
Feed Supplemented with Aronia melanocarpa (AM) Relieves the Oxidative Stress Caused by Ovulation in Peak Laying Hens and Increases the Content of Yolk Precursors
 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Moral Statement
2.2. Plant Material and Animal Preparation
2.3. Sample Collection for Blood and Tissues
2.4. Organ Index and Morphological Observation
2.5. Determination of AST and ALT
2.6. Determination of Antioxidant Parameters
2.7. Assays of Estrogen Levels
2.8. Assays of Yolk Precursor Content and Lipid Parameters
2.9. RNA Isolation and Quantitative Real-Time PCR (qRT-PCR)
2.10. Protein Extraction and Western Blot Analysis
2.11. Production Performance and Egg Quality Determination
2.12. Statistical Analysis
3. Results
3.1. Effects of Feed Supplemented with AM on Organ Indices of Laying Hens
3.2. Effects of Feed Supplemented with AM on Histomorphology during Peak Egg Production
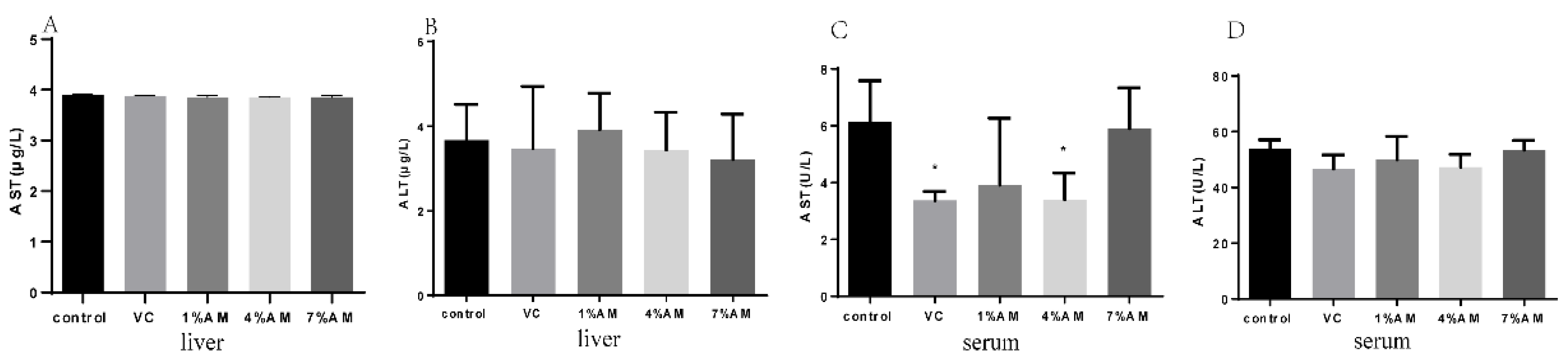
3.3. Effects of Feed Supplemented with AM on AST and ALT
3.4. Effects of Feed Supplemented with AM on Antioxidant Parameters
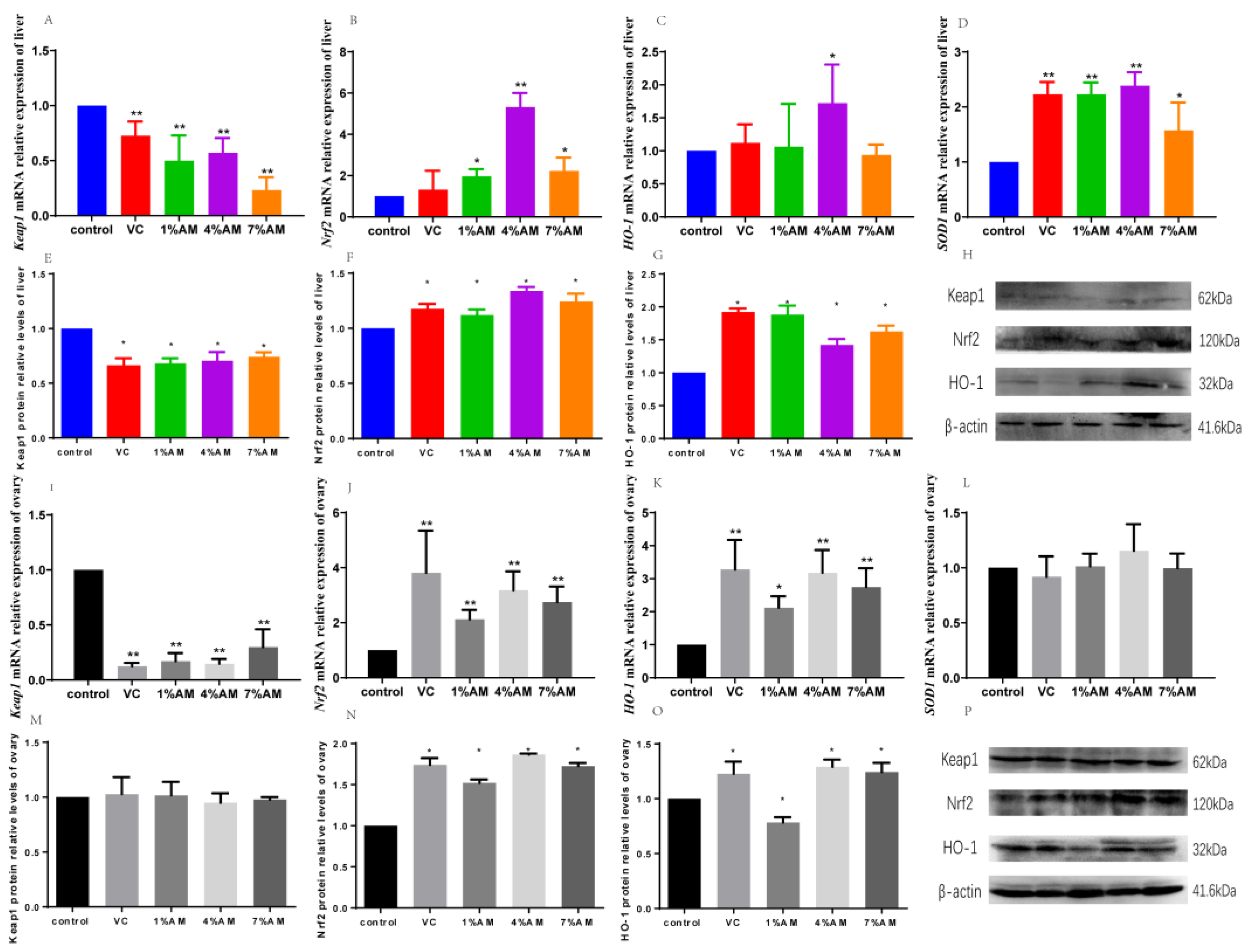
3.5. Effects of Feed Supplemented with AM on Keap1/Nrf2 Pathway
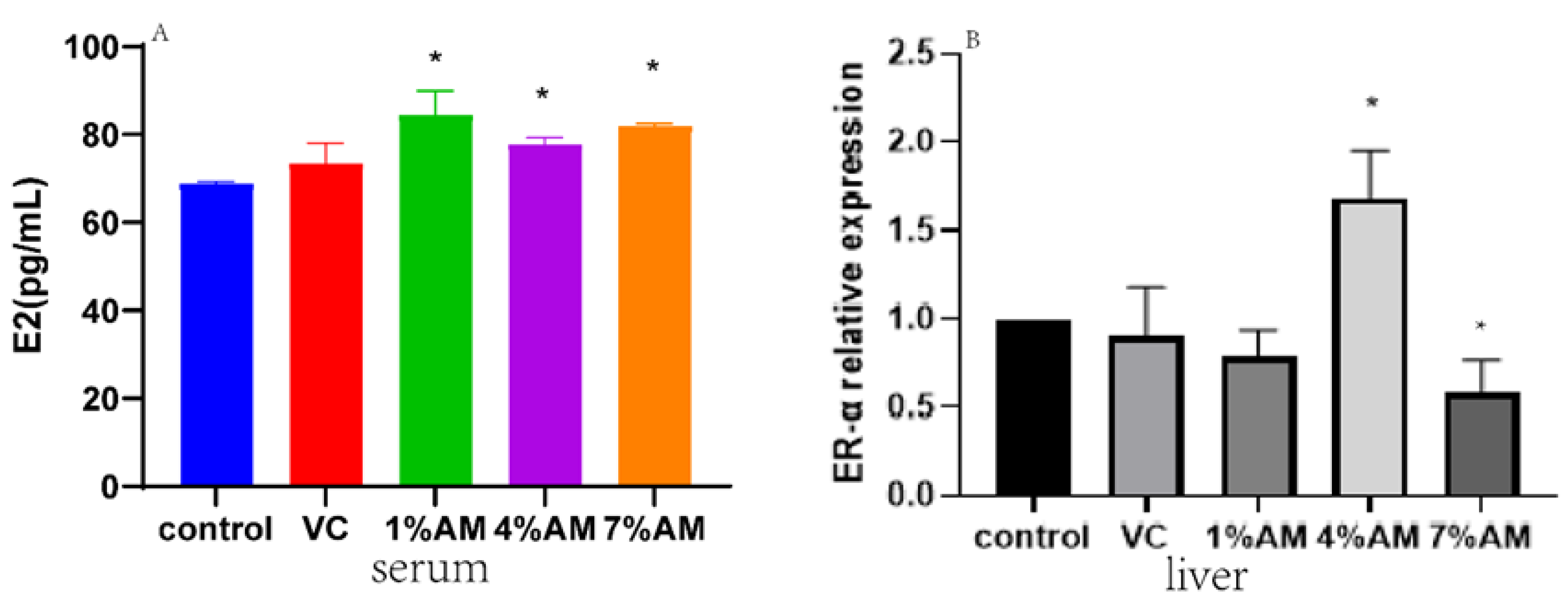
3.6. Effects of Feed Supplemented with AM on Estrogen Levels and Their Receptors during Peak Laying Period
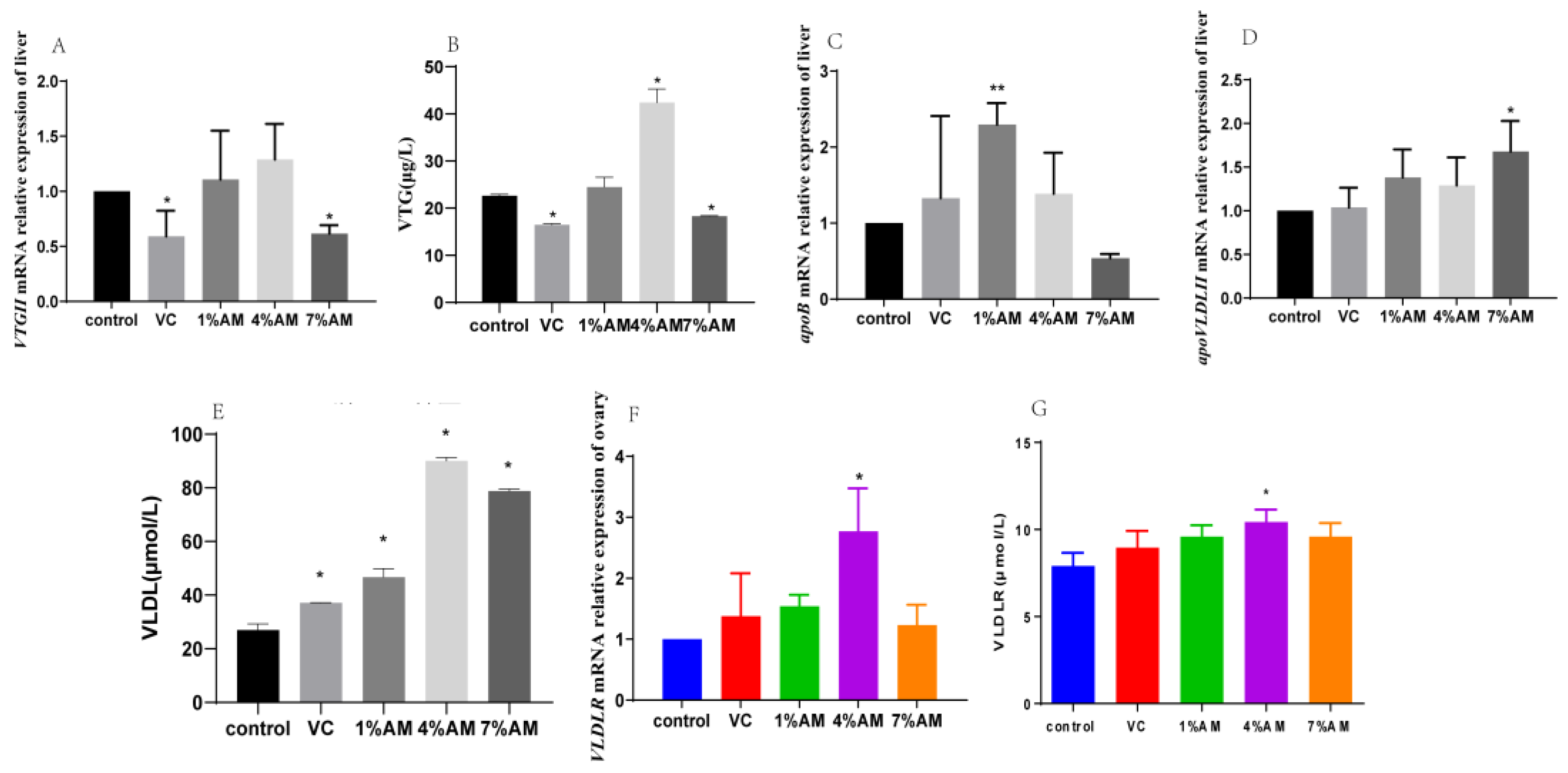
3.7. Effects of Feed Supplemented with AM on Lipid Parameters
3.8. Effects of Feed Supplemented with AM on Gene and Protein Expression of Yolk Precursor and Its Receptor
3.9. Effects of Feed Supplemented with AM on Changes in Yolk Deposition during Peak Laying Period
3.10. Effects of Feed Supplemented with AM on Production Performance and Egg Quality of Peak Laying Hens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scappaticcio, R.; García, J.; Fondevila, G.; de Juan, A.F.; Cámara, L.; Mateos, G.G. Influence of the energy and digestible lysine contents of the diet on performance and egg quality traits of brown-egg laying hens from 19 to 59 weeks of age. Poult. Sci. 2021, 100, 101211. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Chen, Y.; Liu, Y.; Xing, Y.; Miao, C.; Zhao, Y.; Chang, X.; Zhang, Q. The Role of Oxidative Stress and Natural Antioxidants in Ovarian Aging. Front. Pharmacol. 2020, 11, 617843. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Ma, Y.; Lin, X.; Zhou, S.; Mi, Y.; Zhang, C. The Attenuating Effect of the Intraovarian Bone Morphogenetic Protein 4 on Age-Related Endoplasmic Reticulum Stress in Chicken Follicular Cells. Oxid. Med. Cell. Longev. 2020, 2020, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Bendikov-Bar, I.; Malitsky, S.; Itkin, M.; Rusal, M.; Sagi, D.; Le Couteur, D. Metabolomic Changes Are Predictive of Aging in Laying Hens. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2021, 76, 1757–1768. [Google Scholar] [CrossRef]
- Zhou, D.; Luo, M.; Shang, A.; Mao, Q.; Li, B.; Gan, R.; Li, H. Antioxidant Food Components for the Prevention and Treatment of Cardiovascular Diseases: Effects, Mechanisms, and Clinical Studies. Oxid. Med. Cell. Longev. 2021, 2021, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Tao, Z.; Chen, X.; Xiao, J.; Zhang, Y.; Wang, Z. Effects of broussonetia papyrifera-fermented feed on production performance, egg quality, and caecal microbiota of laying hens during the late laying period. Ital. J. Anim. Sci. 2022, 21, 659–672. [Google Scholar] [CrossRef]
- Liu, X.; Lin, X.; Mi, Y.; Li, J.; Zhang, C. Grape Seed Proanthocyanidin Extract Prevents Ovarian Aging by Inhibiting Oxidative Stress in the Hens. Oxid. Med. Cell. Longev. 2018, 2018, 1–16. [Google Scholar] [CrossRef]
- Grabowska, M.G.; Wawrzyniak, D.; Rolle, K.; Chomczy Ski, P.; Oziewicz, S.; Jurga, S.; Barciszewski, J. Let food be your medicine: Nutraceutical properties of lycopene. Food Funct. 2019, 1, 39–312. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Wang, J.; Wang, H.; Zhang, H.; Wu, S.; Qi, G. Dietary tea polyphenol supplementation improved egg production performance, albumen quality, and magnum morphology of Hy-Line Brown hens during the late laying period1. J. Anim. Sci. 2018, 96, 225–235. [Google Scholar] [CrossRef]
- Jurendić, T.; Ščetar, M. Aronia melanocarpa Products and By-Products for Health and Nutrition: A Review. Antioxidants 2021, 10, 1052. [Google Scholar] [CrossRef]
- Bräunlich, M.; Slimestad, R.; Wangensteen, H.; Brede, C.; Malterud, K.; Barsett, H. Extracts, Anthocyanins and Procyanidins from Aronia melanocarpa as Radical Scavengers and Enzyme Inhibitors. Nutrients 2013, 5, 663–678. [Google Scholar] [CrossRef] [PubMed]
- Staszowska-Karkut, M.; Materska, M. Phenolic Composition, Mineral Content, and Beneficial Bioactivities of Leaf Extracts from Black Currant (Ribes nigrum L.), Raspberry (Rubus idaeus), and Aronia (Aronia melanocarpa). Nutrients 2020, 12, 463. [Google Scholar] [CrossRef] [PubMed]
- Hatefi, A.; Zare Shahneh, A.; Ansari Pirsaraie, Z.; Alizadeh, A.M.; Atashnak, M.P.; Masoudi, R.; Pio, F. Pro-and anti-inflammatory effects of glucocorticoid Fluticasone on ovarian and immune functions in commercial-aged laying hens. Sci. Rep. 2021, 11, 21603. [Google Scholar] [CrossRef] [PubMed]
- Kalt, W.; Cassidy, A.; Howard, L.R.; Krikorian, R.; Stull, A.J.; Tremblay, F.; Zamora-Ros, R. Recent Research on the Health Benefits of Blueberries and Their Anthocyanins. Adv. Nutr. 2020, 11, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Waldbauer, K.; McKinnon, R.; Kopp, B. Apple Pomace as Potential Source of Natural Active Compounds. Planta Med. 2017, 83, 994–1010. [Google Scholar] [CrossRef] [PubMed]
- Warner, R.; Wu, B.; MacPherson, S.; Lefsrud, M. A Review of Strawberry Photobiology and Fruit Flavonoids in Controlled Environments. Front. Plant Sci. 2021, 12, 611893. [Google Scholar] [CrossRef]
- Luo, B.S.; Gu, R.H.; Kennelly, E.J.; Long, C.L. Gaultheria Ethnobotany and Bioactivity: Blueberry Relatives with Anti-inflammatory, Antioxidant, and Anticancer Constituents. Curr. Med. Chem. 2018, 25, 5168–5176. [Google Scholar] [CrossRef]
- Ispiryan, A.; Viškelis, J.; Viškelis, P. Red Raspberry (Rubus idaeus L.) Seed Oil: A Review. Plants 2021, 10, 944. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed]
- Oviedo-Solís, C.I.; Cornejo-Manzo, S.; Murillo-Ortiz, B.O.; Guzmán-Barrón, M.M.; Ramírez-Emiliano, J. Los polifenoles de la fresa disminuyen el estrés oxidativo en enfermedades crónicas. Gac. De Mex. 2018, 154, 80–86. [Google Scholar] [CrossRef]
- Rodrigues, E.L.; Marcelino, G.; Silva, G.T.; Figueiredo, P.S.; Garcez, W.S.; Corsino, J.; Guimarães, R.D.C.A.; Freitas, K.D.C. Nutraceutical and Medicinal Potential of the Morus Species in Metabolic Dysfunctions. Int. J. Mol. Sci. 2019, 20, 301. [Google Scholar] [CrossRef] [PubMed]
- Mężyńska, M.; Brzóska, M.; Rogalska, J.; Galicka, A. Extract from Aronia melanocarpa L. Berries Protects Against Cadmium-induced Lipid Peroxidation and Oxidative Damage to Proteins and DNA in the Liver: A Study using a Rat Model of Environmental Human Exposure to this Xenobiotic. Nutrients 2019, 11, 758. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, X.; Yan, H.; Liu, Y.; Li, L.; Li, S.; Wang, X.; Wang, D. Aronia melanocarpa ameliorates gout and hyperuricemia in animal models. Food Agric. Immunol. 2019, 30, 47–59. [Google Scholar] [CrossRef]
- Bahr, J.M.; Johnson, A.L. Regulation of the follicular hierarchy and ovulation. J. Exp. Zool. 1984, 232, 495–500. [Google Scholar] [CrossRef]
- Parenteau, I.A.; Stevenson, M.; Kiarie, E.G. Egg production and quality responses to increasing isoleucine supplementation in Shaver white hens fed a low crude protein corn-soybean meal diet fortified with synthetic amino acids between 20 and 46 weeks of age. Poult. Sci. 2020, 99, 1444–1453. [Google Scholar] [CrossRef]
- Cao, C.; Tachibana, T.; Gilbert, E.R.; Cline, M.A. Prostaglandin E2-induced anorexia involves hypothalamic brain-derived neurotrophic factor and ghrelin in chicks. Prostaglandins Other Lipid Mediat. 2021, 156, 106574. [Google Scholar] [CrossRef]
- Liu, X.; Lin, X.; Zhang, S.; Guo, C.; Li, J.; Mi, Y.; Zhang, C. Lycopene ameliorates oxidative stress in the aging chicken ovary via activation of Nrf2/HO-1 pathway. Aging 2018, 10, 2016–2036. [Google Scholar] [CrossRef]
- Dai, H.; Lv, Z.; Huang, Z.; Ye, N.; Li, S.; Jiang, J.; Cheng, Y.; Shi, F. Dietary hawthorn-leaves flavonoids improves ovarian function and liver lipid metabolism in aged breeder hens. Poult. Sci. 2021, 100, 101499. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, Y.; Liu, Z.; Guo, X.; Sun, Y.; Kang, L.; Jiang, Y. Transcriptomic and proteomic analyses of ovarian follicles reveal the role of VLDLR in chicken follicle selection. BMC Genom. 2020, 21. [Google Scholar] [CrossRef]
- Chen, X.; He, K.; Wei, C.; Yang, W.; Geng, Z. Green Tea Powder Decreased Egg Weight Through Increased Liver Lipoprotein Lipase and Decreased Plasma Total Cholesterol in an Indigenous Chicken Breed. Animals 2020, 10, 370. [Google Scholar] [CrossRef]
- Tao, Y.; Wang, T.; Huang, C.; Lai, C.; Ling, Z.; Zhou, Y.; Yong, Q. Production performance, egg quality, plasma biochemical constituents and lipid metabolites of aged laying hens supplemented with incomplete degradation products of galactomannan. Poult. Sci. 2021, 100, 101296. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Lei, J.; Liu, L.; Qu, X.; Li, P.; Liu, X.; Guo, Y.; Gao, Q.; Lan, F.; Xiao, B.; et al. Effects of Macleaya cordata extract on laying performance, egg quality, and serum indices in Xuefeng black-bone chicken. Poult. Sci. 2021, 100, 101031. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhang, H.; Du, E.; Jin, F.; Zheng, C.; Fan, Q.; Zhao, N.; Guo, W.; Zhang, W.; Huang, S.; et al. Effects of magnolol on egg production, egg quality, antioxidant capacity, and intestinal health of laying hens in the late phase of the laying cycle. Poult. Sci. 2021, 100, 835–843. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Control | VC | 1% AM | 4% AM | 7% AM | p-Value |
|---|---|---|---|---|---|---|
| Liver index | 27.72 ± 1.48 | 29.94 ± 1.75 | 28.70 ± 1.94 | 29.30 ± 3.52 | 29.84 ± 1.63 | 0.4033 |
| Ovary index | 49.39 ± 5.16 | 50.85 ± 5.89 | 49.43 ± 2.05 | 49.69 ± 4.23 | 51.69 ± 3.80 | 0.8631 |
| Items | Control | VC | 1% AM | 4% AM | 7% AM | p-Value |
|---|---|---|---|---|---|---|
| Liver-GPX | 132.70 ± 8.00 | 131.90 ± 5.49 | 172.00 ± 11.78 ** | 126.60 ± 0.93 | 102.60 ± 17.20 * | <0.0001 |
| Liver-MDA | 2.13 ± 0.12 | 1.95 ± 0.18 | 2.10 ± 0.06 | 1.54 ± 0.46** | 1.77 ± 0.25 | 0.0023 |
| Liver- T-SOD | 13.68 ± 0.87 | 14.00 ± 0.14 | 14.25 ± 0.48 | 14.14 ± 0.22 | 14.48 ± 0.31 | 0.0859 |
| Liver- T-AOC | 0.87 ± 0.04 | 0.93 ± 0.02 ** | 0.94 ± 0.01 ** | 0.90 ± 0.05 | 0.88 ± 0.02 | 0.0024 |
| Ovary-GPX | 133.70 ± 19.12 | 140.40 ± 19.20 * | 182.40 ± 10.90 ** | 225.70 ± 39.00 ** | 113.70 ± 11.92 | <0.0001 |
| Ovary-MDA | 1.49 ± 0.34 | 0.91 ± 0.11* | 1.07 ± 0.34 | 1.19 ± 0.30 | 1.51 ± 0.32 | 0.0055 |
| Ovary-T-SOD | 8.57 ± 0.81 | 20.62 ± 6.28 ** | 16.91 ± 0.92 ** | 15.83 ± 0.90 ** | 12.46 ± 0.58 ** | <0.0001 |
| Ovary-T-AOC | 0.38 ± 0.06 | 0.44 ± 0.02 | 0.42 ± 0.05 | 0.46 ± 0.04 ** | 0.57 ± 0.01 ** | <0.0001 |
| Serum-GPX | 38.46 ± 4.00 | 403.60 ± 61.48 ** | 676.40 ± 205.10 ** | 839.20 ± 160.50 ** | 664.10 ± 220.30 ** | <0.0001 |
| Serum-MDA | 4.95 ± 0.85 | 2.31 ± 0.48 ** | 4.48 ± 0.70 | 3.81 ± 0.83 * | 2.72 ± 0.90 ** | <0.0001 |
| Serum-T-SOD | 84.78 ± 3.11 | 111.00 ± 0.80 ** | 110.90 ± 0.52 ** | 113.30 ± 1.60 ** | 111.30 ± 2.80 ** | <0.0001 |
| Serum-T-AOC | 0.60 ± 0.04 | 0.59 ± 0.02 | 0.72 ± 0.02 ** | 0.66 ± 0.02 * | 0.57 ± 0.01 | <0.0001 |
| Yolk-GPX | 20.53 ± 1.39 | 102.70 ± 21.67 ** | 109.30 ± 6.58 ** | 80.38 ± 5.27 ** | 81.82 ± 7.28 ** | <0.0001 |
| Yolk-MDA | 1.52 ± 0.06 | 0.82 ± 0.07 ** | 0.43 ± 0.08 ** | 1.05 ± 0.18 * | 0.67 ± 0.08 ** | <0.0001 |
| Yolk- T-SOD | 82.64 ± 4.01 | 99.38 ± 3.71 ** | 92.89 ± 4.33 * | 93.83 ± 2.32 ** | 100.60 ± 3.53 ** | <0.0001 |
| Yolk- T-AOC | 0.72 ± 0.03 | 0.71 ± 0.04 | 0.72 ± 0.06 | 0.73 ± 0.01 | 0.73 ± 0.04 | 0.8950 |
| Items | Control | VC | 1% AM | 4% AM | 7% AM | p-Value |
|---|---|---|---|---|---|---|
| F1 (cm) | 3.03 ± 0.08 | 3.23 ± 0.12 | 3.27 ± 0.08 * | 3.30 ± 0.29 * | 3.07 ± 0.10 * | 0.0172 |
| F2 (cm) | 2.60 ± 0.06 | 3.13 ± 0.23 ** | 2.95 ± 0.14 ** | 3.02 ± 0.31 ** | 3.03 ± 0.05 ** | 0.0006 |
| F3 (cm) | 2.33 ± 0.12 | 2.73 ± 0.48 | 2.57 ± 0.15 | 2.70 ± 0.33 | 2.30 ± 0.31 | 0.0609 |
| F4 (cm) | 2.10 ± 0.13 | 2.32 ± 0.38 | 2.10 ± 0.18 | 2.22 ± 0.37 | 2.00 ± 0.24 | 0.3420 |
| F5 (cm) | 1.72 ± 0.33 | 1.93 ± 0.33 | 1.70 ± 0.23 | 1.80 ± 0.26 | 1.53 ± 0.41 | 0.3083 |
| Hierarchical follicles (F1–F5) mass (g) | 44.64 ± 4.88 | 54.19 ± 6.58 * | 47.52 ± 5.07 | 49.19 ± 4.41 | 44.75 ± 2.25 | 0.0128 |
| Items | Control | VC | 1% AM | 4% AM | 7% AM | p-Value |
|---|---|---|---|---|---|---|
| Production Performance | ||||||
| BW/kg | 1.82 ± 0.03 | 1.92 ± 0.03 | 1.95 ± 0.03 * | 1.90 ± 0.03 | 1.89 ± 0.03 | 0.033 |
| EW/g | 62.14 ± 0.88 | 59.97 ± 0.77 | 60.86 ± 0.73 | 60.03 ± 0.60 | 58.61 ± 0.67 * | 0.017 |
| Egg rate % | 86.31 ± 1.67 | 88.14 ± 1.48 | 89.95 ± 1.42 | 89.36 ± 2.23 | 88.46 ± 1.80 | 0.277 |
| ADFI/g | 126.67 ± 3.21 | 116.70 ± 6.92 | 148.80 ± 13.92 | 116.65 ± 12.20 | 130.49 ± 7.84 | 0.166 |
| FER | 2.21 ± 0.07 | 2.06 ± 0.11 | 2.62 ± 0.26 | 2.06 ± 0.21 | 2.30 ± 0.13 | 0.172 |
| Egg Quality | ||||||
| Egg shape index | 1.26 ± 0.01 | 1.25 ± 0.01 | 1.28 ± 0.01 | 1.27 ± 0.01 | 1.26 ± 0.00 | 0.197 |
| Egg yolk color | 9.11 ± 0.14 | 8.86 ± 0.16 | 9.42 ± 0.15 | 8.67 ± 0.16 | 9.17 ± 0.15 | 0.007 |
| ETS/mm | 0.31 ± 0.01 | 0.29 ± 0.01 | 0.31 ± 0.01 | 0.28 ± 0.01 ** | 0.29 ± 0.00 | 0.001 |
| ESH/kg.cm−2 | 54.61 ± 0.58 | 57.89 ± 1.31 | 57.95 ± 1.23 | 55.38 ± 1.04 | 55.94 ± 1.25 | 0.037 |
| PH/mm | 7.57 ± 0.19 | 7.39 ± 0.11 | 7.69 ± 0.17 | 7.95 ± 0.16 | 7.59 ± 0.17 | 0.171 |
| Hastelloy unit | 85.86 ± 1.19 | 85.76 ± 0.62 | 87.14 ± 0.91 | 88.15 ± 0.91 | 87.16 ± 0.94 | 0.134 |
| Yolk weight/mg | 32.00 ± 0.00 | 32.70 ± 0.00 | 34.30 ± 0.00 | 33.80 ± 0.00 | 32.60 ± 0.00 | 0.104 |
| EWW/mg | 72.10 ± 0.01 | 72.30 ± 0.01 | 73.00 ± 0.01 | 72.10 ± 0.01 | 70.00 ± 0.01 | 0.473 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, B.; Xiao, H.; Yin, H.; Wei, Y.; Wu, H.; Zhang, D.; Tsopmejio, I.S.N.; Shang, H.; Jin, Z.; Song, H. Feed Supplemented with Aronia melanocarpa (AM) Relieves the Oxidative Stress Caused by Ovulation in Peak Laying Hens and Increases the Content of Yolk Precursors. Animals 2022, 12, 3574. https://doi.org/10.3390/ani12243574
Jing B, Xiao H, Yin H, Wei Y, Wu H, Zhang D, Tsopmejio ISN, Shang H, Jin Z, Song H. Feed Supplemented with Aronia melanocarpa (AM) Relieves the Oxidative Stress Caused by Ovulation in Peak Laying Hens and Increases the Content of Yolk Precursors. Animals. 2022; 12(24):3574. https://doi.org/10.3390/ani12243574
Chicago/Turabian StyleJing, Bo, Huanwei Xiao, Haixu Yin, Yingbo Wei, Haoyuan Wu, Dongliang Zhang, Ivan Stève Nguepi Tsopmejio, Hongmei Shang, Zhouyu Jin, and Hui Song. 2022. "Feed Supplemented with Aronia melanocarpa (AM) Relieves the Oxidative Stress Caused by Ovulation in Peak Laying Hens and Increases the Content of Yolk Precursors" Animals 12, no. 24: 3574. https://doi.org/10.3390/ani12243574
APA StyleJing, B., Xiao, H., Yin, H., Wei, Y., Wu, H., Zhang, D., Tsopmejio, I. S. N., Shang, H., Jin, Z., & Song, H. (2022). Feed Supplemented with Aronia melanocarpa (AM) Relieves the Oxidative Stress Caused by Ovulation in Peak Laying Hens and Increases the Content of Yolk Precursors. Animals, 12(24), 3574. https://doi.org/10.3390/ani12243574