossRef]
- Lee, D.I.; Zhu, G.S.; Sasaki, T.; Cho, G.S.; Hamdani, N.; Holewinski, R.; Jo, S.H.; Danner, T.; Zhang, M.L.; Rainer, P.P.; et al. Phosphodiesterase 9A controls nitric-oxide-independent cGMP and hypertrophic heart disease. Nature 2015, 519, 472–476. [Google Scholar] [CrossRef]
- Li, N.; Xi, Y.G.; Tinsley, H.N.; Gurpinar, E.; Gary, B.D.; Zhu, B.; Li, Y.H.; Chen, X.; Keeton, A.B.; Abadi, A.H.; et al. Sulindac Selectively Inhibits Colon Tumor Cell Growth by Activating the cGMP/PKG Pathway to Suppress Wnt/beta-Catenin Signaling. Mol. Cancer Ther. 2013, 12, 1848–1859. [Google Scholar] [CrossRef]
- Boyer, A.; Goff, A.K.; Boerboom, D. WNT signaling in ovarian follicle biology and tumorigenesis. Trends Endocrin. Met. 2010, 21, 25–32. [Google Scholar] [CrossRef]
- Sirakov, M.; Boussouar, A.; Kress, E.; Frau, C.; Plateroti, M. The thyroid hormone nuclear receptor TRα1 controls the Notch signaling pathway and cell fate in murine intestine. Development 2015, 142, 2764–2774. [Google Scholar] [CrossRef]
- Cicatiello, A.G.; Ambrosio, R.; Entice, M.D. Thyroid hormone promotes differentiation of colon cancer stem cells. Mol. Cell. Endocrinol. 2017, 459, 84–89. [Google Scholar] [CrossRef]
- Frau, C.; Godart, M.; Plateroti, M. Thyroid hormone regulation of intestinal epithelial stem cell biology. Mol. Cell. Endocrinol. 2017, 459, 90–97. [Google Scholar] [CrossRef]
- Catalano, V.; Dentice, M.; Ambrosio, R.; Luongo, C.; Carollo, R.; Benfante, A.; Todaro, M.; Stassi, G.; Salvatore, D. Activated Thyroid Hormone Promotes Differentiation and Chemotherapeutic Sensitization of Colorectal Cancer Stem Cells by Regulating Wnt and BMP4 Signaling. Cancer Res. 2016, 76, 1237–1244. [Google Scholar] [CrossRef]
- Buznikov, G.A.; Lambert, W.H.; Lauder, J.M. Serotonin and serotonin-like substances as regulators of early embryogenesis and morphogenesis. Cell Tissue Res. 2001, 305, 177–186. [Google Scholar] [CrossRef]
- Whibley, A.; Urquhart, J.; Dore, J.; Willatt, L.; Parkin, G.; Gaunt, L.; Black, G.; Donnai, D.; Raymond, F.L. Deletion of MAOA and MAOB in a male patient causes severe developmental delay, intermittent hypotonia and stereotypical hand movements. Eur. J. Hum. Genet. 2010, 18, 1095–1099. [Google Scholar] [CrossRef]
- Li, Q.; Shen, K.; Zhao, Y.; He, X.; Ma, C.; Wang, L.; Wang, B.; Liu, J.; Ma, J. MicroRNA-222 promotes tumorigenesis via targeting DKK2 and activating the Wnt/β-catenin signaling pathway. FEBS Lett. 2013, 595, 1388. [Google Scholar] [CrossRef]
- Sadeghi, S.; Poorebrahim, M.; Rahimi, H.; Karimipoor, M.; Azadmanesh, K.; Khorramizadeh, M.R.; Teimoori-Toolabi, L. In silico studying of the whole protein structure and dynamics of Dickkopf family members showed that N-terminal domain of Dickkopf 2 in contrary to other Dickkopfs facilitates its interaction with low density lipoprotein receptor related protein 5/6. J. Biomol. Struct. Dyn. 2019, 37, 2564–2580. [Google Scholar] [CrossRef]
- Mukhopadhyay, M.; Gorivodsky, M.; Shtrom, S.; Grinberg, A.; Niehrs, C.; Morasso, M.I.; Westphal, H. Dkk2 plays an essential role in the corneal fate of the ocular surface epithelium. Development 2006, 133, 2149–2154. [Google Scholar] [CrossRef]
- Horváth, D.; Sipos, A.; Major, E.; Kónya, Z.; Bátori, R.; Dedinszki, D.; Szöllősi, A.; Támas, I.; Iván, J.; Kiss, A. Myosin phosphatase accelerates cutaneous wound healing by regulating migration and differentiation of epidermal keratinocytes via Akt signaling pathway in human and murine skin. Mol. Basis Dis. 2018, 1864, 3268–3280. [Google Scholar] [CrossRef]
- Nelson, R. PPP1R12A Pathogenic Variants Associated with Human Congenital Malformations Syndrome. Am. J. Med. Genet. A 2020, 182, 616–617. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment (1) | |||
|---|---|---|---|
| Items | H Group | M Group | L Group |
| Ingredients (% of dry matter) | |||
| Corn | 43.62 | 48.00 | 30.03 |
| Wheat bran | 15.00 | 0.00 | 0.00 |
| Soybean meal | 2.90 | 4.30 | 2.57 |
| Dried distiller’s grains with soluble | 15.00 | 15.00 | 15.00 |
| Alfafa | 20.00 | 25.00 | 35.00 |
| Chinese wild rye | 0.00 | 5.00 | 15.00 |
| Limestone | 0.20 | 0.61 | 0.21 |
| CaHPO4 | 1.78 | 0.59 | 0.69 |
| Premix (2) | 1.00 | 1.00 | 1.00 |
| NaCl | 0.50 | 0.50 | 0.50 |
| Total | 100.00 | 100.00 | 100.00 |
| Nutrient levels (% of dry matter, unless noted) | |||
| Dry matter | 91.8 | 90.5 | 91.58 |
| Crude protein | 16.34 | 16.42 | 16.38 |
| Ether extract | 3.71 | 3.54 | 3.82 |
| Ash | 7.57 | 7.93 | 7.44 |
| Neutral detergent fiber | 34.43 | 37.14 | 45.33 |
| Acid detergent fiber | 15.34 | 18.33 | 25.44 |
| Calcium | 1.05 | 1.08 | 1.14 |
| Phosphorus | 0.45 | 0.45 | 0.47 |
| ME/(MJ/kg) (3) | 11.20 | 10.87 | 9.79 |
| NFC/NDF (4) | 1.10 | 0.94 | 0.60 |
| Accession NO. | Gene Symbol | Primer Sequence (5′ to 3′) | Annealing Temperature (°C) | Product Size |
|---|---|---|---|---|
| NM_174314.2 | FABP4 | F: AGTTTGAATGGGGGTGTGGT | 58 | 199 bp |
| R: CGAGTTTTCTCTTTATGGTGGT | ||||
| NM_001037319.1 | SLC16A1 | F: ATGCCACCACCAGTGAAGTG | 60 | 216 bp |
| R: GCCCAAGACCTCCAATGACT | ||||
| NM_001192369.1 | PAX9 | F: AGTATTCGTGAACGGGAGGC | 60 | 164 bp |
| R: GCAAGATCGAGCCTGTCTCA | ||||
| NM_001192245.1 | PLK2 | F: TCTCCATCACAAGCACGTCG | 60 | 273 bp |
| R: GCCAAACCAAAGTCCCCAAC | ||||
| NM_001109981.2 | ITGA6 | F: CGAAGCAGGAATCCCGAGAC | 60 | 296 bp |
| R: TCCACCAACTTCATAAGGCCC | ||||
| NM_181024.2 | PPARG | F: CAAGAGCTGACCCGATGGTT | 60 | 193 bp |
| R: CCTGACGCTTTATCCCCACA | ||||
| NM_001045949.2 | ESPL1 | F: TGAAGCCAGGCACCTATCC | 58 | 195 bp |
| R: CCATCTTGACCCGAACCCA |
| Items | Treatment 1 | SEM 2 | p-Value | ||
|---|---|---|---|---|---|
| H Group | M Group | L Group | |||
| Initial body weight/kg | 95.04 | 93.82 | 93.23 | 9.27 | 0.481 |
| Final body weight/kg | 190.88 | 183.43 | 171.27 | 2.96 | 0.964 |
| Body weight gain/kg | 92.73 a | 77.07 b | 70.87 b | 3.12 | 0.009 |
| Average daily gain/kg | 1.14 a | 1.00 b | 0.93 b | 0.03 | 0.008 |
| Dry matter intake/(kg/d) | 4.09 | 3.81 | 3.85 | 0.11 | 0.605 |
| Feed to gain ratio | 3.65 | 3.81 | 4.15 | 0.09 | 0.169 |
| Items | Treatments 1 | SEM 2 | p-Value | ||
|---|---|---|---|---|---|
| H Group | M Group | L Group | |||
| pH | 6.81 | 7.00 | 6.83 | 0.06 | 0.419 |
| Acetate | 53.43 b | 59.92 a | 60.71 a | 1.24 | 0.026 |
| Propionate (mmol/L) | 25.77 a | 21.66 ab | 18.96 b | 1.01 | 0.014 |
| Butyrate (mmol/L) | 13.36 | 12.26 | 12.76 | 0.80 | 0.875 |
| Papillae length (mm) | 0.75 b | 0.92 b | 1.11 a | 0.08 | 0.023 |
| Papillae width (mm) | 0.38 a | 0.34 a | 0.24 b | 0.01 | <0.001 |
| Muscle layer thickness (mm) | 1.21 | 1.23 | 1.35 | 0.04 | 0.144 |
| Items | H-1 1 | H-2 2 | H-3 3 | L-1 4 | L-2 5 | L-3 6 |
|---|---|---|---|---|---|---|
| Raw reads | 129,008,240 | 97,533,606 | 113,893,888 | 84,213,254 | 89,185,784 | 99,002,628 |
| Clean reads | 126,165,758 | 95,293,260 | 111,116,586 | 81,352,198 | 87,101,750 | 96,764,402 |
| Percentage (%) | 97.80 | 97.70 | 97.56 | 96.60 | 97.66 | 97.74 |
| Mapped reads | 104,668,662 | 80,985,576 | 93,383,745 | 67,890,749 | 72,068,883 | 82,345,847 |
| Mapping rate (%) | 82.96 | 84.99 | 84.04 | 83.45 | 82.74 | 85.10 |
| Q30 (%) | 93.64 | 93.23 | 92.68 | 91.93 | 93.83 | 93.33 |
| Spliced reads (%) | 20.59 | 18.07 | 21.56 | 22.23 | 20.8 | 22.1 |
| Non-spliced reads (%) | 59.65 | 64.54 | 59.59 | 57.47 | 58.57 | 60.21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Xue, M.; Zhang, L.; Li, L.; Lian, H.; Li, M.; Gao, T.; Fu, T.; Tu, Y. Integration of Long Non-Coding RNA and mRNA Profiling Reveals the Mechanisms of Different Dietary NFC/NDF Ratios Induced Rumen Development in Calves. Animals 2022, 12, 650. https://doi.org/10.3390/ani12050650
Li J, Xue M, Zhang L, Li L, Lian H, Li M, Gao T, Fu T, Tu Y. Integration of Long Non-Coding RNA and mRNA Profiling Reveals the Mechanisms of Different Dietary NFC/NDF Ratios Induced Rumen Development in Calves. Animals. 2022; 12(5):650. https://doi.org/10.3390/ani12050650
Chicago/Turabian StyleLi, Jichao, Mingming Xue, Liyang Zhang, Lanjie Li, Hongxia Lian, Ming Li, Tengyun Gao, Tong Fu, and Yan Tu. 2022. "Integration of Long Non-Coding RNA and mRNA Profiling Reveals the Mechanisms of Different Dietary NFC/NDF Ratios Induced Rumen Development in Calves" Animals 12, no. 5: 650. https://doi.org/10.3390/ani12050650
APA StyleLi, J., Xue, M., Zhang, L., Li, L., Lian, H., Li, M., Gao, T., Fu, T., & Tu, Y. (2022). Integration of Long Non-Coding RNA and mRNA Profiling Reveals the Mechanisms of Different Dietary NFC/NDF Ratios Induced Rumen Development in Calves. Animals, 12(5), 650. https://doi.org/10.3390/ani12050650