
Regulation of a High-Iron Diet on Lipid Metabolism and Gut Microbiota in Mice


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Serum Biochemical Assays
2.3. Histological Analysis
2.4. RNA Extraction and Quantitative Real-Time PCR
2.5. Western Blot Analysis
2.6. Gut Microbiota Analysis
2.7. Statistical Analysis
3. Result
3.1. A High-Iron Diet Reduced the Body Weight and Changed Iron Metabolism of Mice
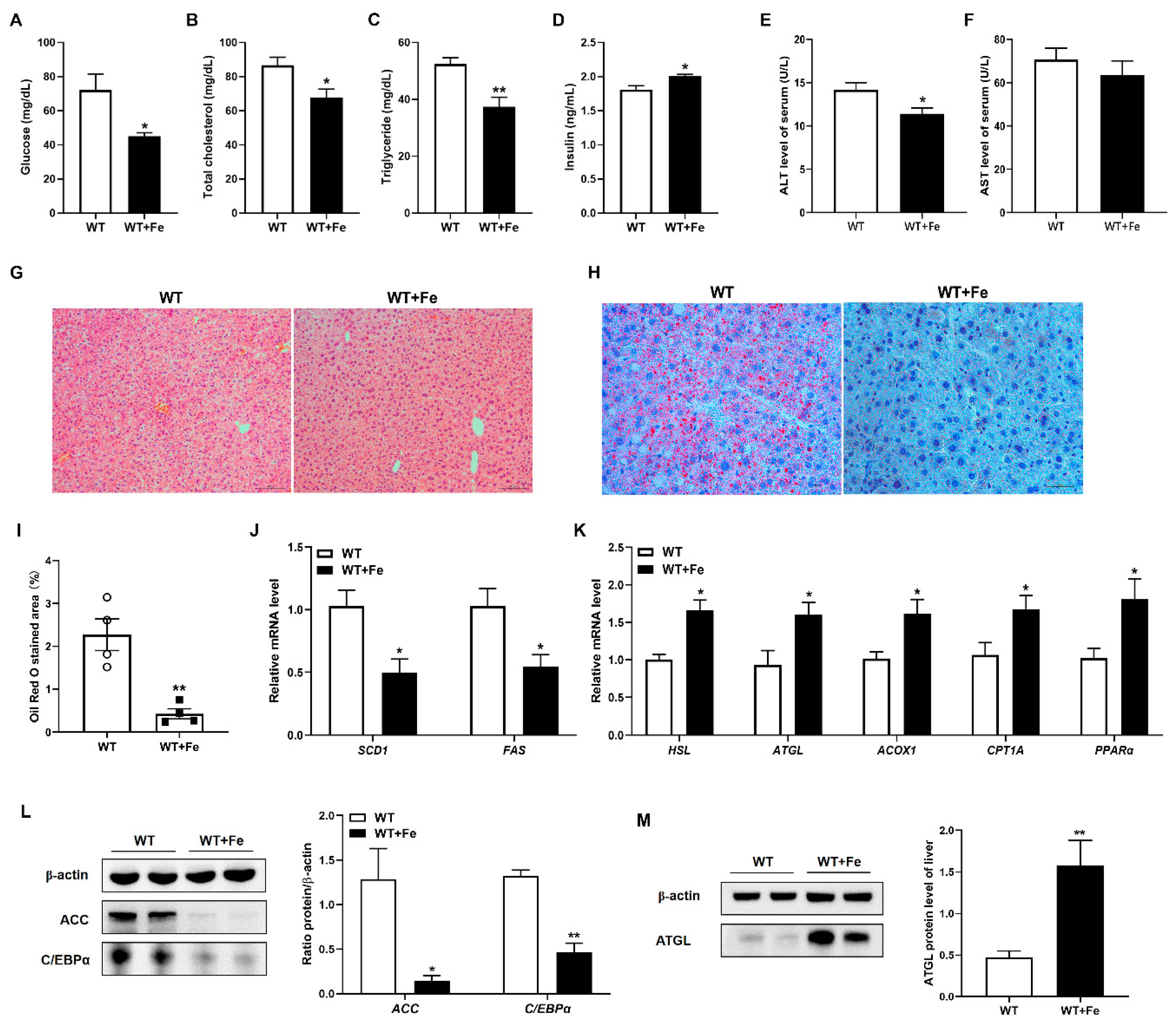
3.2. A High-Iron Diet Inhibited Adipogenesis and Promoted Lipolysis in Liver
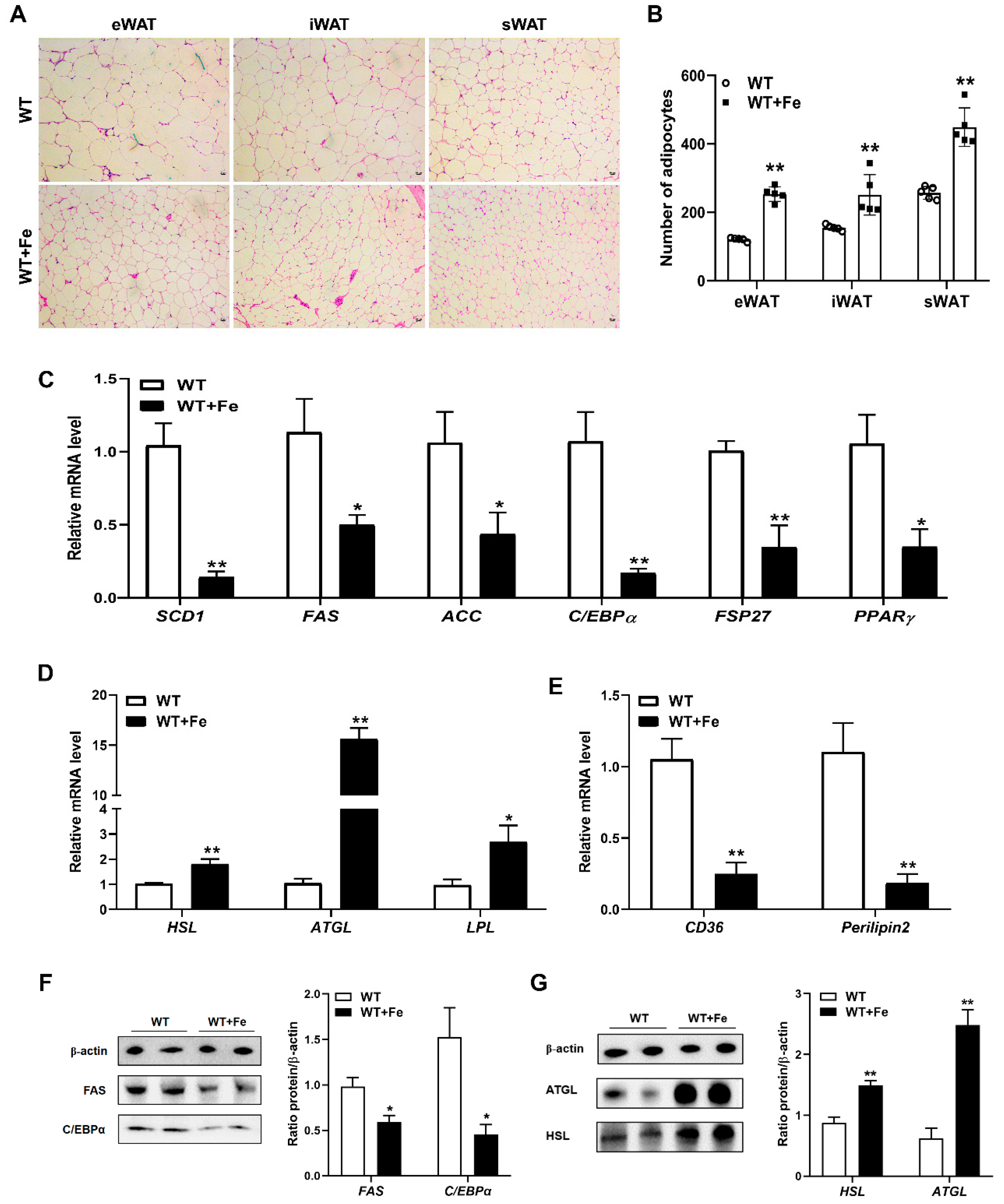
3.3. A High-Iron Diet Inhibited Adipogenesis and Promoted Lipolysis in Adipose Tissues
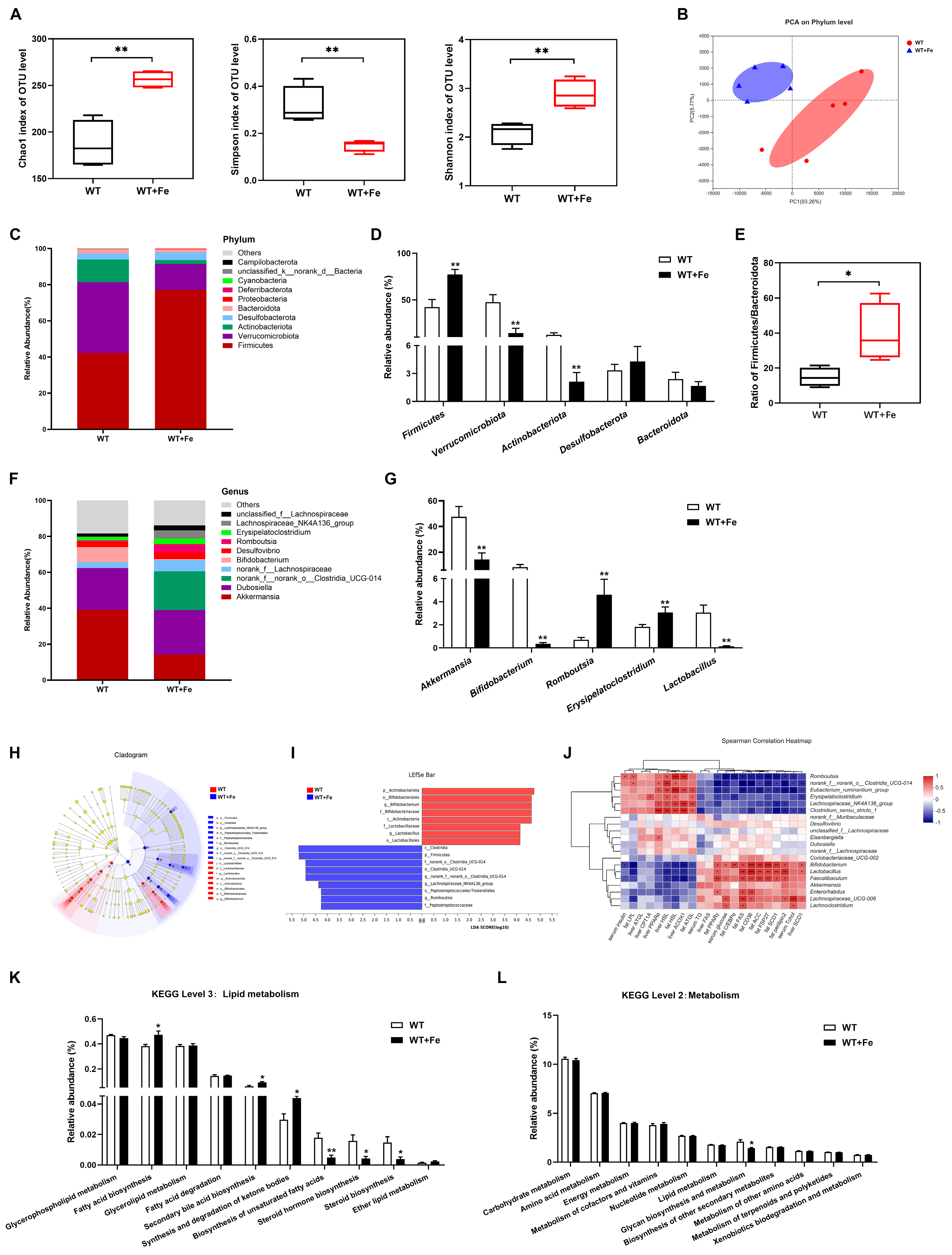
3.4. A High-Iron Diet Reshaped the Gut Microbiota of Mice
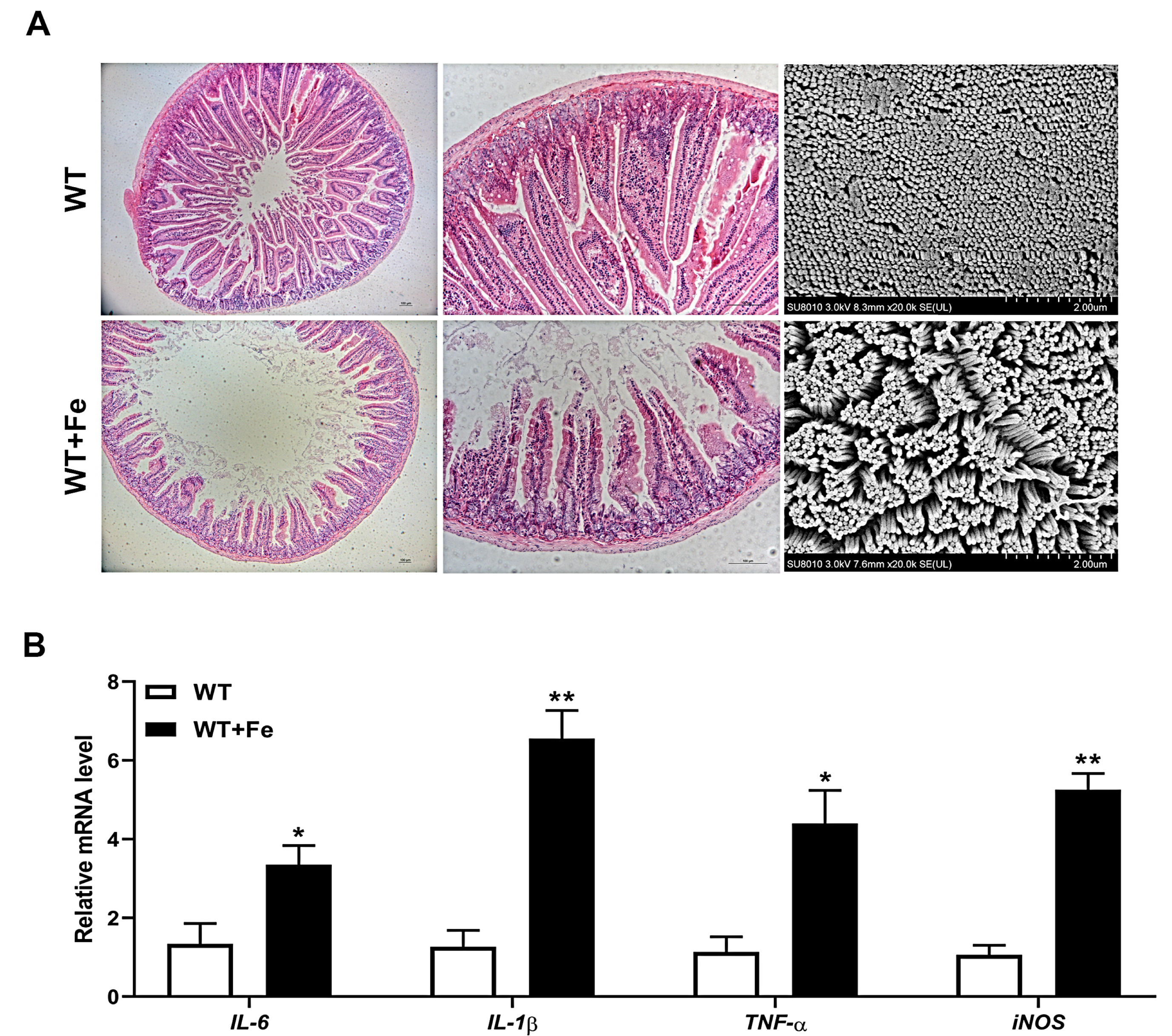
3.5. A High-Iron Diet Induced Intestinal Damage of Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yiannikourides, A.; Latunde-Dada, G. A Short Review of Iron Metabolism and Pathophysiology of Iron Disorders. Medicines 2019, 6, 85. [Google Scholar] [CrossRef]
- Hassan, T.H.; Badr, M.A.; Karam, N.A.; Zkaria, M.; El Saadany, H.F.; Rahman, D.M.A.; Shahbah, D.A.; Al Morshedy, S.M.; Fathy, M.; Esh, A.M.H.; et al. Impact of iron deficiency anemia on the function of the immune system in children. Medicine 2016, 95, e5395. [Google Scholar] [CrossRef]
- Cerami, C. Iron Nutriture of the Fetus, Neonate, Infant, and Child. Ann. Nutr. Metab. 2017, 71, 8–14. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, C.; Vrhovnik, P.; González-Alzaga, B.; Fernández, M.F.; Martin-Olmedo, P.; Olea, N.; Fiket, Ž.; Kniewald, G.; Arrebola, J.P. Socio-demographic, lifestyle, and dietary determinants of essential and possibly-essential trace element levels in adipose tissue from an adult cohort. Environ. Pollut. 2018, 236, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Gotardo, E.M.F.; Caria, C.R.E.P.; de Oliveira, C.C.; Rocha, T.; Ribeiro, M.; Gambero, A. Effects of iron supplementation in mice with hypoferremia induced by obesity. Exp. Biol. Med. 2016, 241, 2049–2055. [Google Scholar] [CrossRef] [PubMed]
- Dongiovanni, P.; Ruscica, M.; Rametta, R.; Recalcati, S.; Steffani, L.; Gatti, S.; Girelli, D.; Cairo, G.; Magni, P.; Fargion, S.; et al. Dietary Iron Overload Induces Visceral Adipose Tissue Insulin Resistance. Am. J. Pathol. 2013, 182, 2254–2263. [Google Scholar] [CrossRef]
- Ma, W.; Feng, Y.; Jia, L.; Li, S.; Li, J.; Wang, Z.; Chen, X.; Du, H. Dietary Iron Modulates Glucose and Lipid Homeostasis in Diabetic Mice. Biol. Trace Element Res. 2018, 189, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.S.; Koller, K.R.; Ramaboli, M.C.; Nesengani, L.T.; Ocvirk, S.; Chen, C.; Flanagan, C.A.; Sapp, F.R.; Merritt, Z.T.; Bhatti, F.; et al. Diet and the Human Gut Microbiome: An International Review. Dig. Dis. Sci. 2020, 65, 723–740. [Google Scholar] [CrossRef]
- Cherayil, B.J.; Ellenbogen, S.; Shanmugam, N.N. Iron and intestinal immunity. Curr. Opin. Gastroenterol. 2011, 27, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S.C.; Robinson, A.K.; Rodríguez-Quiñones, F. Bacterial iron homeostasis. FEMS Microbiol. Rev. 2003, 27, 215–237. [Google Scholar] [CrossRef]
- Ng, O. Iron, microbiota and colorectal cancer. Wien. Med. Wochenschr. 2016, 166, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Jaeggi, T.; Kortman, G.A.M.; Moretti, D.; Chassard, C.; Holding, P.; Dostal, A.; Boekhorst, J.; Timmerman, H.M.; Swinkels, D.W.; Tjalsma, H.; et al. Iron fortification adversely affects the gut microbiome, increases pathogen abundance and induces intestinal inflammation in Kenyan infants. Gut 2014, 64, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Mayneris-Perxachs, J.; Cardellini, M.; Hoyles, L.; Latorre, J.; Davato, F.; Moreno-Navarrete, J.M.; Arnoriaga-Rodríguez, M.; Serino, M.; Abbott, J.; Barton, R.H.; et al. Iron status influences non-alcoholic fatty liver disease in obesity through the gut microbiome. Microbiome 2021, 9, 104. [Google Scholar] [CrossRef] [PubMed]
- Kadis, S.; A Udeze, F.; Polanco, J.; Dreesen, D.W. Relationship of iron administration to susceptibility of newborn pigs to enterotoxic colibacillosis. Am. J. Vet.-Res. 1984, 45, 255–259. [Google Scholar]
- Knight, C.D.; Klasing, K.C.; Forsyth, D.M.E. Coli Growth in Serum of Iron Dextran-Supplemented Pigs. J. Anim. Sci. 1983, 57, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Gan, Z.; Wang, Z.; Tang, X.; Guo, B.; Du, H. Increased Iron Availability Aggravates the Infection of Escherichia coli O157:H7 in Mice. Biol. Trace Element Res. 2018, 190, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Chooi, Y.C.; Ding, C.; Magkos, F. The epidemiology of obesity. Metabolism 2019, 92, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, X.; Shen, Y.; Fang, X.; Wang, Y.; Wang, F. Obesity and iron deficiency: A quantitative meta-analysis. Obes. Rev. 2015, 16, 1081–1093. [Google Scholar] [CrossRef]
- Kell, D.B. Iron behaving badly: Inappropriate iron chelation as a major contributor to the aetiology of vascular and other progressive inflammatory and degenerative diseases. BMC Med Genom. 2009, 2, 2. [Google Scholar] [CrossRef]
- Ahmed, U. Interactions between hepatic iron and lipid metabolism with possible relevance to steatohepatitis. World J. Gastroenterol. 2012, 18, 4651–4658. [Google Scholar] [CrossRef] [PubMed]
- Attie, A.D.; Scherer, P.E. Adipocyte metabolism and obesity. J. Lipid Res. 2009, 50, S395–S399. [Google Scholar] [CrossRef] [PubMed]
- Björntorp, P. Effects of age, sex, and clinical conditions on adipose tissue cellularity in man. Metabolism 1974, 23, 1091–1102. [Google Scholar] [CrossRef]
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Naslund, E.; Britton, T.; et al. Dynamics of fat cell turnover in humans. Nature 2008, 453, 783–787. [Google Scholar] [CrossRef]
- Sims, E.A.; Danforth, E., Jr.; Horton, E.S.; Glennon, J.A.; Bray, G.A.; Salans, L.B. Experimental obesity in man. A progress report. Isr. J. Med. Sci. 1972, 8, 813–814. [Google Scholar] [PubMed]
- Danesch, U.; Hoeck, W.; Ringold, G.M. Cloning and transcriptional regulation of a novel adipocyte-specific gene, FSP27. CAAT-enhancer-binding protein (C/EBP) and C/EBP-like proteins interact with sequences required for differentiation-dependent expression. J. Biol. Chem. 1992, 267, 7185–7193. [Google Scholar] [CrossRef]
- Lefterova, M.I.; Haakonsson, A.K.; Lazar, M.A.; Mandrup, S. PPARγ and the global map of adipogenesis and beyond. Trends Endocrinol. Metab. 2014, 25, 293–302. [Google Scholar] [CrossRef]
- Strable, M.S.; Ntambi, J.M. Genetic control ofde novolipogenesis: Role in diet-induced obesity. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 199–214. [Google Scholar] [CrossRef]
- Glatz, J.F.; Luiken, J.J. From fat to FAT (CD36/SR-B2): Understanding the regulation of cellular fatty acid uptake. Biochimie 2017, 136, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Conte, M.; Franceschi, C.; Sandri, M.; Salvioli, S. Perilipin 2 and Age-Related Metabolic Diseases: A New Perspective. Trends Endocrinol. Metab. 2016, 27, 893–903. [Google Scholar] [CrossRef]
- Nielsen, T.S.; Jessen, N.; Jørgensen, J.O.L.; Møller, N.; Lund, S. Dissecting adipose tissue lipolysis: Molecular regulation and implications for metabolic disease. J. Mol. Endocrinol. 2014, 52, R199–R222. [Google Scholar] [CrossRef] [PubMed]
- Setoyama, D.; Fujimura, Y.; Miura, D. Metabolomics reveals that carnitine palmitoyltransferase-1 is a novel target for oxidative inactivation in human cells. Genes Cells 2013, 18, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Kersten, S.; Stienstra, R. The role and regulation of the peroxisome proliferator activated receptor alpha in human liver. Biochimie 2017, 136, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Jia, L.; Xiong, Q.; Du, H. Iron Overload Protects from Obesity by Ferroptosis. Foods 2021, 10, 1787. [Google Scholar] [CrossRef]
- Chen, G.-H.; Luo, Z.; Chen, F.; Shi, X.; Song, Y.-F.; You, W.-J.; Liu, X. PPARα, PPARγ and SREBP-1 pathways mediated waterborne iron (Fe)-induced reduction in hepatic lipid deposition of javelin goby Synechogobius hasta. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 197, 8–18. [Google Scholar] [CrossRef]
- Dao, M.C.; Clément, K. Gut microbiota and obesity: Concepts relevant to clinical care. Eur. J. Intern. Med. 2018, 48, 18–24. [Google Scholar] [CrossRef]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Fan, L.; Qi, Y.; Xu, C.; Hu, Y.; Chen, S.; Liu, W.; Liu, W.; Si, J. Akkermansia muciniphila Alleviates Dextran Sulfate Sodium (DSS)-Induced Acute Colitis by NLRP3 Activation. Microbiol. Spectr. 2021, 9, e00730-21. [Google Scholar] [CrossRef] [PubMed]
- Macchione, I.G.; Lopetuso, L.R.; Ianiro, G.; Napoli, M.; Gibiino, G.; Rizzatti, G.; Petito, V.; Gasbarrini, A.; Scaldaferri, F. Akkermansia muciniphila: Key player in metabolic and gastrointestinal disorders. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8075–8083. [Google Scholar] [PubMed]
- Binda, C.; Lopetuso, L.R.; Rizzatti, G.; Gibiino, G.; Cennamo, V.; Gasbarrini, A. Actinobacteria: A relevant minority for the maintenance of gut homeostasis. Dig. Liver Dis. 2018, 50, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Yun, J.M.; Kim, M.K.; Kwon, O.; Cho, B. Lactobacillus gasseri BNR17 Supplementation Reduces the Visceral Fat Accumulation and Waist Circumference in Obese Adults: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Med. Food 2018, 21, 454–461. [Google Scholar] [CrossRef]
- Crovesy, L.; Ostrowski, M.; Ferreira, D.M.T.P.; Rosado, E.L.; Soares, M. Effect of Lactobacillus on body weight and body fat in overweight subjects: A systematic review of randomized controlled clinical trials. Int. J. Obes. 2017, 41, 1607–1614. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.-M.; Huang, H.-L.; Liu, Y.-D.; Zhu, J.-Q.; Zhou, Y.-L.; Chen, H.-T.; Xu, J.; Zhao, H.-L.; Guo, X.; Shi, W.; et al. Selection strategy of dextran sulfate sodium-induced acute or chronic colitis mouse models based on gut microbial profile. BMC Microbiol. 2021, 21, 279. [Google Scholar] [CrossRef]
- Burke, K.E.; Lamont, J.T. Clostridium difficileInfection: A Worldwide Disease. Gut Liver 2014, 8, 1–6. [Google Scholar] [CrossRef]
- Ohtani, K.; Shimizu, T. Regulation of Toxin Production in Clostridium perfringens. Toxins 2016, 8, 207. [Google Scholar] [CrossRef]
- Kortman, G.A.; Raffatellu, M.; Swinkels, D.W.; Tjalsma, H. Nutritional iron turned inside out: Intestinal stress from a gut microbial perspective. FEMS Microbiol. Rev. 2014, 38, 1202–1234. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Wu, H.; Zeng, M.; Yu, G.; Dong, S.; Yang, H. Probiotic/prebiotic correction for adverse effects of iron fortification on intestinal resistance to Salmonella infection in weaning mice. Food Funct. 2018, 9, 1070–1078. [Google Scholar] [CrossRef] [PubMed]
- Jahani-Sherafat, S.; Alebouyeh, M.; Moghim, S.; Amoli, H.A.; Safaei, H.G. Role of gut microbiota in the pathogenesis of colorectal cancer; a review article. Gastroenterol. Hepatol. Bed Bench 2018, 11, 101–109. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| ACC | CTGTATGAGAAAGGCTATGTG | AACCTGTCTGAAGAGGTTAG |
| ACOX1 | TGTCTCGCTCCGCTCATAGG | ACATGGAGTAATTGAGGCCAACA |
| ATGL | ACTGAACCAACCCAACCCTTT | CCCGTCTGCTCTTTCATCCA |
| C/EBPα | CAAGAACAGCAACGAGTACCG | GTCACTCGTCAACTCCAGCAC |
| CD36 | ATTCCCTTGGCAACCAACCA | CGTGGCCCGGTTCTAATTCA |
| CPT1A | GGGAGGAATACATCTACCTG | GAAGACGAATAGGTTTGAG |
| FAS | GATTCAGGGAGTGGATATTG | CATTCAGAATCGTGGCATAG |
| FPN | ATGGGAACTGTGGCCTTCAC | TCCAGGCATGAATACGGAGA |
| FSP27 | ATGGACTACGCCATGAAGTCT | CGGTGCTAACACGACAGGG |
| FtH | TGGAACTGCACAAACTGGCTACT | ATGGATTTCACCTGTTCACTCAGATAA |
| FtL | CGTGGA TCTGTGTCTTGCTT | GCGAAGAGACGGTGCAGACT |
| GAPDH | CGGGAAACTGTGGCGTGATG | CAAAGGTGGAGGAGTGGG |
| Hepcidin | CTGCCTGTCTCCTGCTTCT | ATGTCTGCCCTGCTTTCTT |
| HSL | GGAGCTCCAGTCGGAAGAGG | GTCTTCTGCGAGTGTCACCA |
| IL-1β | AGTTGACGGACCCCAAAAG | TTTGAAGCTGGATGCTCTCAT |
| IL-6 | CCCCAATTTCCAATGCTCTCC | CGCACTAGGTTTGCCGAGT |
| iNOS | CTCACCTACTTCCTGGACATTAC | CAATCTCTGCCTATCCGTCTC |
| LPL | TCGCCTTTCTCCTGATGACG | GCAATCACACGGATGGCTTC |
| Perilipin2 | GACACCACCTGCATGGCT | TGAAGCAGGGCCACTCTC |
| PPARα | AGTTCGGGAACAAGACGTTG | CAGTGGGGAGAGAGGACAGA |
| PPARγ | GAAGCGGTGAACCACTGAT | GGAATGCGAGTGGTCTTCCA |
| SCD1 | GTGGGGTAATTATTTGTGACC | TTTTTCCCAGACAGTACAAC |
| TNF-α | GCTCTTCTGTCTACTGAACTTCGG | ATGATCTGAGTGTGAGGGTCTGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, Q.; Zhao, J.; Tian, C.; Ma, W.; Miao, L.; Liang, L.; Zhang, K.; Du, H. Regulation of a High-Iron Diet on Lipid Metabolism and Gut Microbiota in Mice. Animals 2022, 12, 2063. https://doi.org/10.3390/ani12162063
Xiong Q, Zhao J, Tian C, Ma W, Miao L, Liang L, Zhang K, Du H. Regulation of a High-Iron Diet on Lipid Metabolism and Gut Microbiota in Mice. Animals. 2022; 12(16):2063. https://doi.org/10.3390/ani12162063
Chicago/Turabian StyleXiong, Qingqing, Jing Zhao, Chenying Tian, Wan Ma, Linfeng Miao, Li Liang, Kang Zhang, and Huahua Du. 2022. "Regulation of a High-Iron Diet on Lipid Metabolism and Gut Microbiota in Mice" Animals 12, no. 16: 2063. https://doi.org/10.3390/ani12162063
APA StyleXiong, Q., Zhao, J., Tian, C., Ma, W., Miao, L., Liang, L., Zhang, K., & Du, H. (2022). Regulation of a High-Iron Diet on Lipid Metabolism and Gut Microbiota in Mice. Animals, 12(16), 2063. https://doi.org/10.3390/ani12162063