
Island Hopping through Urban Filters: Anthropogenic Habitats and Colonized Landscapes Alter Morphological and Performance Traits of an Invasive Amphibian
 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Study Species
2.2. Study Sites
2.3. Data Collection
2.4. Statistical Analyses
2.4.1. Morphology
2.4.2. Hopping Ability
2.4.3. Climbing Ability
3. Results
3.1. Morphology
3.2. Hopping Ability
3.3. Climbing Ability
4. Discussion
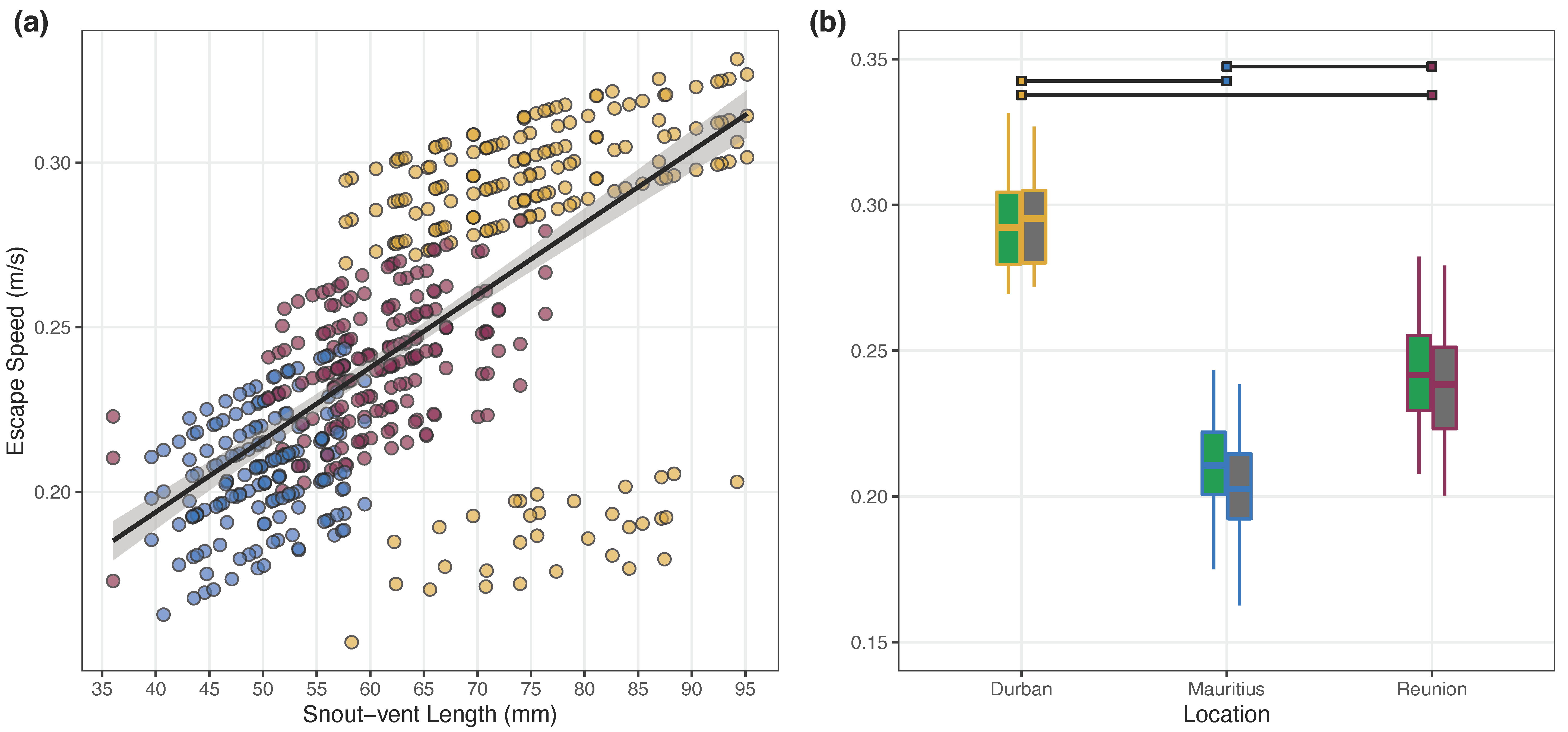
4.1. Escape Speed
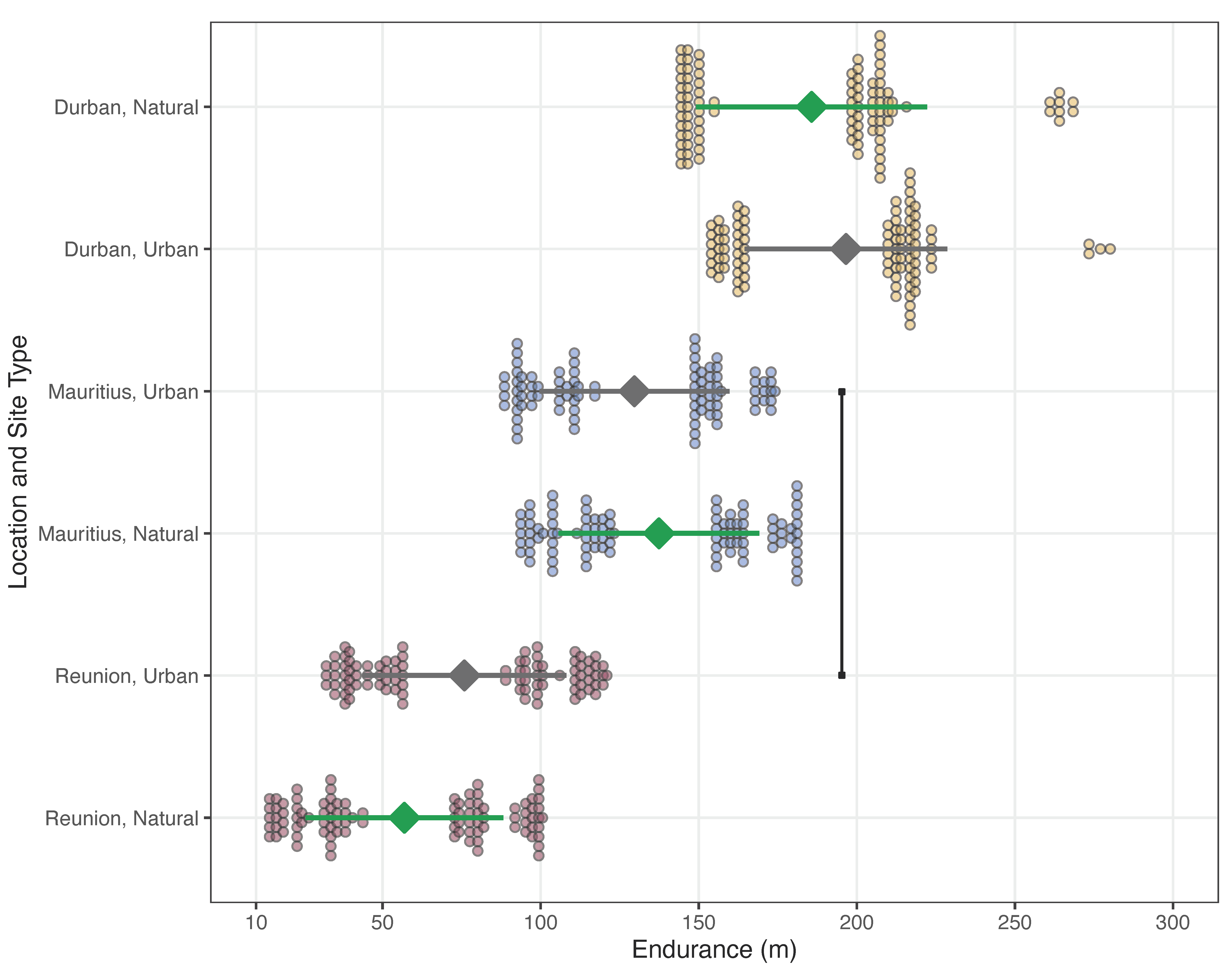
4.2. Endurance Capacity
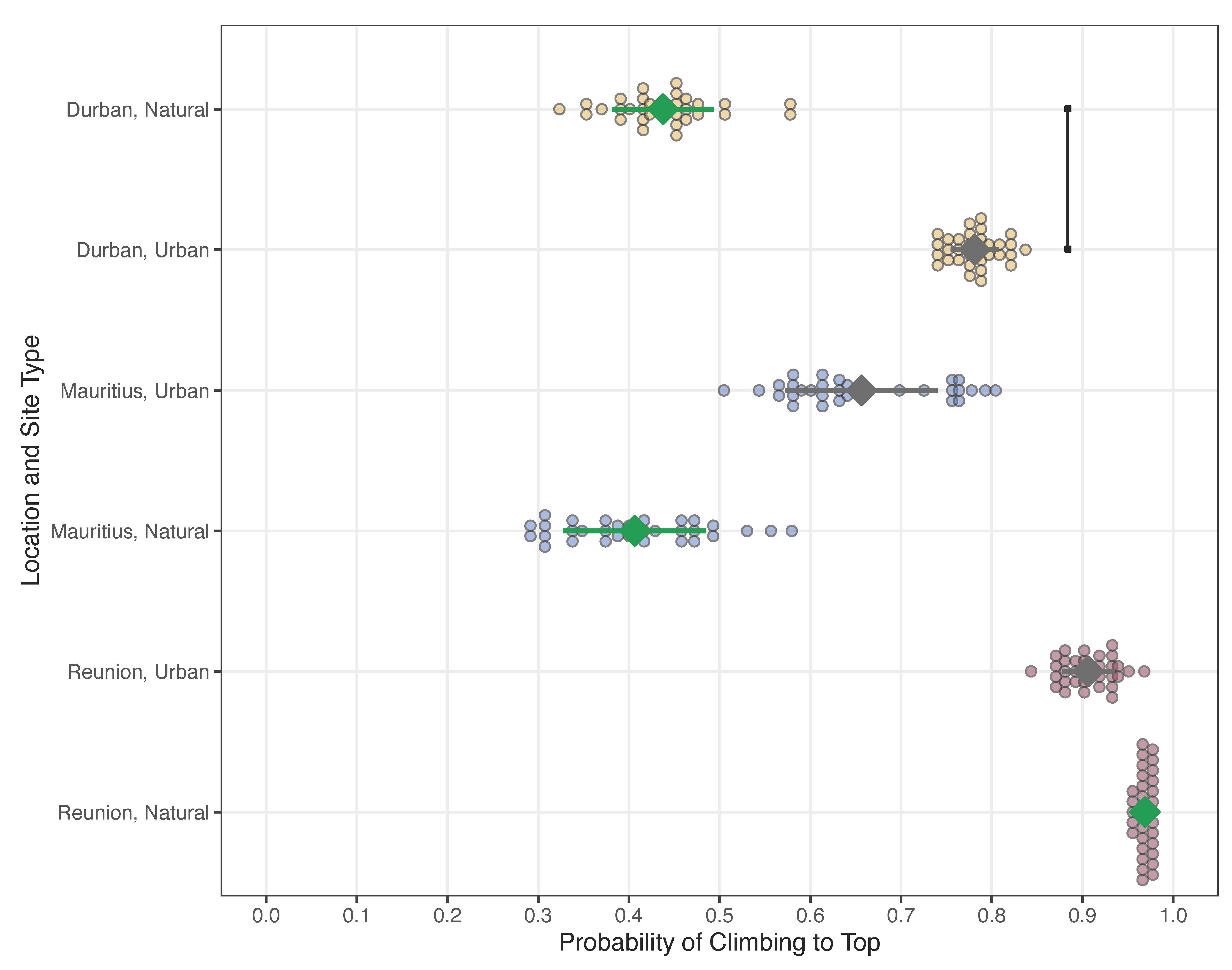
4.3. Climbing Ability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grant, P.R.; Grant, B.R.; Smith, J.N.; Abbott, I.J.; Abbott, L.K. Darwin’s finches: Population variation and natural selection. Proc. Natl. Acad. Sci. USA 1976, 73, 257–261. [Google Scholar] [CrossRef]
- Emerson, B.C. Evolution on oceanic islands: Molecular phylogenetic approaches to understanding pattern and process. Mol. Ecol. 2002, 11, 951–966. [Google Scholar] [CrossRef]
- Graham, N.R.; Gruner, D.S.; Lim, J.Y.; Gillespie, R.G. Island ecology and evolution: Challenges in the Anthropocene. Environ. Conserv. 2017, 44, 323–335. [Google Scholar] [CrossRef]
- Warren, B.H.; Simberloff, D.; Ricklefs, R.E.; Aguilée, R.; Condamine, F.L.; Gravel, D.; Morlon, H.; Mouquet, N.; Rosindell, J.; Casquet, J.; et al. Islands as model systems in ecology and evolution: Prospects fifty years after MacArthur-Wilson. Ecol. Lett. 2015, 18, 200–217. [Google Scholar] [CrossRef] [PubMed]
- Keller, S.R.; Taylor, D.R. History, chance and adaptation during biological invasion: Separating stochastic phenotypic evolution from response to selection. Ecol. Lett. 2008, 11, 852–866. [Google Scholar] [CrossRef]
- Littleford-Colquhoun, B.L.; Clemente, C.; Whiting, M.J.; Ortiz-Barrientos, D.; Frère, C.H. Archipelagos of the Anthropocene: Rapid and extensive differentiation of native terrestrial vertebrates in a single metropolis. Mol. Ecol. 2017, 26, 2466–2481. [Google Scholar] [CrossRef]
- Johnson, M.T.J.; Munshi-South, J. Evolution of life in urban environments. Science 2017, 358, eaam8327. [Google Scholar] [CrossRef]
- Start, D.; Barbour, M.A.; Bonner, C. Urbanization reshapes a food web. J. Anim. Ecol. 2019, 89, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Millien, V. Morphological Evolution Is accelerated among island mammals. PLoS Biol. 2006, 4, e321. [Google Scholar] [CrossRef]
- Holtmann, B.; Santos, E.S.A.; Lara, C.E.; Nakagawa, S. Personality-matching habitat choice, rather than behavioural plasticity, is a likely driver of a phenotype–environment covariance. Proc. R. Soc. B Boil. Sci. 2017, 284, 20170943. [Google Scholar] [CrossRef] [PubMed]
- Pellitteri-Rosa, D.; Bellati, A.; Cocca, W.; Gazzola, A.; Martín, J.; Fasola, M. Urbanization affects refuge use and habituation to predators in a polymorphic lizard. Anim. Behav. 2017, 123, 359–367. [Google Scholar] [CrossRef]
- Baxter-Gilbert, J.; Riley, J.L.; Whiting, M.J. Bold New World: Urbanization promotes an innate behavioral trait in a lizard. Behav. Ecol. Sociobiol. 2019, 73, 105. [Google Scholar] [CrossRef]
- Brown, C.R.; Brown, M.B. Where has all the road kill gone? Curr. Biol. 2013, 23, R233–R234. [Google Scholar] [CrossRef] [PubMed]
- Winchell, K.M.; Reynolds, R.G.; Prado-Irwin, S.R.; Puente-Rolón, A.R.; Revell, L.J. Phenotypic shifts in urban areas in the tropical lizard Anolis cristatellus. Evolution 2016, 70, 1009–1022. [Google Scholar] [CrossRef] [PubMed]
- Baxter-Gilbert, J.; Riley, J.L.; Frère, C.H.; Whiting, M.J. Shrinking into the big city: Influence of genetic and environmental factors on urban dragon lizard morphology and performance capacity. Urban Ecosyst. 2020, 24, 661–674. [Google Scholar] [CrossRef]
- Winchell, K.M.; Maayan, I.; Fredette, J.R.; Revell, L.J. Linking locomotor performance to morphological shifts in urban lizards. Proc. R. Soc. B Boil. Sci. 2018, 285, 20180229. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.S.; Turelli, M. Stable Underdominance and the evolutionary invasion of empty niches. Am. Nat. 1986, 127, 835–850. [Google Scholar] [CrossRef]
- Sol, D.; Griffin, A.; Bartomeus, I.; Boyce, H. Exploring or avoiding novel food resources? The novelty conflict in an invasive bird. PLoS ONE 2011, 6, e19535. [Google Scholar] [CrossRef] [PubMed]
- Palacio, F.X. Urban exploiters have broader dietary niches than urban avoiders. Ibis 2019, 162, 42–49. [Google Scholar] [CrossRef]
- de Rivera, C.E.; Ruiz, G.; Hines, A.H.; Jivoff, P. Biotic resistance to invasion: Native predator limits abundance and distribution of an introduced crab. Ecology 2005, 86, 3364–3376. [Google Scholar] [CrossRef]
- Loss, S.R.; Will, T.; Marra, P.P. Direct mortality of birds from anthropogenic causes. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 99–120. [Google Scholar] [CrossRef]
- Lapwong, Y.; Dejtaradol, A.; Webb, J. Shifts in thermal preference of introduced Asian house geckos (Hemidactylus frenatus) in temperate regions of southeastern Australia. J. Therm. Biol. 2020, 91, 102625. [Google Scholar] [CrossRef] [PubMed]
- Madelaire, C.B.; Barsotti, A.M.G.; Wagener, C.; Sugano, Y.Y.V.; Baxter-Gilbert, J.; Gomes, F.R.; Measey, J. Challenges of dehydration result in a behavioral shift in invasive toads. Behav. Ecol. Sociobiol. 2020, 74, 83. [Google Scholar] [CrossRef]
- Hufbauer, R.A.; Facon, B.; Ravigné, V.; Turgeon, J.; Foucaud, J.; Lee, C.E.; Rey, O.; Estoup, A. Anthropogenically induced adaptation to invade (AIAI): Contemporary adaptation to human-altered habitats within the native range can promote invasions. Evol. Appl. 2011, 5, 89–101. [Google Scholar] [CrossRef]
- Borden, J.B.; Flory, S.L. Urban evolution of invasive species. Front. Ecol. Environ. 2021, 19, 184–191. [Google Scholar] [CrossRef]
- Baxter-Gilbert, J.; Riley, J.L.; Measey, J. Fortune favors the bold toad: Urban-derived behavioral traits may provide advantages for invasive amphibian populations. Behav. Ecol. Sociobiol. 2021, 75, 130. [Google Scholar] [CrossRef]
- Estoup, A.; Guillemaud, T. Reconstructing routes of invasion using genetic data: Why, how and so what? Mol. Ecol. 2010, 19, 4113–4130. [Google Scholar] [CrossRef]
- Williamson, M. Biological Invasions; Chapman & Hall: London, UK, 1996; p. 244. [Google Scholar]
- Lombaert, E.; Guillemaud, T.; Cornuet, J.-M.; Malausa, T.; Facon, B.; Estoup, A. Bridgehead effect in the worldwide invasion of the biocontrol harlequin ladybird. PLoS ONE 2010, 5, e9743. [Google Scholar] [CrossRef]
- Bertelsmeier, C.; Ollier, S.; Liebhold, A.M.; Brockerhoff, E.G.; Ward, D.; Keller, L. Recurrent bridgehead effects accelerate global alien ant spread. Proc. Natl. Acad. Sci. USA 2018, 115, 5486–5491. [Google Scholar] [CrossRef]
- Pemberton, C.E. Introduction to Hawaii of the tropical American toad Bufo marinus. Hawaii. Plant. Rec. 1933, 38, 15–16. [Google Scholar]
- Oliver, J.A. The peripatetic toad. Nat. Hist. 1949, 58, 29–33. [Google Scholar]
- Zug, G.R.; Lindgren, E.; Pippet, J.R. Distribution and ecology of the marine toad, Bufo marinus, in Papua New Guinea. Pac. Sci. 1975, 29, 31–50. [Google Scholar]
- Shine, R. The Ecological impact of invasive cane toads (Bufo Marinus) in Australia. Q. Rev. Biol. 2010, 85, 253–291. [Google Scholar] [CrossRef] [PubMed]
- Gruber, J.; Brown, G.; Whiting, M.J.; Shine, R. Behavioural divergence during biological invasions: A study of cane toads (Rhinella marina) from contrasting environments in Hawai’i. R. Soc. Open Sci. 2018, 5, 180197. [Google Scholar] [CrossRef] [PubMed]
- Seebacher, F.; Franklin, C. Physiology of invasion: Cane toads are constrained by thermal effects on physiological mechanisms that support locomotor performance. J. Exp. Biol. 2011, 214, 1437–1444. [Google Scholar] [CrossRef]
- Hudson, C.M.; McCurry, M.R.; Lundgren, P.; McHenry, C.R.; Shine, R. Constructing an invasion machine: The rapid evolution of a dispersal-enhancing phenotype during the cane toad invasion of Australia. PLoS ONE 2016, 11, e0156950. [Google Scholar] [CrossRef]
- Gruber, J.; Brown, G.; Whiting, M.J.; Shine, R. Is the behavioural divergence between range-core and range-edge populations of cane toads (Rhinella marina) due to evolutionary change or developmental plasticity? R. Soc. Open Sci. 2017, 4, 170789. [Google Scholar] [CrossRef]
- Selechnik, D.; Richardson, M.F.; Shine, R.; DeVore, J.L.; Ducatez, S.; Rollins, L.A. Increased Adaptive Variation Despite Reduced Overall Genetic Diversity in a Rapidly Adapting Invader. Front. Genet. 2019, 10, 1221. [Google Scholar] [CrossRef]
- Battles, A.C.; Irschick, D.J.; Kolbe, J.J. Do structural habitat modifications associated with urbanization influence locomotor performance and limb kinematics in Anolis lizards? Biol. J. Linn. Soc. 2019, 127, 100–112. [Google Scholar] [CrossRef]
- Waselkov, K.E.; Regenold, N.D.; Lum, R.C.; Olsen, K.M. Agricultural adaptation in the native North American weed waterhemp, Amaranthus tuberculatus (Amaranthaceae). PLoS ONE 2020, 15, e0238861. [Google Scholar] [CrossRef]
- Rey, O.; Estoup, A.; Vonshak, M.; Loiseau, A.; Blanchet, S.; Calcaterra, L.; Chifflet, L.; Rossi, J.-P.; Kergoat, G.J.; Foucaud, J.; et al. Where do adaptive shifts occur during invasion? A multidisciplinary approach to unravelling cold adaptation in a tropical ant species invading the Mediterranean area. Ecol. Lett. 2012, 15, 1266–1275. [Google Scholar] [CrossRef]
- Jackson, H.; Strubbe, D.; Tollington, S.; Prys-Jones, R.; Matthysen, E.; Groombridge, J.J. Ancestral origins and invasion pathways in a globally invasive bird correlate with climate and influences from bird trade. Mol. Ecol. 2015, 24, 4269–4285. [Google Scholar] [CrossRef]
- Strubbe, D.; Jackson, H.; Groombridge, J.; Matthysen, E. Invasion success of a global avian invader is explained by within-taxon niche structure and association with humans in the native range. Divers. Distrib. 2015, 21, 675–685. [Google Scholar] [CrossRef]
- Cheke, A.; Hume, J.P. Lost Land of the Dodo: The Ecological History of Mauritius, Réunion and Rodrigues; T & AD Poyser: London, UK, 2010. [Google Scholar]
- Telford, N.S.; Channing, A.; Measey, J. Origin of invasive populations of the Guttural Toad (Sclerophrys gutturalis) on Reunion and Mauritius Islands and in Constantia, South Africa. Herpeto-Log. Conserv. Biol. 2019, 14, 380–392. [Google Scholar]
- Vimercati, G.; Davies, S.J.; Measey, J. Rapid adaptive response to a mediterranean environment reduces phenotypic mismatch in a recent amphibian invader. J. Exp. Biol. 2018, 221, jeb174797. [Google Scholar] [CrossRef]
- Barsotti, A.M.G.; Madelaire, C.B.; Wagener, C.; Titon, B., Jr.; Measey, J.; Gomes, F.R. Challenges of a novel range: Water balance, stress, and immunity in an invasive toad. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 253, 110870. [Google Scholar] [CrossRef] [PubMed]
- Mühlenhaupt, M.; Baxter-Gilbert, J.; Makhubo, B.G.; Riley, J.L.; Measey, J. Growing up in a new world: Trait divergence between rural, urban, and invasive populations of an amphibian urban invader. NeoBiota 2021, 69, 103–132. [Google Scholar] [CrossRef]
- Baxter-Gilbert, J.; Riley, J.L.; Wagener, C.; Mohanty, N.P.; Measey, J. Shrinking before our isles: The rapid expression of insular dwarfism in two invasive populations of guttural toad (Sclerophrys gutturalis). Biol. Lett. 2020, 16, 20200651. [Google Scholar] [CrossRef]
- Hudson, C.M.; Brown, G.P.; Shine, R. Athletic anurans: The impact of morphology, ecology and evolution on climbing ability in invasive cane toads. Biol. J. Linn. Soc. 2016, 119, 992–999. [Google Scholar] [CrossRef]
- Llewelyn, J.; Phillips, B.; Alford, R.; Schwarzkopf, L.; Shine, R. Locomotor performance in an invasive species: Cane toads from the invasion front have greater endurance, but not speed, compared to conspecifics from a long-colonised area. Oecologia 2009, 162, 343–348. [Google Scholar] [CrossRef] [PubMed]
- du Preez, L.H.; Weldon, C.; Cunningham, M.J.; Turner, A.A. Bufo gutturalis Power, 1927. In Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland; Minter, L.R., Burger, M., Harrison, J.A., Braack, H.H., Bishop, P.J., Kloepfer, D., Eds.; SI/MAB Series #9; Smithsonian Institute: Washington, DC, USA, 2004; pp. 67–69. [Google Scholar]
- Baxter-Gilbert, J.; Florens, F.B.V.; Baider, C.; Perianen, Y.D.; Citta, D.S.; Appadoo, C.; Measey, J. Toad-kill: Prey diversity and preference of invasive guttural toads (Sclerophrys gutturalis) in Mauritius. Afr. J. Ecol. 2020, 59, 168–177. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Florens, F.B.V.; Baider, C. Ecological restoration in a developing island nation: How useful is the science? Restor. Ecol. 2012, 21, 1–5. [Google Scholar] [CrossRef]
- Florens, F.B.V.; Baider, C.; Martin, G.M.N.; Strasberg, D. Surviving 370 years of human impact: What remains of tree diversity and structure of the lowland wet forests of oceanic island Mauritius? Biodivers. Conserv. 2012, 21, 2139–2167. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2022. Available online: https://www.R-project.org/ (accessed on 28 April 2022).
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- du Preez, L.H.; Carruthers, V.C. Frogs of Southern Africa; Penguin Random House: Cape Town, South Africa, 2017. [Google Scholar]
- Lleonart, J.; Salat, J.; Torres, G.J. Removing allometric effects of body size in morphological analysis. J. Theor. Biol. 2000, 205, 85–93. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package (Version 1.4.7). 2020. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 17 June 2020).
- Nations, J.A.; Heaney, L.R.; Demos, T.C.; Achmadi, A.S.; Rowe, K.C.; A Esselstyn, J. A simple skeletal measurement effectively predicts climbing behaviour in a diverse clade of small mammals. Biol. J. Linn. Soc. 2019, 128, 323–336. [Google Scholar] [CrossRef]
- Aerts, P.; Van Damme, R.; Vanhooydonck, B.; Zaaf, A.; Herrel, A. Lizard locomotion: How mor-phology meets ecology. Neth. J. Zool. 2000, 50, 261–277. [Google Scholar] [CrossRef]
- Garland, T., Jr.; Losos, J.B. Ecological morphology of locomotor performance in squamate reptiles. In Ecological Morphology: Integrative Organismal Biology; Wainwright, P.C., Reilly, S.M., Eds.; University of Chicago Press: Chicago, IL, USA, 1994; pp. 240–302. [Google Scholar]
- Bauwens, D.; Garland, T.; Castilla, A.M.; Van Damme, R. Evolution of sprint speed in lacertid lizards: Morphological, physiological, and behavioral covariation. Evolution 1995, 49, 848–863. [Google Scholar] [CrossRef]
- Vanhooydonck, B.; James, B.; Tallis, J.; Aerts, P.; Tadic, Z.; Tolley, K.A.; Measey, G.J.; Herrel, A.; Leducq, J.-B.; Charron, G.; et al. Is the whole more than the sum of its parts? Evolutionary trade-offs between burst and sustained locomotion in lacertid lizards. Proc. R. Soc. B Boil. Sci. 2014, 281, 20132677. [Google Scholar] [CrossRef] [PubMed]
- Keane, R.; Crawley, M.J. Exotic plant invasions and the enemy release hypothesis. Trends Ecol. Evol. 2002, 17, 164–170. [Google Scholar] [CrossRef]
- Colautti, R.I.; Ricciardi, A.; Grigorovich, I.A.; MacIsaac, H.J. Is invasion success explained by the enemy release hypothesis? Ecol. Lett. 2004, 7, 721–733. [Google Scholar] [CrossRef]
- Huey, R.; Bennett, A.F.; John-Alder, H.; Nagy, K.A. Locomotor capacity and foraging behaviour of kalahari lacertid lizards. Anim. Behav. 1984, 32, 41–50. [Google Scholar] [CrossRef]
- Vanhooydonck, B.; Van Damme, R.; Aerts, P. Speed and stamina trade-ff in lacertid lizards. Evolution 2001, 55, 1040–1048. [Google Scholar] [CrossRef]
- Herrel, A.; Bonneaud, C. Trade-offs between burst performance and maximal exertion capacity in a wild amphibian, Xenopus tropicalis. J. Exp. Biol. 2012, 215, 3106–3111. [Google Scholar] [CrossRef] [PubMed]
- Sorci, G.; Swallow, J.G.; Garland, T.; Clobert, J. Quantitative genetics of locomotor epeed and Endurance in the lizard Lacerta vivipara. Physiol. Zooll. 1995, 68, 698–720. [Google Scholar] [CrossRef]
- Garland, T. Physiological correlates of locomotory performance in a lizard: An allometric approach. Am. J. Physiol. Integr. Comp. Physiol. 1984, 247, R806–R815. [Google Scholar] [CrossRef]
- Wang, A.Z.; Husak, J.F.; Lovern, M. Leptin ameliorates the immunity, but not reproduction, trade-off with endurance in lizards. J. Comp. Physiol. B 2019, 189, 261–269. [Google Scholar] [CrossRef]
- Phillips, B.L.; Brown, G.P.; Webb, J.K.; Shine, R. Invasion and the evolution of speed in toads. Nature 2006, 439, 803. [Google Scholar] [CrossRef]
- McNab, B.K. Bioenergetics and the determination of home range size. Am. Nat. 1963, 97, 133–140. [Google Scholar] [CrossRef]
- Schoener, T.W.; Schoener, A. Intraspecific variation in home-range size in some anolis lizards. Ecology 1982, 63, 809–823. [Google Scholar] [CrossRef]
- Makarieva, A.M.; Gorshkov, V.G.; Li, B.-L. Why do population density and inverse home range scale differently with body size? Implications for ecosystem stability. Ecol. Complex. 2005, 2, 259–271. [Google Scholar] [CrossRef]
- Husak, J.F.; Keith, A.R.; Wittry, B.N. Making Olympic lizards: The effects of specialised exercise training on performance. J. Exp. Biol. 2015, 218, 899–906. [Google Scholar] [CrossRef]
- Hitchings, S.P.; Beebee, T.J.C. Genetic substructuring as a result of barriers to gene flow in urban Rana temporaria (common frog) populations: Implications for biodiversity conservation. Heredity 1997, 79, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Delaney, K.S.; Riley, S.P.D.; Fisher, R.N. A rapid, Strong, and convergent genetic response to urban habitat fragmentation in four divergent and widespread vertebrates. PLoS ONE 2010, 5, e12767. [Google Scholar] [CrossRef]
- Bergey, E.A. Dispersal of a non-native land snail across a residential area is modified by yard management and movement barriers. Urban Ecosyst. 2018, 22, 325–334. [Google Scholar] [CrossRef]
- Bates, A.J.; Sadler, J.P.; Fairbrass, A.J.; Falk, S.J.; Hale, J.D.; Matthews, T.J. Changing Bee and Hoverfly Pollinator Assemblages along an Urban-Rural Gradient. PLoS ONE 2011, 6, e23459. [Google Scholar] [CrossRef]
- Braby, M.F.; Williams, M.R.; Douglas, F.; Beardsell, C.; Crosby, D.F. Changes in a peri-urban butterfly assemblage over 80 years near Melbourne, Australia. Austral Entomol. 2021, 60, 27–51. [Google Scholar] [CrossRef]
- Florens, F.B.V.; Baider, C. Relocation of Omphalotropis plicosa (Pfeiffer, 1852), a Mauritian endemic landsnail believed extinct. J. Molluscan Stud. 2007, 73, 205–206. [Google Scholar] [CrossRef]
- Griffiths, O.L.; Florens, F.B.V. A Field Guide to the Non-Marine Molluscs of the Mascarene Islands (Mauritius, Rodrigues, Réunion) and the Northern Dependencies of Mauritius; Bioculture Press: Riviere des Anguilles, Mauritius, 2006. [Google Scholar]
- Moya-Laraño, J.; Halaj, J.; Wise, D.H. Climbing to reach females: Romeo should be small. Evolution 2002, 56, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Edgington, H.A.; Taylor, D.R. Ecological contributions to body shape evolution in salamanders of the genus Eurycea (Plethodontidae). PLoS ONE 2019, 14, e0216754. [Google Scholar] [CrossRef] [PubMed]
- Maida, J.R.; Bishop, C.A.; Larsen, K.W. Migration and disturbance: Impact of fencing and development on Western Rattlesnake (Crotalus oreganus) spring movements in British Columbia. Can. J. Zooll. 2020, 98, 1–12. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Diedenhofen, B.; Musch, J. Cocor: A comprehensive solution for the statistical comparison of correlations. PloS ONE 2015, 10, e0121945. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Ellison, A.M. A Primer of Ecological Statistics; Sinauer Associates Inc.: Sunderland, MA, USA, 2004; Volume 1. [Google Scholar]
- Mayer, M. Confintr: Confidence Intervals. R Package Version 0.1.2. 2022. Available online: https://CRAN.R-project.org/package=confintr (accessed on 28 April 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Coordinates | Elevation | Location | Site Type | nF | nM | ntotal |
|---|---|---|---|---|---|---|---|
| Total for Durban (Native Range) and Natural | 33 | 25 | 58 | ||||
| Amatikulu Nature Reserve | 29.11° S, 31.60° E | 5 | Durban | Natural | 2 | 1 | 3 |
| Orchard Near Ballito | 29.47° S, 31.23° E | 50 | Durban | Natural | 6 | 5 | 11 |
| Private Reserve near Hilton | 29.50° S, 30.28° E | 1030 | Durban | Natural | 25 | 19 | 44 |
| Total for Durban (Native Range) and Urban | 59 | 34 | 93 | ||||
| Durban Botanic Gardens | 29.85° S, 31.01° E | 40 | Durban | Urban | 59 | 34 | 93 |
| Total for Mauritius (Invasive Range) and Natural | 51 | 20 | 71 | ||||
| Black River Gorges National Park | 20.37° S, 57.44° E | 580 | Mauritius | Natural | 51 | 20 | 71 |
| Total for Mauritius (Invasive Range) and Urban | 43 | 44 | 87 | ||||
| Notre Dame | 20.14° S, 57.56° E | 130 | Mauritius | Urban | 43 | 44 | 87 |
| Total for Réunion (Invasive Range) and Natural | 49 | 31 | 80 | ||||
| Point Payet | 21.10° S, 55.66° E | 510 | Réunion | Natural | 49 | 31 | 80 |
| Total for Réunion (Invasive Range) and Urban | 73 | 33 | 106 | ||||
| Villèle | 21.06° S, 55.26° E | 380 | Réunion | Urban | 73 * | 33 | 106 |
| Total Across Locations and Site Types | 308 | 187 | 495 | ||||
| Location | Site Type | Population | Escape Speed (s) | Endurance Distance (m) | Climbing Ability | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| nF | nM | ntotal | nF | nM | ntotal | nF | nM | ntotal | |||
| Durban | Natural | Private Reserve near Hilton | 16 | 16 | 32 | 16 | 16 | 32 | 16 | 16 | 32 |
| Durban | Urban | Durban Botanic Gardens | 16 | 16 | 32 | 16 | 16 | 32 | 16 | 16 | 32 |
| Mauritius | Natural | Black River Gorges National Park | 16 | 16 | 32 | 16 | 16 | 32 | 16 | 16 | 32 |
| Mauritius | Urban | Notre Dame | 16 | 15 | 31 | 16 | 16 | 32 | 16 | 16 | 32 |
| Réunion | Natural | Point Payet | 16 | 17 | 33 | 16 | 17 | 33 | 16 | 17 | 33 |
| Réunion | Urban | Villèle | 16 | 16 | 32 | 16 | 16 | 32 | 16 | 16 | 32 |
| Total Across Locations and Site Types | 96 | 96 | 192 | 96 | 97 | 193 | 96 | 97 | 193 | ||
| (a) Output for the LMM Analyzing Snout-Vent Length. | ||||
|---|---|---|---|---|
| Variable Names | ||||
| Fixed Effects | β | SE | t | p |
| Intercept (Durban, Natural, Female) | 1.844 | 0.015 | 119.052 | <0.001 |
| Study Location (Mauritius) | −0.153 | 0.022 | −6.849 | 0.004 |
| Study Location (Réunion) | −0.099 | 0.022 | −4.430 | 0.017 |
| Site Type (Urban) | 0.007 | 0.019 | 0.377 | 0.730 |
| Sex (Male) | −0.006 | 0.005 | −1.129 | 0.259 |
| Random Effects | σ2 | |||
| Population | 0.001 | |||
| Residuals | 0.003 | |||
| (b) Multiple Comparisons between Study Locations | ||||
| Study Locations | β | SE | t | pcorr |
| Durban vs. Mauritius | 0.153 | 0.023 | 6.786 | 0.013 |
| Durban vs. Réunion | 0.099 | 0.023 | 4.389 | 0.044 |
| Mauritius vs. Réunion | −0.054 | 0.024 | −2.244 | 0.209 |
| (a) Output for the LMM Analyzing Hindlimb Length | ||||
|---|---|---|---|---|
| Variable Names | ||||
| Fixed Effects | β | SE | T | P |
| Intercept (Durban, Natural, Female) | 0.115 | 0.037 | 3.095 | 0.002 |
| Snout-Vent Length | 0.886 | 0.020 | 44.407 | <0.001 |
| Study Location (Mauritius) | −0.035 | 0.005 | −7.746 | <0.001 |
| Study Location (Réunion) | −0.018 | 0.004 | −4.858 | 0.002 |
| Site Type (Urban) | −0.005 | 0.002 | −2.141 | 0.129 |
| Sex (Male) | 0.003 | 0.002 | 1.409 | 0.159 |
| Random Effects | σ2 | |||
| Population | 0.000 | |||
| Residuals | 0.001 | |||
| (b) Multiple Comparisons between Study Locations | ||||
| Study Locations | β | SE | T | pcorr |
| Durban vs. Mauritius | 0.035 | 0.005 | 7.654 | <0.001 |
| Durban vs. Réunion | 0.018 | 0.004 | 4.698 | 0.020 |
| Mauritius vs. Réunion | −0.017 | 0.003 | −5.294 | 0.060 |
| (a) Output for the LMM Analyzing Hindfoot Length | ||||
|---|---|---|---|---|
| Variable Names | ||||
| Fixed Effects | β | SE | T | p |
| Intercept (Durban, Natural, Female) | 0.230 | 0.044 | 5.209 | <0.001 |
| Snout-Vent Length | 0.771 | 0.024 | 32.515 | <0.001 |
| Study Location (Mauritius) | −0.072 | 0.008 | −8.914 | <0.001 |
| Study Location (Réunion) | −0.035 | 0.007 | −4.738 | 0.007 |
| Site Type (Urban) | −0.017 | 0.006 | −2.974 | 0.052 |
| Sex (Male) | 0.003 | 0.003 | 1.319 | 0.187 |
| Random Effects | σ2 | |||
| Population | 0.000 | |||
| Residuals | 0.001 | |||
| (b) Multiple Comparisons between Study Locations | ||||
| Study Locations | β | SE | T | pcorr |
| Durban vs. Mauritius | 0.072 | 0.008 | 8.792 | 0.002 |
| Durban vs. Réunion | 0.035 | 0.008 | 4.658 | 0.037 |
| Mauritius vs. Réunion | −0.036 | 0.008 | −4.804 | 0.073 |
| (a) Output for the LMM Analyzing Escape Speed | ||||
|---|---|---|---|---|
| Variable Names | ||||
| Fixed Effects | β | SE | T | p |
| Intercept (Durban, Natural, Female) | −0.106 | 0.097 | −1.103 | 0.272 |
| Snout-Vent Length | 0.171 | 0.048 | 3.569 | <0.001 |
| Study Location (Mauritius) | −0.051 | 0.010 | −4.903 | <0.001 |
| Study Location (Réunion) | −0.031 | 0.008 | −4.069 | <0.001 |
| Site Type (Urban) | −0.005 | 0.004 | −1.214 | 0.225 |
| Sex (Male) | 0.000 | 0.005 | −0.092 | 0.927 |
| Order | 0.013 | 0.003 | 4.640 | <0.001 |
| Random Effects | σ2 | |||
| Researcher Identity | 0.004 | |||
| Experimental Group | 0.000 | |||
| Residuals | 0.003 | |||
| (b) Multiple Comparisons between Study Locations | ||||
| Study Locations | β | SE | T | pcorr |
| Durban vs. Mauritius | 0.051 | 0.010 | 4.885 | <0.001 |
| Durban vs. Réunion | 0.031 | 0.008 | 4.053 | <0.001 |
| Mauritius vs. Réunion | −0.020 | 0.007 | −2.954 | 0.009 |
| (a) Output for the LMM Analyzing Endurance Distance | ||||
|---|---|---|---|---|
| Variable Names | ||||
| Fixed Effects | Β | SE | t | P |
| Intercept (Durban, Natural, Female) | 153.494 | 175.397 | 0.875 | 0.386 |
| Snout-Vent Length | 48.429 | 83.991 | 0.577 | 0.564 |
| Study Location (Mauritius) | −38.993 | 17.828 | −2.187 | 0.029 |
| Study Location (Réunion) | −123.479 | 14.942 | −8.264 | <0.001 |
| Site Type (Urban) | 9.634 | 13.052 | 0.732 | 0.461 |
| Sex (Male) | 59.321 | 7.718 | 7.687 | <0.001 |
| Order | 0.311 | 4.481 | 0.069 | 0.945 |
| Study Location (Mauritius) × Site Type (Urban) | −15.389 | 18.927 | −0.813 | 0.417 |
| Study Location (Réunion) × Site Type (Urban) | 8.634 | 18.054 | 0.478 | 0.633 |
| Random Effects | σ2 | |||
| Researcher Identity | 22848.780 | |||
| Experimental Group | 33.470 | |||
| Residuals | 7499.740 | |||
| (b) Multiple Comparisons between Interaction Effects | ||||
| Study Locations and Site Types | Β | SE | T | pcorr |
| DN vs. MN | 38.990 | 17.900 | 2.184 | 0.229 |
| DN vs. RN | 123.480 | 15.000 | 8.256 | <0.001 |
| (1) DN vs. DU | −9.630 | 13.100 | −0.738 | 0.974 |
| DN vs. MU | 44.750 | 19.500 | 2.291 | 0.184 |
| DN vs. RU | 105.210 | 14.800 | 7.113 | <0.001 |
| (2) DU vs. MU | 54.380 | 22.300 | 2.434 | 0.134 |
| DU vs. RU | 114.840 | 16.600 | 6.928 | <0.001 |
| MN vs. RN | 84.490 | 13.600 | 6.199 | <0.001 |
| MN vs. DU | −48.630 | 20.400 | −2.379 | 0.152 |
| (3) MN vs. MU | 5.760 | 13.000 | 0.442 | 0.998 |
| MN vs. RU | 66.220 | 14.000 | 4.730 | <0.001 |
| (4) MU vs. RU | 60.460 | 15.400 | 3.938 | 0.001 |
| RN vs. DU | −133.110 | 16.900 | −7.897 | <0.001 |
| RN vs. MU | −78.730 | 14.900 | −5.277 | <0.001 |
| (5) RN vs. RU | −18.270 | 12.700 | −1.444 | 0.678 |
| (c) Summary of Raw Data | ||||
| Study Locations and Site Types | Average ± SE | |||
| DN | 185.688 ± 10.978 | |||
| DU | 200.047 ± 13.441 | |||
| MN | 137.418 ± 9.042 | |||
| MU | 129.659 ± 11.020 | |||
| RN | 56.911 ± 3.162 | |||
| RU | 75.874 ± 6.415 | |||
| (a) Output for the GLMM Analyzing Climbing Ability | ||||
|---|---|---|---|---|
| Variable Names | ||||
| Fixed Effects | β | SE | Z | p |
| Intercept (Durban, Natural, Female) | 10.877 | 8.058 | 1.350 | 0.177 |
| Snout-Vent Length | −5.884 | 4.315 | −1.363 | 0.173 |
| Study Location (Mauritius) | −0.937 | 0.783 | −1.197 | 0.231 |
| Study Location (Réunion) | 3.281 | 1.116 | 2.938 | 0.003 |
| Site Type (Urban) | 1.789 | 0.594 | 3.013 | 0.003 |
| Sex (Male) | −0.479 | 0.379 | −1.262 | 0.207 |
| Study Location (Mauritius) × Site Type (Urban) | −0.915 | 0.825 | −1.108 | 0.268 |
| Study Location (Réunion) × Site Type (Urban) | −2.895 | 1.319 | −2.195 | 0.028 |
| Random Effects | σ2 | |||
| Experimental Group | 0.000 | |||
| Within-Day Batch | 0.000 | |||
| Residuals | 1.000 | |||
| (b) Multiple Comparisons between Interaction Effects | ||||
| Study Locations and Site Types | β | SE | Z | pcorr |
| DN vs. MN | 2.553 | 1.999 | 1.197 | 0.819 |
| DN vs. RN | 0.038 | 0.042 | −2.938 | 0.034 |
| (1) DR vs. DU | 0.167 | 0.099 | −3.013 | 0.027 |
| DN vs. MU | 1.065 | 0.927 | 0.073 | 1.000 |
| DN vs. RU | 0.114 | 0.085 | −2.918 | 0.036 |
| (2) DU vs. MU | 6.373 | 6.778 | 1.741 | 0.472 |
| DU vs. RU | 0.680 | 0.587 | −0.447 | 0.997 |
| MN vs. RN | 0.015 | 0.017 | −3.756 | 0.002 |
| MN vs. DU | 0.066 | 0.064 | −2.802 | 0.050 |
| (3) MN vs. MU | 0.417 | 0.224 | −1.631 | 0.546 |
| MN vs. RU | 0.045 | 0.036 | −3.906 | 0.001 |
| RN vs. DU | 4.445 | 5.367 | 1.236 | 0.798 |
| (4) MU vs. RU | 0.107 | 0.091 | −2.623 | 0.082 |
| RN vs. MU | 28.328 | 32.833 | 2.885 | 0.040 |
| (5) RN vs. RU | 3.022 | 3.587 | 0.932 | 0.929 |
| (c) Summary of Raw Data | ||||
| Study Locations and Site Types | Average ± SE | |||
| DN | 0.438 ± 0.089 | |||
| DU | 0.781 ± 0.074 | |||
| MN | 0.406 ± 0.088 | |||
| MU | 0.656 ± 0.085 | |||
| RN | 0.969 ± 0.031 | |||
| RU | 0.906 ± 0.052 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baxter-Gilbert, J.; Riley, J.L.; Wagener, C.; Baider, C.; Florens, F.B.V.; Kowalski, P.; Campbell, M.; Measey, J. Island Hopping through Urban Filters: Anthropogenic Habitats and Colonized Landscapes Alter Morphological and Performance Traits of an Invasive Amphibian. Animals 2022, 12, 2549. https://doi.org/10.3390/ani12192549
Baxter-Gilbert J, Riley JL, Wagener C, Baider C, Florens FBV, Kowalski P, Campbell M, Measey J. Island Hopping through Urban Filters: Anthropogenic Habitats and Colonized Landscapes Alter Morphological and Performance Traits of an Invasive Amphibian. Animals. 2022; 12(19):2549. https://doi.org/10.3390/ani12192549
Chicago/Turabian StyleBaxter-Gilbert, James, Julia L. Riley, Carla Wagener, Cláudia Baider, F. B. Vincent Florens, Peter Kowalski, May Campbell, and John Measey. 2022. "Island Hopping through Urban Filters: Anthropogenic Habitats and Colonized Landscapes Alter Morphological and Performance Traits of an Invasive Amphibian" Animals 12, no. 19: 2549. https://doi.org/10.3390/ani12192549
APA StyleBaxter-Gilbert, J., Riley, J. L., Wagener, C., Baider, C., Florens, F. B. V., Kowalski, P., Campbell, M., & Measey, J. (2022). Island Hopping through Urban Filters: Anthropogenic Habitats and Colonized Landscapes Alter Morphological and Performance Traits of an Invasive Amphibian. Animals, 12(19), 2549. https://doi.org/10.3390/ani12192549