
Effects of Restricted Availability of Drinking Water on Blood Characteristics and Constituents in Dorper, Katahdin, and St. Croix Sheep from Different Regions of the USA
 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
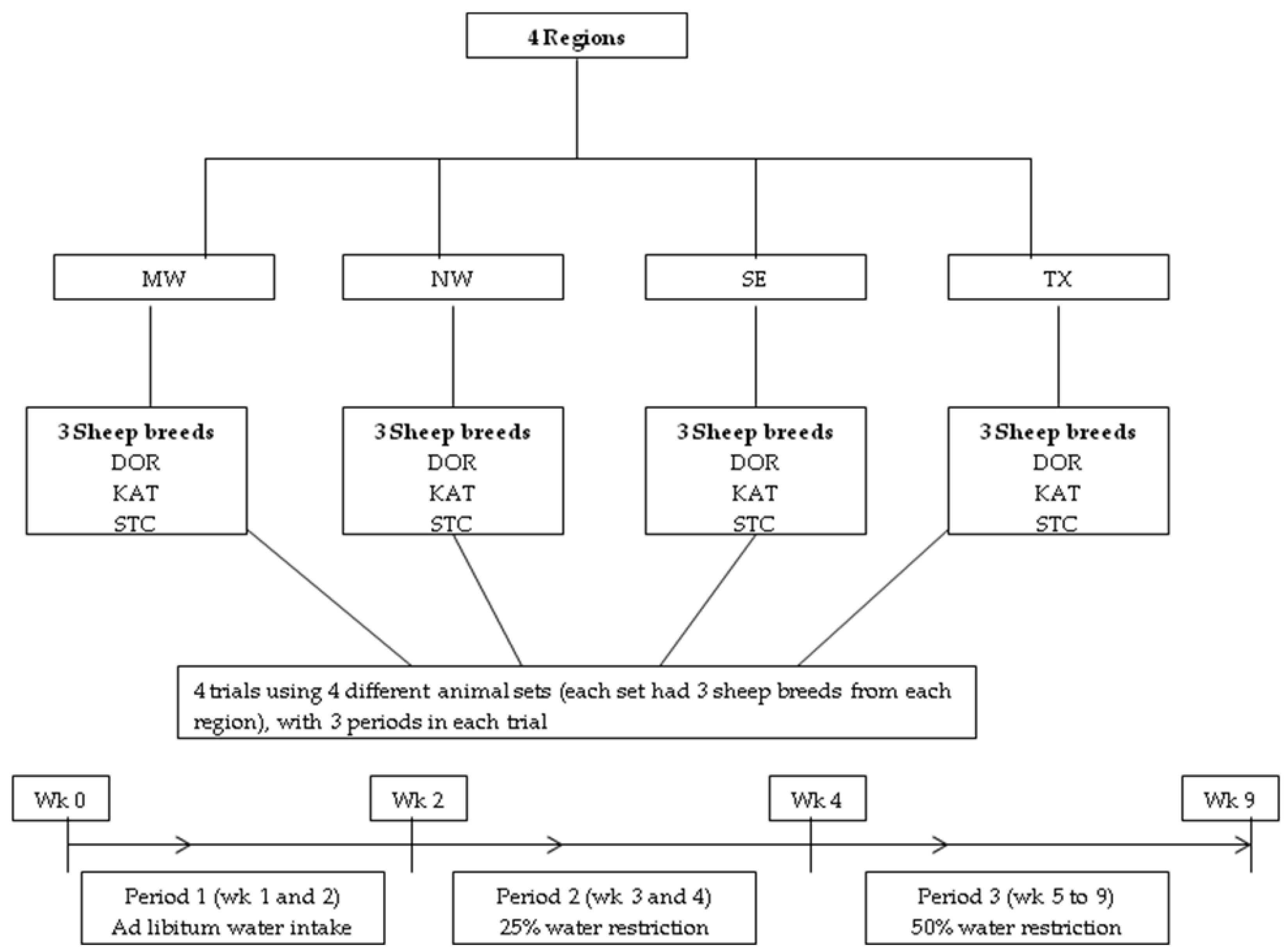
2. Materials and Methods
2.1. Animals, Housing, and Diet
2.2. Periods and Water Intake
2.3. Measures
2.4. Statistical Analysis

{kind=link}
| Source of Variation 2 | Variable 1 | ||||
|---|---|---|---|---|---|
| OSM (mosmol/kg) | PCV (%) | Hb (g/dL) | Hb O2 (%) | O2 (mmol/L) | |
| Set | <0.001 | 0.279 | 0.146 | <0.001 | <0.001 |
| Age | 0.957 | 0.521 | 0.763 | 0.180 | 0.212 |
| Brd | 0.009 | 0.102 | 0.674 | 0.001 | 0.001 |
| Reg | 0.564 | 0.203 | 0.176 | 0.576 | 0.795 |
| Brd *Reg | 0.209 | 0.377 | 0.567 | 0.203 | 0.421 |
| Prd | <0.001 | 0.033 | 0.004 | <0.001 | <0.001 |
| Brd *Prd | 0.264 | 0.001 | 0.001 | 0.374 | 0.575 |
| Reg *Prd | 0.082 | 0.004 | 0.030 | 0.631 | 0.206 |
| Brd *Reg *Prd | 0.172 | 0.390 | 0.086 | 0.058 | 0.076 |
| Wk | 0.003 | 0.139 | 0.441 | 0.033 | 0.011 |
| Brd *Wk | 0.490 | 0.259 | 0.386 | 0.962 | 0.898 |
| Reg *Wk | 0.511 | 0.171 | 0.149 | 0.296 | 0.058 |
| Brd *Reg *Wk | 0.482 | 0.780 | 0.858 | 0.648 | 0.780 |
| Prd *Wk | <0.001 | 0.026 | 0.270 | 0.396 | 0.655 |
| Brd *Prd *Wk | 0.805 | 0.112 | 0.042 | 0.008 | 0.016 |
| Reg *Prd *Wk | 0.357 | 0.508 | 0.225 | 0.563 | 0.134 |
| Brd* Reg *Prd *Wk | 0.252 | 0.001 | 0.023 | 0.013 | 0.002 |
| T | <0.001 | <0.001 | <0.001 | 0.005 | <0.001 |
| Brd *T | 0.474 | 0.085 | 0.127 | 0.157 | 0.056 |
| Reg *T | 0.399 | 0.500 | 0.974 | 0.108 | 0.148 |
| Brd *Reg *T | 0.917 | 0.905 | 0.624 | 0.114 | 0.195 |
| Prd *T | <0.001 | 0.285 | 0.550 | 0.008 | 0.011 |
| Brd *Prd*T | 0.221 | 0.745 | 0.689 | 0.850 | 0.878 |
| Reg *Prd *T | 0.587 | 0.279 | 0.200 | 0.439 | 0.877 |
| Brd *Reg *Prd *T | 0.765 | 0.603 | 0.418 | 0.227 | 0.359 |
| Wk *T | <0.001 | 0.180 | 0.856 | 0.259 | 0.777 |
| Brd *Wk *T | 0.796 | 0.902 | 0.892 | 0.192 | 0.197 |
| Reg *Wk *T | 0.162 | 0.674 | 0.283 | 0.852 | 0.865 |
| Brd *Reg *Wk *T | 0.465 | 0.860 | 0.505 | 0.247 | 0.372 |
| Prd *Wk *T | <0.001 | <0.001 | 0.031 | 0.003 | 0.083 |
| Brd *Prd *Wk *T | 0.332 | 0.710 | 0.923 | 0.961 | 0.974 |
| Reg *Prd *Wk *T | 0.647 | 0.310 | 0.807 | 0.582 | 0.718 |
| Brd *Reg *Prd *Wk *T | 0.400 | 0.286 | 0.417 | 0.980 | 0.970 |
| Time | Source of Variation 2 | Variable 1 | ||||
|---|---|---|---|---|---|---|
| OSM (mosmol/kg) | PCV (%) | Hb (g/dL) | Hb O2 (%) | O2 (mmol/L) | ||
| 08:00 h | Set | <0.001 | 0.311 | 0.133 | <0.001 | <0.001 |
| Age | 0.794 | 0.684 | 0.813 | 0.659 | 0.661 | |
| Brd | 0.037 | 0.047 | 0.604 | <0.001 | <0.001 | |
| Reg | 0.446 | 0.252 | 0.188 | 0.328 | 0.595 | |
| Brd *Reg | 0.332 | 0.457 | 0.554 | 0.083 | 0.346 | |
| Prd | <0.001 | 0.329 | 0.066 | 0.246 | 0.118 | |
| Brd *Prd | 0.504 | 0.002 | 0.002 | 0.485 | 0.572 | |
| Reg *Prd | 0.070 | 0.006 | 0.025 | 0.530 | 0.497 | |
| Brd *Reg *Prd | 0.137 | 0.354 | 0.166 | 0.121 | 0.380 | |
| Wk | 0.678 | 0.044 | 0.492 | 0.417 | 0.075 | |
| Brd *Wk | 0.620 | 0.340 | 0.567 | 0.405 | 0.568 | |
| Reg *Wk | 0.567 | 0.409 | 0.177 | 0.288 | 0.081 | |
| Brd *Reg *Wk | 0.438 | 0.840 | 0.717 | 0.069 | 0.117 | |
| Prd *Wk | <0.001 | 0.472 | 0.942 | 0.166 | 0.310 | |
| Brd *Prd *Wk | 0.751 | 0.285 | 0.257 | 0.179 | 0.177 | |
| Reg *Prd *Wk | 0.621 | 0.475 | 0.313 | 0.887 | 0.424 | |
| Brd *Reg *Prd *Wk | 0.614 | <0.001 | 0.025 | 0.028 | 0.011 | |
| 14:00 h | Set | <0.001 | 0.200 | 0.172 | <0.001 | <0.001 |
| Age | 0.895 | 0.387 | 0.726 | 0.036 | 0.044 | |
| Brd | 0.022 | 0.260 | 0.679 | 0.026 | 0.021 | |
| Reg | 0.346 | 0.179 | 0.202 | 0.574 | 0.639 | |
| Brd *Reg | 0.391 | 0.352 | 0.567 | 0.156 | 0.270 | |
| Prd | <0.001 | 0.008 | 0.001 | <0.001 | <0.001 | |
| Brd *Prd | 0.158 | 0.009 | 0.009 | 0.520 | 0.812 | |
| Reg *Prd | 0.576 | 0.027 | 0.095 | 0.384 | 0.251 | |
| Brd *Reg *Prd | 0.817 | 0.594 | 0.106 | 0.032 | 0.008 | |
| Wk | <0.001 | 0.887 | 0.626 | 0.013 | 0.019 | |
| Brd *Wk | 0.494 | 0.667 | 0.461 | 0.443 | 0.428 | |
| Reg *Wk | 0.251 | 0.200 | 0.133 | 0.662 | 0.408 | |
| Brd *Reg *Wk | 0.546 | 0.624 | 0.695 | 0.874 | 0.988 | |
| Prd *Wk | <0.001 | <0.001 | 0.014 | 0.023 | 0.352 | |
| Brd *Prd *Wk | 0.030 | 0.302 | 0.078 | 0.051 | 0.074 | |
| Reg *Prd *Wk | <0.001 | 0.370 | 0.597 | 0.422 | 0.476 | |
| Brd *Reg *Prd *Wk | <0.001 | 0.251 | 0.136 | 0.594 | 0.398 | |
3. Results
3.1. OSM, PCV, Hb, and Oxygen
| Variable 1 | ||||||
|---|---|---|---|---|---|---|
| Time | Source of Variation 2 | OSM (mosmol/kg) | PCV (%) | Hb (g/dL) | Hb O2 (%) | O2 (mmol/L) |
| 08:00 h | Set | <0.001 | 0.071 | 0.047 | <0.001 | 0.001 |
| Age | 0.830 | 0.891 | 0.879 | 0.278 | 0.292 | |
| Brd | 0.120 | 0.009 | 0.300 | 0.006 | 0.001 | |
| Reg | 0.568 | 0.501 | 0.338 | 0.453 | 0.694 | |
| Brd *Reg | 0.214 | 0.754 | 0.586 | <0.001 | 0.031 | |
| Prd | 0.001 | 0.604 | 0.098 | 0.623 | 0.250 | |
| Brd *Prd | 0.057 | 0.010 | 0.030 | 0.540 | 0.095 | |
| Reg *Prd | 0.233 | 0.042 | 0.204 | 0.916 | 0.633 | |
| Brd *Reg *Prd | 0.030 | 0.037 | 0.145 | 0.113 | 0.028 | |
| 14:00 h | Set | <0.001 | 0.166 | 0.166 | <0.001 | <0.001 |
| Age | 0.679 | 0.299 | 0.884 | 0.006 | 0.027 | |
| Brd | 0.032 | 0.257 | 0.487 | 0.003 | 0.003 | |
| Reg | 0.391 | 0.102 | 0.155 | 0.999 | 0.459 | |
| Brd *Reg | 0.301 | 0.063 | 0.264 | 0.063 | 0.362 | |
| Prd | 0.339 | 0.008 | <0.001 | 0.002 | <0.001 | |
| Brd *Prd | 0.341 | 0.049 | 0.039 | 0.656 | 0.131 | |
| Reg *Prd | 0.323 | 0.172 | 0.291 | 0.026 | 0.064 | |
| Brd *Reg *Prd | 1.000 | 0.078 | 0.004 | 0.162 | 0.007 | |
| Breed 1 | Region 2 | Period 3 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Item 4 | Time (h) 5 | Brd 6 | Prd 7 | DOR | KAT | STC | SEM | MW | NW | SE | TX | SEM | 1 | 3 | SEM |
| OSM (mosmol/kg) | 14:00 | 301.0 a | 303.2 b | 303.4 b | 0.70 | 301.6 | 302.3 | 302.7 | 303.6 | 0.81 | 302.8 | 302.2 | 0.50 | ||
| PCV (%) | 14:00 | 32.2 | 30.7 | 31.7 | 32.4 | 0.50 | |||||||||
| 1 | 31.4 a | 31.2 a | 31.3 a | 0.52 | |||||||||||
| 3 | 31.7 a | 31.6 a | 33.4 b | ||||||||||||
| Hb (g/dL) | 08:00 | 12.3 | 11.9 | 12.2 | 12.4 | 0.18 | |||||||||
| 1 | 12.1 a | 12.1 a | 12.1 a | 0.20 | |||||||||||
| 3 | 12.1 a | 12.0 a | 12.8 b | ||||||||||||
| Hb O 2 (%) | 08:00 | 69.5 | 70.2 | 1.17 | |||||||||||
| DOR | 66.3 bc | 74.3 cd | 74.2 cd | 57.1 a | 3.1 | ||||||||||
| KAT | 70.4 cd | 60.6 ab | 67.4 bcd | 71.3 cd | |||||||||||
| STC | 71.1 cd | 75.5 d | 74.2 cd | 75.6 d | |||||||||||
| 14:00 | 62.7 a | 65.9 a | 70.3 b | 1.50 | |||||||||||
| 1 | 63.9 ab | 65.5 ab | 60.7 a | 66.3 abc | 2.26 | ||||||||||
| 3 | 68.4 bc | 67.0 bc | 73.2 c | 66.6 abc | |||||||||||
| Dorper1 | Katahdin | St. Croix | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Item 2 | Time (h) 3 | Prd 4 | MW | NW | SE | TX | MW | NW | SE | TX | MW | NW | SE | TX | SEM | ||
| OSM (mosmol/kg) | 08:00 | 1 | 300.5 x | 299.9 | 300.4 | 300.4 | 301.9 a | 299.4 x,a | 299.4 x,a | 307.9 b | 303.2 | 304.7 | 305.1 | 302.9 | 1.80 | ||
| 3 | 305.6 y | 302.7 | 304.5 | 303.5 | 301.4 a | 309.6 y,b | 304.9 y,ab | 305.7 ab | 304.2 | 305.4 | 302.4 | 303.8 | |||||
| PCV (%) | 08:00 | 1 | 31.1 | 32.6 y | 31.2 | 32.8 | 33.2 y | 31.4 | 31.9 | 32.2 | 33.0 | 32.7 | 31.7 x | 33.2 | 1.12 | ||
| 3 | 32.5 ab | 28.5 x,a | 31.7 ab | 33.4 b | 30.3 x | 31.9 | 33.0 | 30.0 | 34.3 | 32.9 | 35.6 y | 35.1 | |||||
| O2 (mmol/L) | 08:00 | 1 | 4.60 a | 5.60 y,b | 4.84 ab | 4.42 a | 4.97 ab | 4.14 a | 5.21 ab | 5.37 b | 5.37 | 5.24 | 5.07 | 5.31 | 0.317 | ||
| 3 | 5.07 ab | 4.65 x,ab | 5.49 b | 4.36 a | 5.34 b | 4.40 a | 4.78 ab | 4.72 ab | 5.25 | 5.79 | 5.84 | 6.09 | |||||
| Hb (g/dL) | 14:00 | 1 | 12.2 | 11.5 | 11.4 x | 12.1 | 11.8 | 11.4 | 12.1 | 11.7 | 12.0 | 11.6 | 11.5 x | 11.5 x | 0.38 | ||
| 3 | 12.7 b | 10.8 a | 12.4 y,b | 12.8 b | 11.9 | 12.1 | 12.0 | 11.4 | 12.2 ab | 12.1 a | 12.3 y,ab | 13.2 y,b | |||||
| O2 (mmol/L) | 14:00 | 1 | 4.33 ab | 5.09 y,b | 3.66 x,a | 4.39 ab | 4.76 b | 3.91 a | 4.53 x,ab | 5.23 b | 4.80 | 4.66 | 4.80 x | 4.69 x | 0.308 | ||
| 3 | 4.92 | 4.16 x | 4.74 y | 4.84 | 5.00 | 4.54 | 5.28 y | 4.42 | 5.15 | 5.21 | 5.65 y | 5.84 y | |||||
3.2. Blood Constituent Concentrations—Wk 2, 4, 6, 8, and 9
3.3. Blood Constituent Concentrations—Wk 2 and 9
4. Discussion
4.1. OSM, PCV, Hb, and Oxygen
4.2. Blood Constituent Concentrations—Wk 2, 4, 6, 8, and 9
4.2.1. Period
4.2.2. Breed
4.3. Blood Constituent Concentrations—Wk 2 and 9
5. Conclusions and Implications
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aganga, A.A.; Umunna, N.N.; Oyedipe, E.O.; Okoh, P.N. Influence of Water Restriction on Some Serum Components in Yankasa Ewes. Small Rumin. Res. 1989, 2, 19–26. [Google Scholar] [CrossRef]
- Hamadeh, S.K.; Rawda, N.; Jaber, L.S.; Habre, A.; Abi Said, M.; Barbour, E.K. Physiological Responses to Water Restriction in Dry and Lactating Awassi Ewes. Livest. Sci. 2006, 101, 101–109. [Google Scholar] [CrossRef]
- Kumar, D.; De, K.; Singh, A.K.; Kumar, K.; Sahoo, A.; Naqvi, S.M.K. Effect of Water Restriction on Physiological Responses and Certain Reproductive Traits of Malpura Ewes in a Semiarid Tropical Environment. J. Vet. Behav. 2016, 12, 54–59. [Google Scholar] [CrossRef]
- Alamer, M. Physiological Responses of Saudi Arabia Indigenous Goats to Water Deprivation. Small Rumin. Res. 2006, 63, 100–109. [Google Scholar] [CrossRef]
- Kaliber, M.; Koluman, N.; Silanikove, N. Physiological and Behavioral Basis for the Successful Adaptation of Goats to Severe Water Restriction under Hot Environmental Conditions. Animal 2016, 10, 82–88. [Google Scholar] [CrossRef]
- Mengistu, U.; Dahlborn, K.; Olsson, K. Mechanisms of Water Economy in Lactating Ethiopian Somali Goats during Repeated Cycles of Intermittent Watering. Animal 2007, 1, 1009–1017. [Google Scholar] [CrossRef]
- Ghanem, A.M.; Jaber, L.S.; Abi Said, M.; Barbour, E.K.; Hamadeh, S.K. Physiological and Chemical Responses in Water-Deprived Awassi Ewes Treated with Vitamin C. J. Arid Environ. 2008, 72, 141–149. [Google Scholar] [CrossRef]
- Jaber, L.S.; Habre, A.; Rawda, N.; Abi Said, M.; Barbour, E.K.; Hamadeh, S. The Effect of Water Restriction on Certain Physiological Parameters in Awassi Sheep. Small Rumin. Res. 2004, 54, 115–120. [Google Scholar] [CrossRef]
- Igbokwe, I.O. Haemoconcentration in Yankasa Sheep Exposed to Prolonged Water Deprivation. Small Rumin. Res. 1993, 12, 99–105. [Google Scholar] [CrossRef]
- Mengistu, U.; Dahlborn, K.; Olsson, K. Effects of Intermittent Watering on Water Balance and Feed Intake in Male Ethiopian Somali Goats. Small Rumin. Res. 2007, 67, 45–54. [Google Scholar] [CrossRef]
- De, K.; Kumar, D.; Singh, A.K.; Kumar, K.; Sahoo, A.; Naqvi, S.M.K. Resilience of Malpura Ewes on Water Restriction and Rehydration during Summer under Semi-Arid Tropical Climatic Conditions. Small Rumin. Res. 2015, 133, 123–127. [Google Scholar] [CrossRef]
- Vosooghi-Postindoz, V.; Tahmasbi, A.; Naserian, A.A.; Valizade, R.; Ebrahimi, H. Effect of Water Deprivation and Drinking Saline Water on Performance, Blood Metabolites, Nutrient Digestibility, and Rumen Parameters in Baluchi Lambs. Iran. J. Appl. Anim. Sci. 2018, 8, 445–456. [Google Scholar]
- Ahmed, M.M.; El Kheir, I.M. Thermoregulation and Water Balance as Affected by Water and Food Restrictions in Sudanese Desert Goats Fed Good-Quality and Poor-Quality Diets. Trop. Anim. Health Prod. 2004, 36, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Alamer, M. Effect of Water Restriction on Lactation Performance of Aardi Goats under Heat Stress Conditions. Small Rumin. Res. 2009, 84, 76–81. [Google Scholar] [CrossRef]
- Aleena, J.; Sejian, V.; Krishnan, G.; Bagath, M.; Pragna, P.; Bhatta, R. Heat Stress Impact on Blood Biochemical Response and Plasma Aldosterone Level in Three Different Indigenous Goat Breeds. J. Anim. Behav. Biometeorol. 2020, 8, 266–275. [Google Scholar] [CrossRef]
- Kataria, N.; Kataria, A.K. Compartmental Water Management of Marwari Sheep during Dehydration and Rehydration. Vet. Arh. 2007, 77, 551. [Google Scholar]
- D’Ambrosio, C.; Sarubbi, F.; Scaloni, A.; Rossetti, C.; Grazioli, G.; Auriemma, G.; Perucatti, A.; Spagnuolo, M.S. Effect of Short-Term Water Restriction on Oxidative and Inflammatory Status of Sheep (Ovis Aries) Reared in Southern Italy. Small Rumin. Res. 2018, 162, 77–84. [Google Scholar] [CrossRef]
- Olsson, K.; Dahlborn, K. Fluid Balance during Heat Stress in Lactating Goats. Q. J. Exp. Physiol. Transl. Integr. 1989, 74, 645–659. [Google Scholar] [CrossRef]
- Wittenberg, C.; Choshniak, I.; Shkolnik, A.; Thurau, K.; Rosenfeld, J. Effect of Dehydration and Rapid Rehydration on Renal Function and on Plasma Renin and Aldosterone Levels in the Black Bedouin Goat. Pflüg. Arch. Eur. J. Physiol. 1986, 406, 405–408. [Google Scholar] [CrossRef]
- Casamassima, D.; Pizzo, R.; Palazzo, M.; D’alessandro, A.G.; Martemucci, G. Effect of Water Restriction on Productive Performance and Blood Parameters in Comisana Sheep Reared under Intensive Condition. Small Rumin. Res. 2008, 78, 169–175. [Google Scholar] [CrossRef]
- Hussein, A.; Puchala, R.; Portugal, I.; Wilson, B.K.; Gipson, T.A.; Goetsch, A.L. Effects of Restricted Availability of Drinking Water on Body Weight and Feed Intake by Dorper, Katahdin, and St. Croix Sheep from Different Regions of the USA. J. Anim. Sci. 2020, 98, skz367. [Google Scholar] [CrossRef] [PubMed]
- Kataria, N.; Kataria, A.K. Endocrine and Metabolic Responses of Marwari Sheep in Arid Tract. Slov. Vet. Res. 2006, 43, 135–142. [Google Scholar]
- Laden, S.; Nehmadi, L.; Yagil, R. Dehydration Tolerance in Awassi Fat-Tailed Sheep. Can. J. Zool. 1987, 65, 363–367. [Google Scholar] [CrossRef]
- Marques, R.S.; Bohnert, D.W.; De Sousa, O.A.; Brandão, A.P.; Schumaher, T.F.; Schubach, K.M.; Vilela, M.P.; Rett, B.; Cooke, R.F. Impact of 24-h Feed, Water, or Feed and Water Deprivation on Feed Intake, Metabolic, and Inflammatory Responses in Beef Heifers. J. Anim. Sci. 2019, 97, 398–406. [Google Scholar] [CrossRef]
- Silanikove, N. The Physiological Basis of Adaptation in Goats to Harsh Environments. Small Rumin. Res. 2000, 35, 181–193. [Google Scholar] [CrossRef]
- Hussein, A.H.; Puchala, R.; Gipson, T.A.; Tadesse, D.; Wilson, B.K.; Goetsch, A.L. Effects of Water Restriction on Feed Intake, Digestion, and Energy Utilization by Mature Female St. Croix Sheep. Vet. Anim. Sci. 2020, 10, 100132. [Google Scholar] [CrossRef]
- Tadesse, D.; Patra, A.K.; Puchala, R.; Goetsch, A.L. Effects of High Heat Load Conditions on Blood Constituent Concentrations in Dorper, Katahdin, and St. Croix Sheep from Different Regions of the USA. Animals 2022, 12, 2273. [Google Scholar] [CrossRef] [PubMed]
- Tadesse, D.; Puchala, R.; Goetsch, A.L. Effects of Restricted Feed Intake on Blood Constituent Concentrations in Dorper, Katahdin, and St. Croix Sheep from Different Regions of the USA. Vet. Anim. Sci. 2021, 14, 100211. [Google Scholar] [CrossRef]
- Tadesse, D.; Puchala, R.; Gipson, T.A.; Goetsch, A.L. Effects of High Heat Load Conditions on Body Weight, Feed Intake, Temperature, and Respiration of Dorper, Katahdin, and St. Croix Sheep. J. Appl. Anim. Res. 2019, 47, 492–505. [Google Scholar] [CrossRef]
- Tadesse, D.; Puchala, R.; Goetsch, A.L. Effects of Hair Sheep Breed and Region of Origin on Feed Dry Matter Required for Maintenance without and with a Marked Feed Restriction. Livest. Sci. 2019, 226, 114–121. [Google Scholar] [CrossRef]
- Kampe, T.U.; Johnson, B.R.; Kuester, M.A.; Keller, M. NEON: The First Continental-Scale Ecological Observatory with Airborne Remote Sensing of Vegetation Canopy Biochemistry and Structure. J. Appl. Remote Sens. 2010, 4, 043510. [Google Scholar] [CrossRef]
- Fovell, R.G.; Fovell, M.-Y.C. Climate Zones of the Conterminous United States Defined Using Cluster Analysis. J. Clim. 1993, 6, 2103–2135. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids; National Academy Press: Washington DC, USA, 2007. [Google Scholar]
- Mengistu, U.L.; Puchala, R.; Sahlu, T.; Gipson, T.A.; Dawson, L.J.; Goetsch, A.L. Comparison of Different Levels and Lengths of Restricted Drinking Water Availability and Measurement Times with Katahdin Sheep and Boer and Spanish Goat Wethers. Small Rumin. Res. 2016, 144, 320–333. [Google Scholar] [CrossRef]
- Eisemann, J.H.; Nienaber, J.A. Tissue and Whole-Body Oxygen Uptake in Fed and Fasted Steers. Br. J. Nutr. 1990, 64, 399–411. [Google Scholar] [CrossRef]
- Oladele, S.B.; Morou, M.; Sambo, S.J.; Ibu, O.J. Comparative Studies of Packed Cell Volume, Haemoglobin, Total Protein, Haemagglutination Inhibition Antibodies and Rectal Temperature of Pigeons (Columbia Livia) Administered Newcastle Disease Virus through Different Routes. Int. J. Poult. Sci. 2008, 7, 898–902. [Google Scholar] [CrossRef][Green Version]
- Littell, R.C.; Henry, P.R.; Ammerman, C.B. Statistical Analysis of Repeated Measures Data Using SAS Procedures. J. Anim. Sci. 1998, 76, 1216–1231. [Google Scholar] [CrossRef] [PubMed]
- SAS. SAS/STAT User’s Guide; Version 9.3; SAS Institute Inc.: Cary, NC, USA, 2011. [Google Scholar]
- Silanikove, N. The Struggle to Maintain Hydration and Osmoregulation in Animals Experiencing Severe Dehydration and Rapid Rehydration: The Story of Ruminants. Exp. Physiol. Transl. Integr. 1994, 79, 281–300. [Google Scholar] [CrossRef]
- Schaefer, A.L.; Jones, S.D.M.; Tong, A.K.W.; Lepage, P.; Murray, N.L. The Effects of Withholding Feed and Water on Selective Blood Metabolites in Market-Weight Beef Steers. Can. J. Anim. Sci. 1990, 70, 1155–1158. [Google Scholar] [CrossRef]
- Degen, A.A.; Kam, M. Body Mass Loss and Body Fluid Shifts during Dehydration in Dorper Sheep. J. Agric. Sci. 1992, 119, 419–422. [Google Scholar] [CrossRef]
- Martín-Calderón, J.L.; Bustos, F.; Tuesta-Reina, L.R.; Varona, J.M.; Caballero, L.; Solano, F. Choice of the Best Equation for Plasma Osmolality Calculation: Comparison of Fourteen Formulae. Clin. Biochem. 2015, 48, 529–533. [Google Scholar] [CrossRef]
- Chen, J.S.; Sabir, S.; Al Khalili, Y. Physiology, Osmoregulation and Excretion. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Gäbel, G.; Martens, H.; Sündermann, M.; Galfi, P. The Effect of Diet, Intraruminal PH and Osmolarity on Sodium, Chloride and Magnesium Absorption from the Temporarily Isolated and Washed Reticulo-Rumen of Sheep. Q. J. Exp. Physiol. Transl. Integr. 1987, 72, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Carter, R.R.; Grovum, W.L. A Review of the Physiological Significance of Hypertonic Body Fluids on Feed Intake and Ruminal Function: Salivation, Motility and Microbes. J. Anim. Sci. 1990, 68, 2811–2832. [Google Scholar] [CrossRef] [PubMed]
- Silanikove, N.; Tagari, H.; Shkolnik, A. Comparison of Rate of Passage, Fermentation Rate and Efficiency of Digestion of High Fiber Diet in Desert Bedouin Goats Compared to Swiss Saanen Goats. Small Rumin. Res. 1993, 12, 45–60. [Google Scholar] [CrossRef]
- Ahmed, M.M.; Abdelatif, A.M. Effects of Restriction of Water and Food Intake on Thermoregulation, Food Utilization and Water Economy in Desert Sheep. J. Arid Environ. 1994, 28, 147–153. [Google Scholar] [CrossRef]
- Leow, M.K.-S. Configuration of the Hemoglobin Oxygen Dissociation Curve Demystified: A Basic Mathematical Proof for Medical and Biological Sciences Undergraduates. Adv. Physiol. Educ. 2007, 31, 198–201. [Google Scholar] [CrossRef]
- Annison, E.F.; White, R.R. Glucose Utilization in Sheep. Biochem. J. 1961, 80, 162. [Google Scholar] [CrossRef] [PubMed]
- Baird, G.D.; Lomax, M.A.; Symonds, H.W.; Shaw, S.R. Net Hepatic and Splanchnic Metabolism of Lactate, Pyruvate and Propionate in Dairy Cows in Vivo in Relation to Lactation and Nutrient Supply. Biochem. J. 1980, 186, 47–57. [Google Scholar] [CrossRef]
- Allen, M.S.; Bradford, B.J.; Oba, M. Board-Invited Review: The Hepatic Oxidation Theory of the Control of Feed Intake and Its Application to Ruminants. J. Anim. Sci. 2009, 87, 3317–3334. [Google Scholar] [CrossRef]
- Woerle, H.J.; Meyer, C.; Dostou, J.M.; Gosmanov, N.R.; Islam, N.; Popa, E.; Wittlin, S.D.; Welle, S.L.; Gerich, J.E. Pathways for Glucose Disposal after Meal Ingestion in Humans. Am. J. Physiol.-Endocrinol. Metab. 2003, 284, E716–E725. [Google Scholar] [CrossRef]
- Ewaschuk, J.B.; Naylor, J.M.; Zello, G.A. D-Lactate in Human and Ruminant Metabolism. J. Nutr. 2005, 135, 1619–1625. [Google Scholar] [CrossRef]
- Huntington, G.B.; Archibeque, S.L. Practical Aspects of Urea and Ammonia Metabolism in Ruminants. J. Anim. Sci. 2000, 77 (Suppl. E), 1–11. [Google Scholar] [CrossRef]
- Caldeira, R.M.; Belo, A.T.; Santos, C.C.; Vazques, M.I.; Portugal, A.V. The Effect of Body Condition Score on Blood Metabolites and Hormonal Profiles in Ewes. Small Rumin. Res. 2007, 68, 233–241. [Google Scholar] [CrossRef]
- Marini, J.C.; Klein, J.D.; Sands, J.M.; Van Amburgh, M.E. Effect of Nitrogen Intake on Nitrogen Recycling and Urea Transporter Abundance in Lambs. J. Anim. Sci. 2004, 82, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Weiner, I.D.; Mitch, W.E.; Sands, J.M. Urea and Ammonia Metabolism and the Control of Renal Nitrogen Excretion. Clin. J. Am. Soc. Nephrol. 2015, 10, 1444–1458. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.D.; Blount, M.A.; Sands, J.M. Urea Transport in the Kidney. Compr. Physiol. 2011, 1, 699–729. [Google Scholar]
- Hosten, A.O. BUN and Creatinine. In Clinical Methods: The History, Physical, and Laboratory Examinations, 3rd ed.; Walker, H.K., Hall, W.D., Hurst, J.W., Eds.; Butterworths: Boston, MA, USA, 1990; pp. 874–878. [Google Scholar]
- Sjöström, P.A.; Odlind, B.G.; Wolgast, M. Extensive Tubular Secretion and Reabsorption of Creatinine in Humans. Scand. J. Urol. Nephrol. 1988, 22, 129–131. [Google Scholar] [CrossRef]
- Caldeira, R.M.; Belo, A.T.; Santos, C.C.; Vazques, M.I.; Portugal, A.V. The Effect of Long-Term Feed Restriction and over-Nutrition on Body Condition Score, Blood Metabolites and Hormonal Profiles in Ewes. Small Rumin. Res. 2007, 68, 242–255. [Google Scholar] [CrossRef]
- Moorby, J.M.; Dewhurst, R.J.; Evans, R.T.; Fisher, W.J. Effects of Level of Concentrate Feeding during the Second Gestation of Holstein-Friesian Dairy Cows. 2. Nitrogen Balance and Plasma Metabolites. J. Dairy Sci. 2002, 85, 178–189. [Google Scholar] [CrossRef]
- Burton, R.F. The Protein Content of Extracellular Fluids and Its Relevance to the Study of Ionic Regulation: Net Charge and Colloid Osmotic Pressure. Comp. Biochem. Physiol. A 1988, 90, 11–16. [Google Scholar] [CrossRef]
- El-Sherif, M.M.A.; Assad, F. Changes in Some Blood Constituents of Barki Ewes during Pregnancy and Lactation under Semi Arid Conditions. Small Rumin. Res. 2001, 40, 269–277. [Google Scholar] [CrossRef]
- Umunna, N.N.; Chineme, C.N.; Saror, D.I.; Ahmed, A.; Abed, S. Response of Yankasa Sheep to Various Lengths of Water Deprivation. J. Agric. Sci. 1981, 96, 619–622. [Google Scholar] [CrossRef]
- Nejad, J.G.; Lohakare, J.D.; Son, J.K.; Kwon, E.G.; West, J.W.; Sung, K.I. Wool Cortisol Is a Better Indicator of Stress than Blood Cortisol in Ewes Exposed to Heat Stress and Water Restriction. Animal 2014, 8, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Chedid, M.; Jaber, L.S.; Giger-Reverdin, S.; Duvaux-Ponter, C.; Hamadeh, S.K. Water Stress in Sheep Raised under Arid Conditions. Can. J. Anim. Sci. 2014, 94, 243–257. [Google Scholar] [CrossRef]
- Rizos, C.V.; Elisaf, M.S.; Liberopoulos, E.N. Effects of Thyroid Dysfunction on Lipid Profile. Open Cardiovasc. Med. J. 2011, 5, 76. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Kar, I. Heat Stress on Microbiota Composition, Barrier Integrity, and Nutrient Transport in Gut, Production Performance, and Its Amelioration in Farm Animals. J. Anim. Sci. Technol. 2021, 63, 211–247. [Google Scholar] [CrossRef] [PubMed]

| Dorper3 | Katahdin | St. Croix | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variable 4 | Prd 5 | Wk 5 | MW | NW | SE | TX | MW | NW | SE | TX | MW | NW | SE | TX | SEM | ||
| PCV (%) | 1 | 1 | 35.3 | 31.8 | 31.8 | 32.7 | 32.0 | 32.5 | 33.0 | 29.9 | 33.2 | 32.7 | 31.0 | 32.7 | 1.08 | ||
| 1 | 2 | 31.2 | 32.7 | 31.0 | 32.9 | 33.4 | 31.8 | 32.0 | 32.0 | 33.0 | 32.6 | 31.4 | 33.1 | ||||
| 2 | 3 | 33.2 | 30.9 | 30.4 | 33.8 | 33.1 | 30.7 | 33.2 | 32.7 | 33.9 | 33.3 | 32.7 | 35.7 | ||||
| 2 | 4 | 33.7 | 29.4 | 30.6 | 33.6 | 31.7 | 31.2 | 32.5 | 33.1 | 33.2 | 33.3 | 32.5 | 34.1 | ||||
| 3 | 8 | 32.2 | 29.2 | 33.0 | 33.8 | 33.0 | 32.1 | 32.5 | 32.7 | 34.1 | 34.7 | 34.4 | 34.5 | ||||
| 3 | 9 | 32.5 | 28.6 | 31.5 | 33.5 | 30.5 | 32.0 | 33.2 | 29.8 | 34.4 | 32.7 | 35.3 | 35.1 | ||||
| Hb (g/dL) | 1 | 1 | 13.2 | 12.1 | 12.1 | 12.2 | 12.2 | 12.4 | 12.4 | 11.4 | 12.3 | 12.2 | 11.3 | 11.9 | 0.42 | ||
| 1 | 2 | 12.0 | 12.1 | 11.7 | 12.6 | 12.5 | 12.0 | 12.2 | 12.2 | 12.2 | 12.0 | 11.6 | 11.3 | ||||
| 2 | 3 | 12.6 | 11.8 | 11.8 | 12.6 | 12.7 | 11.7 | 12.7 | 12.4 | 12.6 | 12.3 | 12.2 | 12.4 | ||||
| 2 | 4 | 12.7 | 11.6 | 11.6 | 12.5 | 12.5 | 11.7 | 12.4 | 12.7 | 12.6 | 12.5 | 12.1 | 12.6 | ||||
| 3 | 8 | 12.5 | 10.6 | 12.5 | 12.7 | 12.6 | 12.0 | 12.2 | 12.3 | 12.8 | 12.8 | 12.8 | 12.7 | ||||
| 3 | 9 | 12.5 | 11.1 | 12.1 | 12.9 | 12.0 | 12.2 | 12.5 | 11.4 | 12.7 | 12.4 | 12.9 | 13.0 | ||||
| Hb O2 (%) | 1 | 1 | 72.0 | 69.7 | 59.7 | 59.2 | 64.8 | 65.1 | 71.4 | 66.1 | 75.8 | 79.5 | 78.4 | 67.3 | 4.24 | ||
| 1 | 2 | 64.6 | 77.7 | 70.6 | 59.5 | 66.5 | 58.9 | 73.2 | 74.5 | 73.6 | 72.0 | 73.0 | 71.2 | ||||
| 2 | 3 | 65.1 | 75.1 | 67.4 | 57.6 | 66.4 | 74.3 | 62.4 | 65.4 | 76.8 | 74.0 | 74.9 | 74.9 | ||||
| 2 | 4 | 65.8 | 71.2 | 64.7 | 64.9 | 65.3 | 60.8 | 66.0 | 54.9 | 71.3 | 73.9 | 70.5 | 73.7 | ||||
| 3 | 8 | 70.8 | 74.1 | 67.7 | 64.8 | 68.2 | 66.0 | 71.9 | 70.1 | 73.0 | 70.4 | 71.5 | 72.6 | ||||
| 3 | 9 | 68.4 | 70.9 | 77.5 | 57.2 | 74.1 | 58.8 | 64.9 | 70.9 | 69.9 | 77.8 | 76.1 | 78.3 | ||||
| O2 (mmol/L) | 1 | 1 | 5.66 | 4.96 | 4.42 | 4.38 | 4.87 | 4.82 | 5.31 | 4.51 | 5.58 | 5.94 | 5.27 | 4.82 | 0.317 | ||
| 1 | 2 | 4.60 | 5.63 | 4.82 | 4.51 | 5.00 | 4.06 | 5.36 | 5.45 | 5.40 | 5.18 | 5.04 | 5.27 | ||||
| 2 | 3 | 4.91 | 5.27 | 4.78 | 4.33 | 5.00 | 5.18 | 4.82 | 4.96 | 5.80 | 5.45 | 5.49 | 5.89 | ||||
| 2 | 4 | 5.00 | 4.96 | 4.55 | 4.91 | 4.91 | 4.24 | 4.91 | 4.24 | 5.31 | 5.49 | 5.09 | 5.49 | ||||
| 3 | 8 | 5.27 | 5.09 | 5.00 | 4.91 | 5.13 | 4.73 | 5.27 | 5.13 | 5.58 | 5.54 | 5.49 | 5.54 | ||||
| 3 | 9 | 5.09 | 4.64 | 5.45 | 4.46 | 5.36 | 4.33 | 4.91 | 4.78 | 5.31 | 5.71 | 5.85 | 5.03 | ||||
| Period 1 | Period 2 | Period 3 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Item | Time (h) | Wk 1 | Wk 2 | Wk 3 | Wk 4 | Wk 8 | Wk 9 | SEM | ||
| OSM (mosmol/kg) | 08:00 | 298.3 a | 302.1 b | 304.7 c | 301.6 b | 305.8 c | 304.4 c | 0.63 | ||
| 14:00 | 303.0 a | 302.9 a | 307.0 bc | 305.4 b | 307.3 c | 302.3 a | 0.65 | |||
| PCV (%) | 14:00 | 31.0 a | 31.3 ab | 32.0 cd | 31.1 ab | 31.7 bc | 32.2 d | 0.30 | ||
| Hb (g/dL) | 14:00 | 11.8 ab | 11.7 a | 12.1 cd | 11.9 abc | 12.0 bc | 12.2 d | 0.11 | ||
| Hb O2 (%) | 14:00 | 67.3 bc | 63.9 a | 65.2 ab | 66.6 abc | 73.8 d | 68.4 c | 1.33 | ||
| Source of Variation 2 | Variable 1 | |||||||
|---|---|---|---|---|---|---|---|---|
| GLC (mg/dL) | LAC (mg/dL) | UN (mg/dL) | TP (g/dL) | ALB (g/dL) | CRT (mg/dL) | CHL (mg/dL) | TG (mg/dL) | |
| Set | 0.030 | <0.001 | 0.024 | <0.001 | <0.001 | 0.068 | 0.001 | <0.001 |
| Age | 0.164 | 0.199 | 0.100 | 0.114 | 0.008 | 0.219 | 0.878 | 0.731 |
| Brd | 0.018 | 0.638 | 0.010 | 0.368 | 0.920 | <0.001 | 0.235 | 0.001 |
| Reg | 0.980 | 0.219 | 0.481 | 0.162 | 0.473 | 0.413 | 0.550 | 0.656 |
| Brd *Reg | 0.612 | 0.818 | 0.509 | 0.072 | 0.256 | 0.025 | 0.523 | 0.177 |
| Wk 3 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Brd *Wk | 0.894 | 0.696 | 0.753 | 0.910 | 0.756 | 0.774 | 0.659 | 0.745 |
| Reg *Wk | 0.051 | 0.183 | 0.199 | 0.994 | 0.951 | 0.564 | 0.553 | 0.503 |
| Brd *Reg *Wk | 0.228 | 0.187 | 0.659 | 0.876 | 0.773 | 0.134 | 0.894 | 0.854 |
| Breed 1 | Region 2 | Week 3 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Item 4 | Brd 5 | DOR | KAT | STC | SEM | MW | NW | SE | TX | SEM | 2 | 4 | 6 | 8 | 9 | SEM |
| GLC | 53.7 a | 55.4 ab | 57.6 b | 0.94 | 55.6 | 55.5 | 56.0 | 55.3 | 1.08 | 55.5 b | 51.0 a | 59.0 c | 56.2 b | 56.3 b | 0.91 | |
| LAC | 22.0 | 21.9 | 22.8 | 0.68 | 22.1 | 23.5 | 22.2 | 21.2 | 0.77 | 24.0 c | 20.5 a | 21.6 ab | 22.8 bc | 22.3 abc | 0.73 | |
| UN | 20.6 a | 20.7 a | 22.4 b | 0.44 | 21.1 | 21.0 | 21.9 | 20.9 | 0.50 | 19.4 a | 20.8 b | 22.5 c | 22.5 c | 21.0 b | 0.38 | |
| TP | 7.07 | 7.07 | 7.22 | 0.084 | 7.05 | 7.23 | 7.23 | 6.98 | 0.098 | 7.08 b | 6.62 a | 7.29 bc | 7.43 c | 7.19 b | 0.084 | |
| ALB | 2.63 | 2.62 | 2.61 | 0.029 | 2.62 | 2.64 | 2.65 | 2.58 | 0.033 | 2.59 b | 2.43 a | 2.68 cd | 2.74 d | 2.65 bc | 0.029 | |
| CRT | 0.840 ab | 0.815 a | 0.959 d | 0.919 c | 0.864 b | 0.0131 | ||||||||||
| DOR | 0.970 c | 1.070 d | 0.944 c | 0.939 c | 0.0331 | |||||||||||
| KAT | 0.860 bc | 0.915 c | 0.907 c | 0.845 b | ||||||||||||
| STC | 0.773 ab | 0.730a | 0.787ab | 0.811ab | ||||||||||||
| CHL | 64.7 | 67.3 | 63.1 | 1.72 | 66.9 | 62.8 | 65.2 | 65.4 | 1.99 | 56.2 a | 57.3 a | 69.5 b | 72.9 c | 69.3 b | 1.33 | |
| TG | 32.3 b | 35.1 b | 28.3 a | 1.24 | 31.7 | 33.4 | 31.7 | 30.9 | 1.44 | 28.6 a | 28.3 a | 33.7 b | 34.5 b | 34.5 b | 0.98 | |
| Item 2 | Source of Variation 1 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Set | Age | Brd | Reg | Brd *Reg | Prd2 | Brd *Prd | Reg *Prd | Brd *Reg *Prd | |
| GLC (mg/dL) | 0.010 | 0.512 | 0.032 | 0.937 | 0.698 | 0.511 | 0.209 | 0.931 | 0.873 |
| LAC (mg/dL) | <0.001 | 0.772 | 0.795 | 0.107 | 0.946 | 0.153 | 0.259 | 0.261 | 0.098 |
| UN (mg/dL) | 0.085 | 0.174 | 0.001 | 0.727 | 0.683 | 0.001 | 0.981 | 0.059 | 0.324 |
| TP (g/L) | 0.368 | 0.037 | 0.243 | 0.085 | 0.225 | 0.357 | 0.613 | 0.851 | 0.928 |
| ALB (g/L) | 0.017 | 0.078 | 0.929 | 0.516 | 0.348 | 0.090 | 0.556 | 0.813 | 0.946 |
| CRT (mg/dL) | 0.385 | 0.338 | <0.001 | 0.363 | 0.020 | 0.070 | 0.297 | 0.134 | 0.004 |
| CHL (mg/dL) | 0.005 | 0.860 | 0.371 | 0.965 | 0.817 | <0.001 | 0.469 | 0.325 | 0.714 |
| TG (mg/dL) | <0.001 | 0.575 | <0.001 | 0.179 | 0.153 | <0.001 | 0.222 | 0.777 | 0.883 |
| COR (ng/mL) | 0.630 | 0.699 | 0.882 | 0.744 | 0.867 | 0.324 | 0.223 | 0.513 | 0.334 |
| ALD (pg/mL) | 0.150 | 0.831 | 0.008 | 0.881 | 0.252 | 0.312 | 0.524 | 0.274 | 0.185 |
| Breed 1 | Region 2 | Prd 3 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Item 4 | Brd 5 | Prd | DOR | KAT | STC | SEM | MW | NW | SE | TX | SEM | 1 | 3 | SEM |
| GLC | 53.8 a | 55.3 ab | 58.0 b | 1.10 | 56.3 | 55.5 | 55.7 | 55.2 | 1.26 | 55.3 | 56.1 | 0.84 | ||
| LAC | 22.9 | 23.3 | 23.8 | 0.98 | 22.5 | 25.6 | 23.4 | 21.8 | 1.11 | 24.1 | 22.5 | 0.79 | ||
| UN | 19.2 a | 19.7 a | 21.6 b | 0.46 | 19.8 | 20.1 | 20.6 | 20.1 | 0.53 | 19.4 a | 21.0 b | 0.34 | ||
| TP | 7.04 | 7.06 | 7.27 | 0.102 | 6.94 | 7.32 | 7.23 | 7.01 | 0.117 | 7.08 | 7.17 | 0.078 | ||
| ALB | 2.63 | 2.61 | 2.62 | 0.039 | 2.58 | 2.65 | 2.66 | 2.59 | 0.044 | 2.59 | 2.65 | 0.029 | ||
| CRT | DOR | 1 | 0.991y | 0.961 x | 0.901 | 0.890 | 0.0421 | |||||||
| DOR | 3 | 0.882x | 1.118 y | 0.889 | 0.900 | |||||||||
| KAT | 1 | 0.761 x,a | 0.865 ab | 0.879 b | 0.826 ab | |||||||||
| KAT | 3 | 0.914 y,b | 0.941 b | 0.896 ab | 0.801 a | |||||||||
| STC | 1 | 0.761 | 0.699 | 0.777 | 0.756 | |||||||||
| STC | 3 | 0.742 | 0.704 | 0.764 | 0.818 | |||||||||
| CHL | 64.7 | 67.3 | 63.1 | 1.72 | 66.9 | 62.8 | 65.2 | 65.4 | 1.99 | 56.2 a | 69.3 b | 1.33 | ||
| TG | 32.3 b | 35.1 b | 28.3 a | 1.24 | 31.7 | 33.4 | 31.7 | 30.9 | 1.44 | 28.6 a | 34.5 b | 0.98 | ||
| COR | 6.43 | 6.98 | 6.65 | 0.787 | 5.94 | 7.31 | 6.53 | 6.97 | 0.907 | 7.02 | 6.36 | 0.560 | ||
| ALD | 52.0 a | 62.5 a | 108.4 b | 13.07 | 78.8 | 80.8 | 65.5 | 72.0 | 15.08 | 78.2 | 70.3 | 8.45 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussein, A.H.; Patra, A.K.; Puchala, R.; Wilson, B.K.; Goetsch, A.L. Effects of Restricted Availability of Drinking Water on Blood Characteristics and Constituents in Dorper, Katahdin, and St. Croix Sheep from Different Regions of the USA. Animals 2022, 12, 3167. https://doi.org/10.3390/ani12223167
Hussein AH, Patra AK, Puchala R, Wilson BK, Goetsch AL. Effects of Restricted Availability of Drinking Water on Blood Characteristics and Constituents in Dorper, Katahdin, and St. Croix Sheep from Different Regions of the USA. Animals. 2022; 12(22):3167. https://doi.org/10.3390/ani12223167
Chicago/Turabian StyleHussein, Ali Hussein, Amlan Kumar Patra, Ryszard Puchala, Blake Kenyon Wilson, and Arthur Louis Goetsch. 2022. "Effects of Restricted Availability of Drinking Water on Blood Characteristics and Constituents in Dorper, Katahdin, and St. Croix Sheep from Different Regions of the USA" Animals 12, no. 22: 3167. https://doi.org/10.3390/ani12223167
APA StyleHussein, A. H., Patra, A. K., Puchala, R., Wilson, B. K., & Goetsch, A. L. (2022). Effects of Restricted Availability of Drinking Water on Blood Characteristics and Constituents in Dorper, Katahdin, and St. Croix Sheep from Different Regions of the USA. Animals, 12(22), 3167. https://doi.org/10.3390/ani12223167