
A Survey of Genome-Wide Genetic Characterizations of Crossbred Dairy Cattle in Local Farms in Cambodia

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample and DNA Extraction
2.2. Analysis of Milk Fat Content Genetic Structure of Crossbred Cows
3. Results
3.1. Summary of Milk Production in the Farms
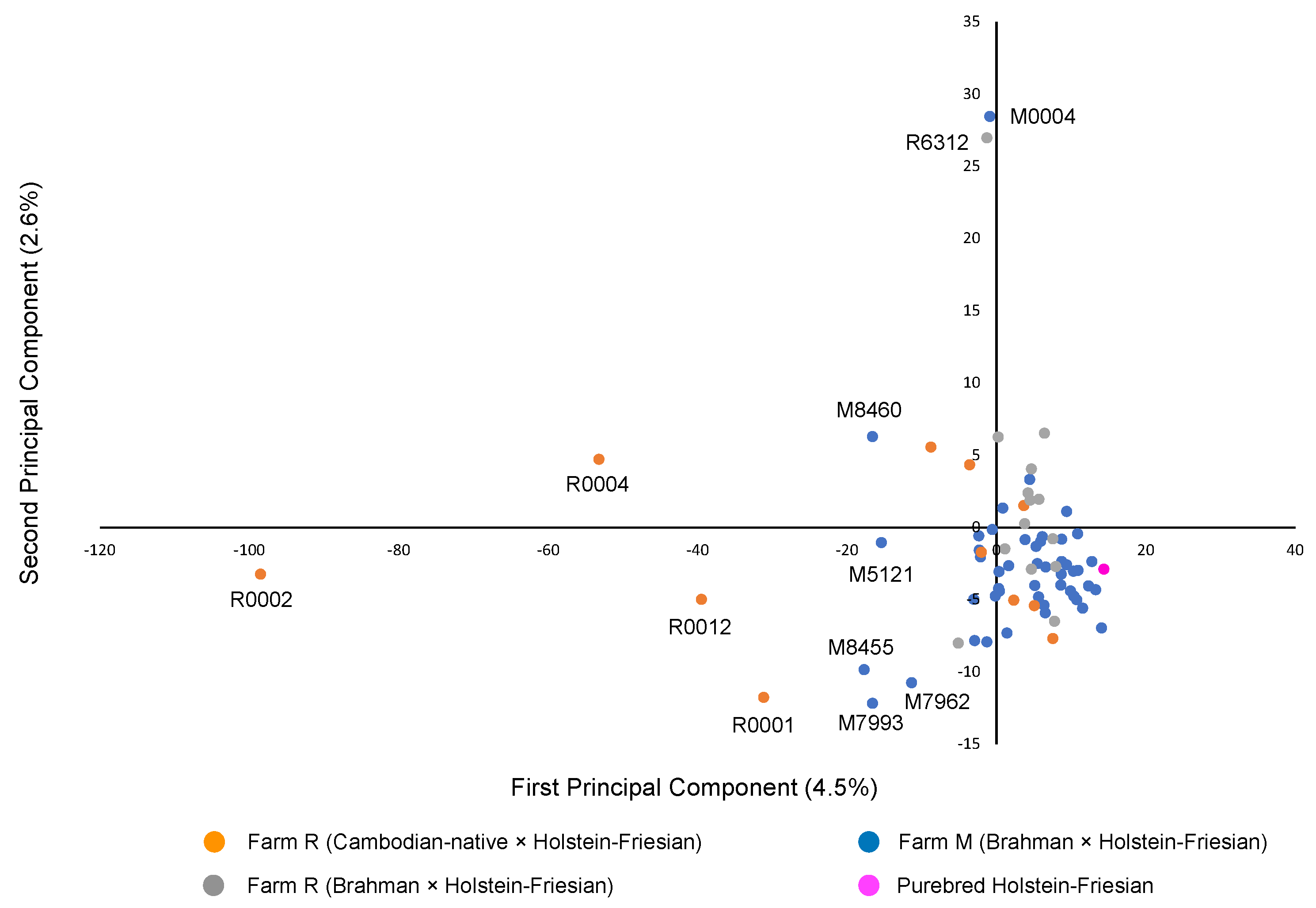
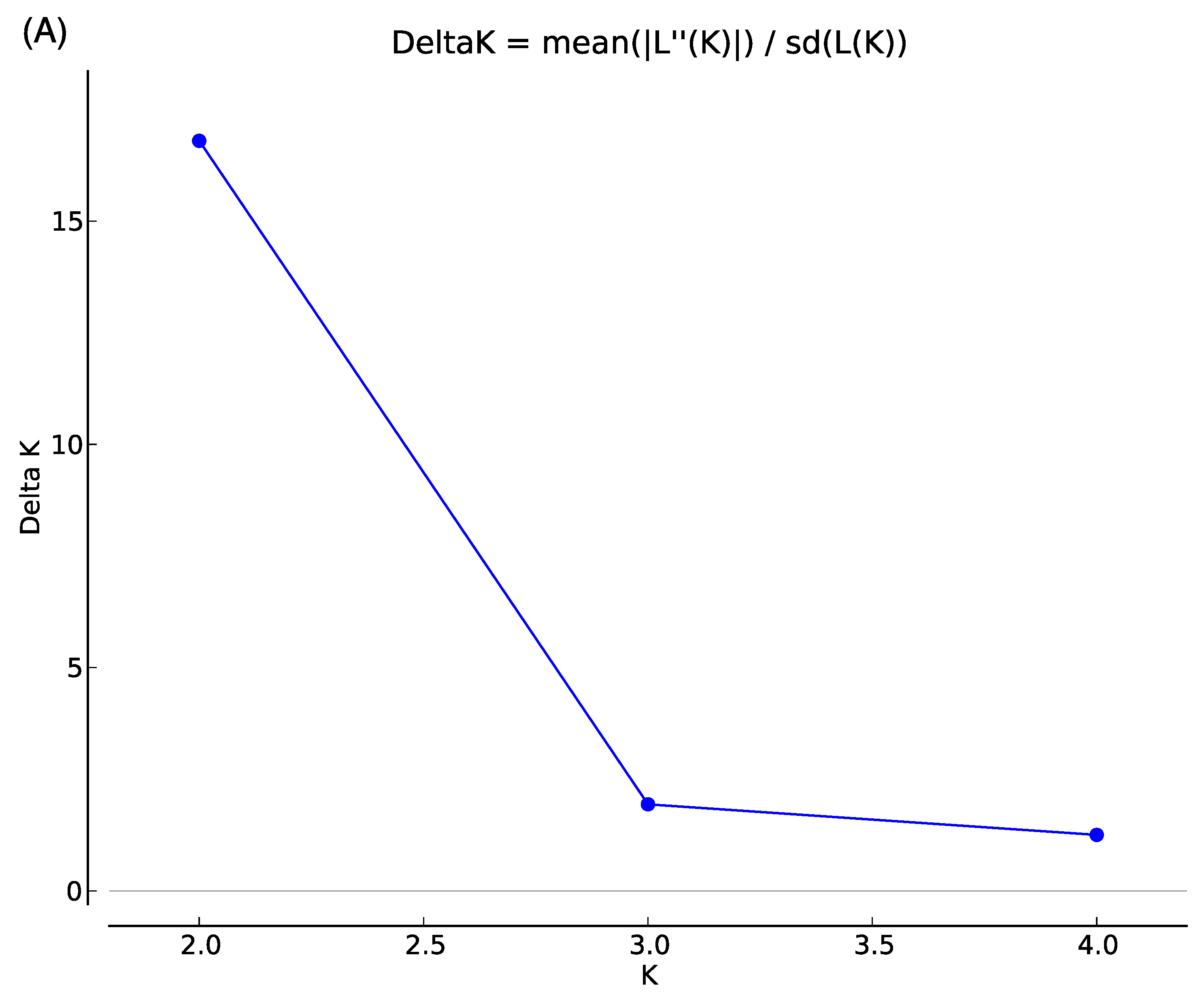
3.2. Genetic Structure
4. Discussion
4.1. Genetic Characteristics of Cambodian Dairy Cattle
4.2. Implications for Future Breeding in Cambodia
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tep, B.; Morita, Y.; Matsuyama, S.; Ohkura, S.; Inoue, N.; Tsukamura, H.; Uenoyama, Y.; Pheng, V. Seasonal changes in the reproductive performance in local cows receiving artificial insemination in the Pursat province of Cambodia. Asian-Australasian J. Anim. Sci. 2020, 33, 1922–1929. [Google Scholar] [CrossRef] [PubMed]
- Nampanya, S.; Suon, S.; Rast, L.; Windsor, P.A. Improvement in smallholder farmer knowledge of cattle production, health and biosecurity in Southern Cambodia between 2008 and 2010. Transbound. Emerg. Dis. 2012, 59, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Ieda, N.; Watanabe, Y.; Bun, C.; Vutha, P.; Uenoyama, Y. Characterization of Smallholder Livestock Farming in Kampong Cham Province, Cambodia: A Pilot Study in Prey Chhor District. J. Int. Coop. Agricult. Dev. 2017, 15, 42–47. Available online: https://icrea.agr.nagoya-u.ac.jp/jpn/journal/Vol15_42-47_05-WorkingPaper-Ieda.pdf (accessed on 1 December 2021).
- Decker, J.E.; McKay, S.D.; Rolf, M.M.; Kim, J.; Alcalá, A.M.; Sonstegard, T.S.; Hanotte, O.; Götherström, A.; Seabury, C.M.; Praharani, L.; et al. Worldwide Patterns of Ancestry, Divergence, and Admixture in Domesticated Cattle. PLoS Genet. 2014, 10, e1004254. [Google Scholar] [CrossRef] [PubMed]
- Sath, K.; Borin, K.; Preston, T.R. Survey on feed utilization for cattle production in Takeo province. Livestock Res. Rural Dev. 2008, 20. Available online: http://www.lrrd.org/lrrd20/supplement/sath1.htm (accessed on 1 August 2022).
- Baird, N.A.; Etter, P.D.; Atwood, T.S.; Currey, M.C.; Shiver, A.L.; Lewis, Z.A.; Selker, E.U.; Cresko, W.A.; Johnson, E.A. Rapid SNP discovery and genetic mapping using sequenced RAD markers. PLoS ONE 2008, 3, e3376. [Google Scholar] [CrossRef] [PubMed]
- Peterson, B.K.; Weber, J.N.; Kay, E.H.; Fisher, H.S.; Hoekstra, H.E. Double Digest RADseq: An Inexpensive Method for De Novo SNP Discovery and Genotyping in Model and Non-Model Species. PLoS ONE 2012, 7, e37135. [Google Scholar] [CrossRef] [PubMed]
- Suyama, Y.; Matsuki, Y. MIG-seq: An effective PCR-based method for genome-wide single-nucleotide polymorphism genotyping using the next-generation sequencing platform. Sci. Rep. 2015, 5, 16963. [Google Scholar] [CrossRef]
- Hosoya, S.; Hirase, S.; Kikuchi, K.; Nanjo, K.; Nakamura, Y.; Kohno, H.; Sano, M. Random PCR-based genotyping by sequencing technology GRAS-Di (genotyping by random amplicon sequencing, direct) reveals genetic structure of mangrove fishes. Mol. Ecol. Resour. 2019, 19, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- Miki, Y.; Yoshida, K.; Enoki, H.; Komura, S.; Suzuki, K.; Inamori, M.; Nishijima, R.; Takumi, S. GRAS-Di system facilitates high-density genetic map construction and QTL identification in recombinant inbred lines of the wheat progenitor Aegilops tauschii. Sci. Rep. 2020, 10, 21455. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, S.; Hamasaki, M.; Kadomura, K.; Yamada, T.; Chuda, H.; Kikuchi, K.; Hosoya, S. Genetic Dissection of a Precocious Phenotype in Male Tiger Pufferfish (Takifugu rubripes) using Genotyping by Random Amplicon Sequencing, Direct (GRAS-Di). Mar. Biotechnol. 2021, 23, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Nakajima, N.; Onuma, M.; Murayama, M. Genetic Diversity and Genetic Structure of the Wild Tsushima Leopard Cat from Genome-Wide Analysis. Animals 2020, 10, 1375. [Google Scholar] [CrossRef]
- Matsudaira, K.; Ishida, T. Divergence and introgression in small apes, the genus Hylobates, revealed by reduced representation sequencing. Heredity 2021, 127, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience 2015, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Porras-Hurtado, L.; Ruiz, Y.; Santos, C.; Phillips, C.; Carracedo, Á.; Lareu, M.V. An overview of STRUCTURE: Applications, parameter settings, and supporting software. Front. Genet. 2013, 4, 98. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed]
- Serey, M.; Mom, S.; Kouch, T.; Bunna, C. Cattle production systems in NW Cambodia. Livestock Res. Rural Dev. 2014, 26, 42. Available online: http://www.lrrd.org/lrrd26/3/sere26042.htm (accessed on 1 December 2021).
- Corbet, G.B.; Hill, J.E. The Mammals of the Indomalayan Region: A Systematic Review; Oxford University Press: Oxford, UK, 1992; Volume 488. [Google Scholar]
- Animal Husbandry Division, DLD. Beef Cattle Production and Management Systems in Livestock Research and Breeding Center in DLD; Department of Livestock Development, Minister of Agriculture and Cooperative: Bangkok, Thailand, 1991.
- Lin, B.Z.; Sasazaki, S.; Mannen, H. Genetic diversity and structure in Bos taurus and Bos indicus populations analyzed by SNP markers. Anim. Sci. J. 2010, 81, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Yonesaka, R.; Sasazaki, S.; Yasue, H.; Niwata, S.; Inayoshi, Y.; Mukai, F.; Mannen, H. Genetic structure and relationships of 16 Asian and European cattle populations using DigiTag2 assay. Anim. Sci. J. 2015, 87, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Rivera, E.J.; Rodríguez-Miranda, J.; Huerta-Mora, I.R.; Cárdenas-Cágal, A.; Juárez-Barrientos, J.M. Tropical milk production systems and milk quality: A review. Trop. Anim. Heal. Prod. 2019, 51, 1295–1305. [Google Scholar] [CrossRef]
- Bohmanova, J.; Misztal, I.; Cole, J. Temperature-Humidity Indices as Indicators of Milk Production Losses due to Heat Stress. J. Dairy Sci. 2007, 90, 1947–1956. [Google Scholar] [CrossRef]
- Kadzere, C.; Murphy, M.; Silanikove, N.; Maltz, E. Heat stress in lactating dairy cows: A review. Livest. Prod. Sci. 2002, 77, 59–91. [Google Scholar] [CrossRef]
- Li, Q.; Yang, C.; Du, J.; Zhang, B.; He, Y.; Hu, Q.; Li, M.; Zhang, Y.; Wang, C.; Zhong, J. Characterization of miRNA profiles in the mammary tissue of dairy cattle in response to heat stress. BMC Genom. 2018, 19, 975. [Google Scholar] [CrossRef]
- Ravagnolo, O.; Misztal, I. Genetic Component of Heat Stress in Dairy Cattle, Parameter Estimation. J. Dairy Sci. 2000, 83, 2126–2130. [Google Scholar] [CrossRef]
- Madalena, F.; Teodoro, R.; Lemos, A.; Monteiro, J.; Barbosa, R. Evaluation of Strategies for Crossbreeding of Dairy Cattle in Brazil. J. Dairy Sci. 1990, 73, 1887–1901. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Cattle Breed | Milk Production | ||||||
|---|---|---|---|---|---|---|---|
| Location (Province) | Herd Size | Cow Provider | Breeding | Breeding Method | Average Amount/Cow (Mean ± SD) | Average Milk Fat (Mean ± SD) | |
| Farm R | Phnom Penh city | 35 | Cambodian farmers | Holstein-Friesian (Thailand) × Cambodian Local | AI (Local, mating) | 7.81 ± 2.66 L/day | 3.77 ± 0.98% ** |
| Thailand breeders | Holstein-Friesian (Thailand) × Brahman | AI | |||||
| Farm M | Kandal | 300 | Thailand breeders | Holstein-Friesian (New Zealand) × Brahman | AI | 6.5–7.5 L/day † | 3.35 ± 0.54% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mam, S.; Tep, B.; Rin, S.; Uenoyama, Y.; Matsuyama, S.; Ohkura, S.; Murase, T.; Nunome, M.; Morita, Y. A Survey of Genome-Wide Genetic Characterizations of Crossbred Dairy Cattle in Local Farms in Cambodia. Animals 2022, 12, 2072. https://doi.org/10.3390/ani12162072
Mam S, Tep B, Rin S, Uenoyama Y, Matsuyama S, Ohkura S, Murase T, Nunome M, Morita Y. A Survey of Genome-Wide Genetic Characterizations of Crossbred Dairy Cattle in Local Farms in Cambodia. Animals. 2022; 12(16):2072. https://doi.org/10.3390/ani12162072
Chicago/Turabian StyleMam, Somony, Bengthay Tep, Soriya Rin, Yoshihisa Uenoyama, Shuichi Matsuyama, Satoshi Ohkura, Tetsuma Murase, Mitsuo Nunome, and Yasuhiro Morita. 2022. "A Survey of Genome-Wide Genetic Characterizations of Crossbred Dairy Cattle in Local Farms in Cambodia" Animals 12, no. 16: 2072. https://doi.org/10.3390/ani12162072
APA StyleMam, S., Tep, B., Rin, S., Uenoyama, Y., Matsuyama, S., Ohkura, S., Murase, T., Nunome, M., & Morita, Y. (2022). A Survey of Genome-Wide Genetic Characterizations of Crossbred Dairy Cattle in Local Farms in Cambodia. Animals, 12(16), 2072. https://doi.org/10.3390/ani12162072