Comparison of Nutritive Values of Tropical Pasture Species Grown in Different Environments, and Implications for Livestock Methane Production: A Meta-Analysis

 , , ,
, , ,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
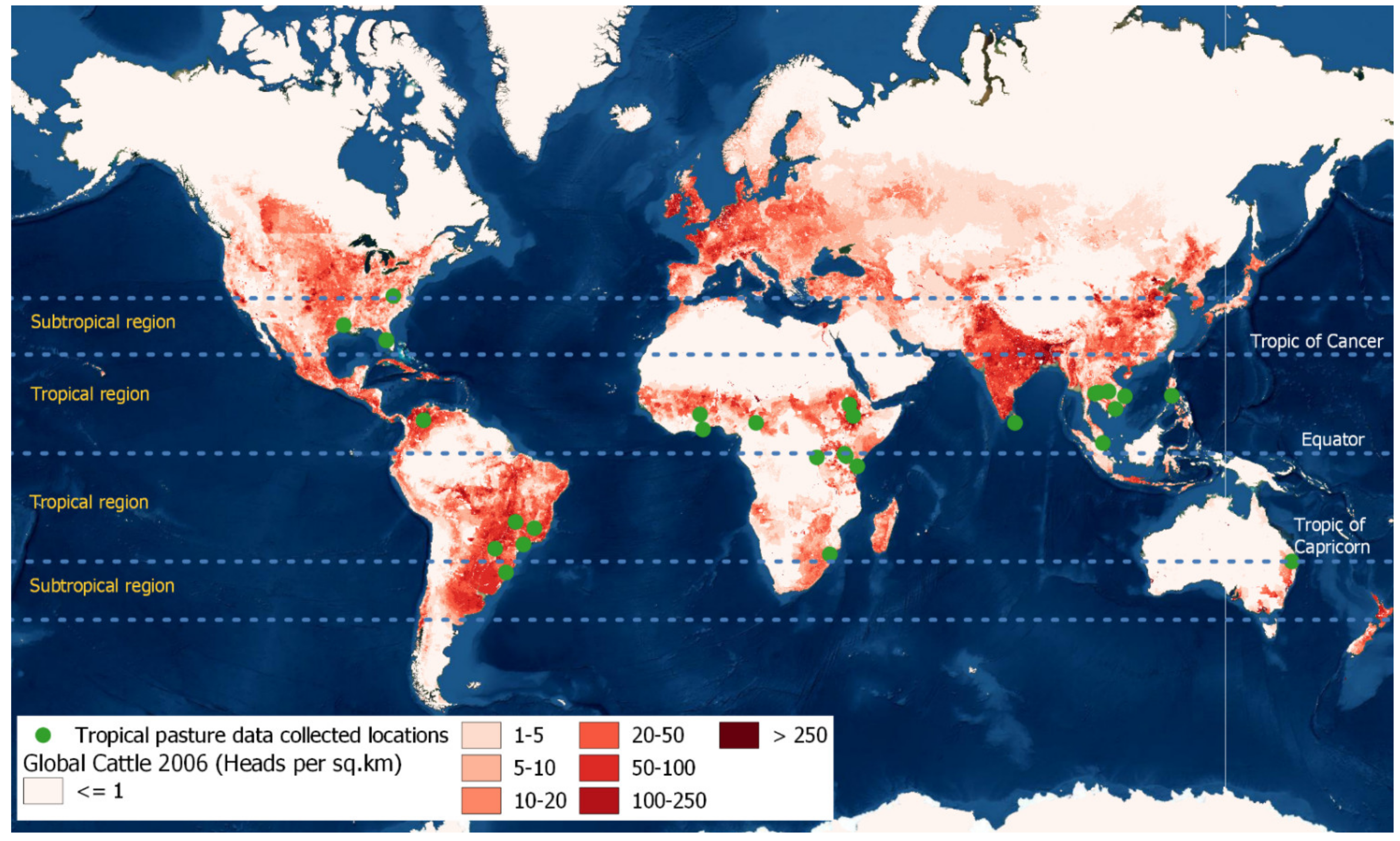
2.1. Data Collection
2.2. Data Processing and Analyses
3. Results
3.1. Description of the Analytical Database
3.2. Comparisons of the Nutritive Values
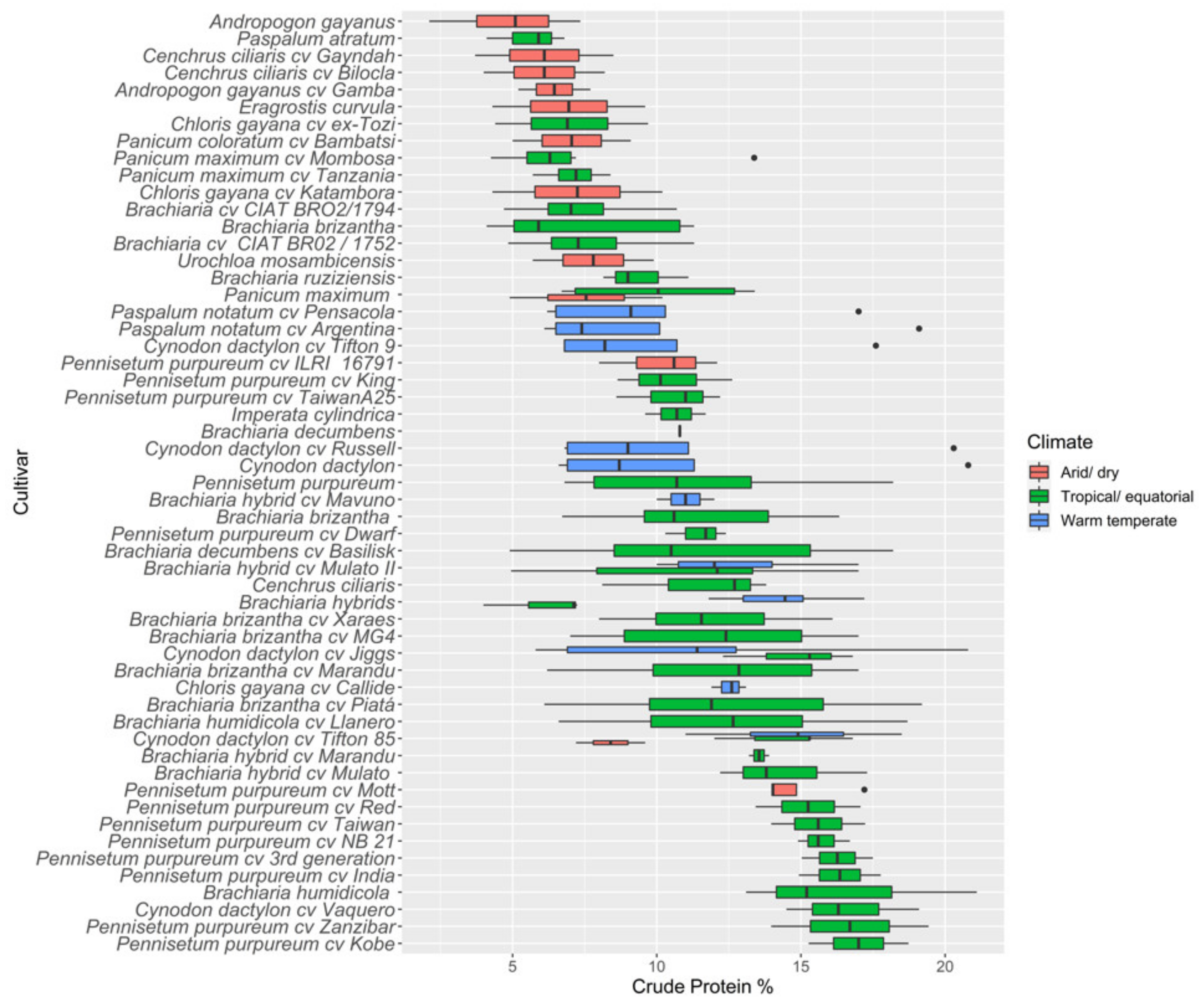
3.2.1. Crude Protein
3.2.2. Fibre
3.2.3. Digestibility
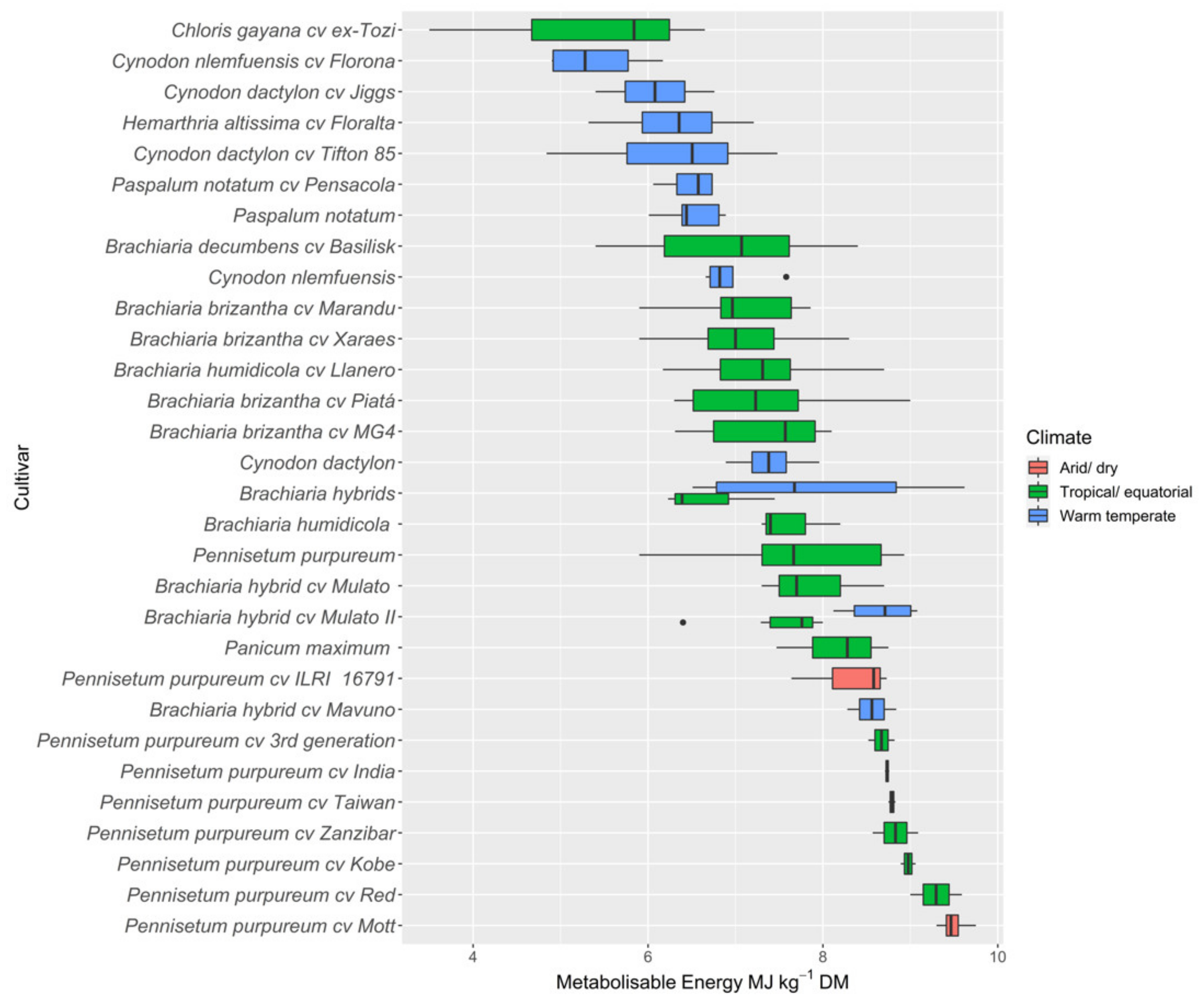
3.2.4. Metabolisable Energy
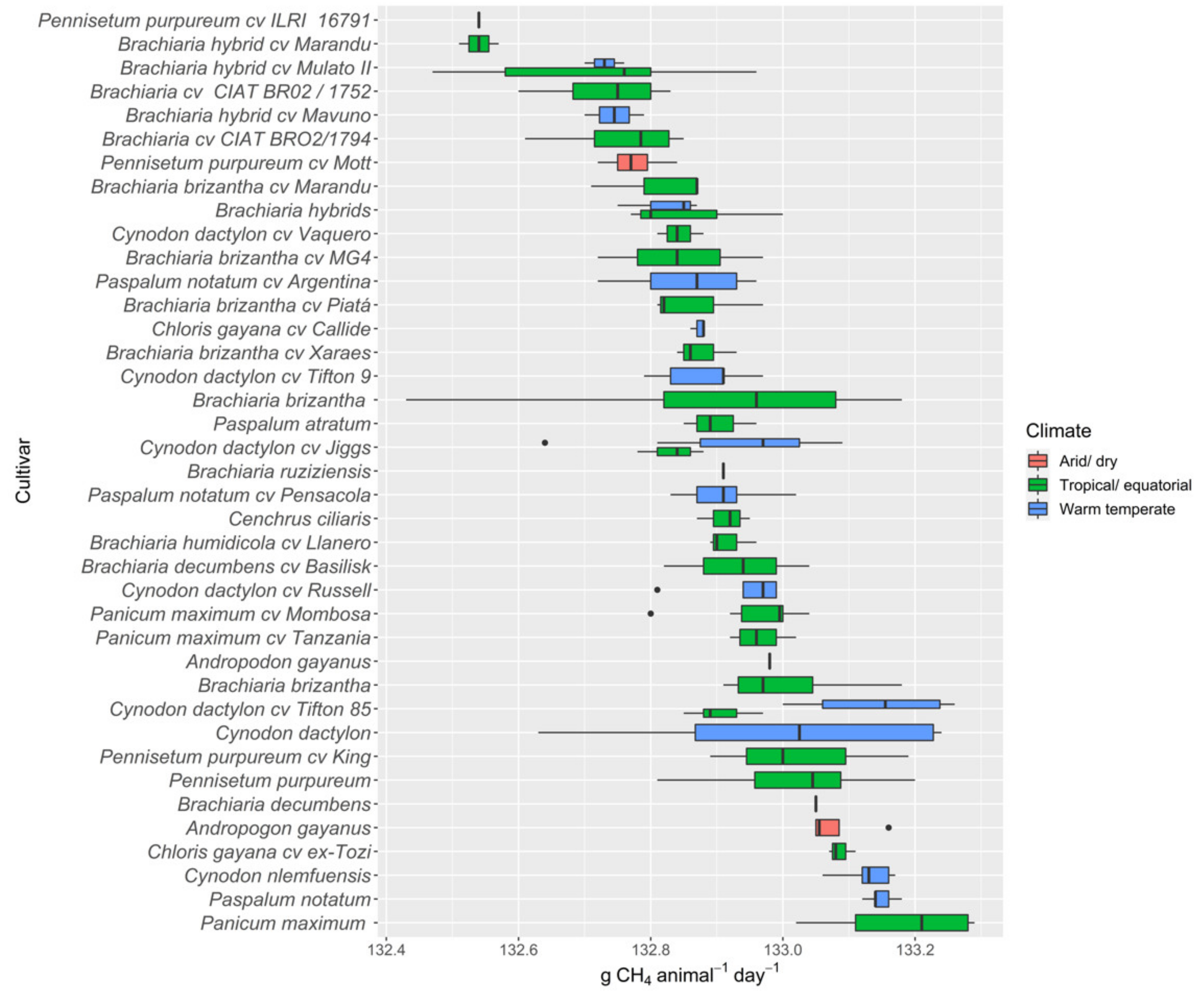
3.2.5. Methane Gas
3.3. Bioclimatic Variations in Nutritive Metrics
3.4. Management and Environmental Determinants
4. Discussion
4.1. Nutritive Value of Tropical Pastures
4.2. Bioclimatic Variation
4.3. Management Determinants
4.4. Implications for Livestock Methane Production
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Opio, C.; Gerber, P.; Mottet, A.; Falcucci, A.; Tempio, G.; MacLeod, M.; Vellinga, T.; Henderson, B.; Steinfeld, H. Greenhouse Gas Emissions from Ruminant Supply Chains—A Global Life Cycle Assessment; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013. [Google Scholar]
- FAO Stat. FAOSTAT Statistical Database 2018. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 20 May 2021).
- FAO (Ed.) Mitigation of Greenhouse Gas Emissions in Livestock Production: A Review of Technical Options for Non CO2 Emissions; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; p. 226. [Google Scholar]
- Thornton, P.K.; Herrero, M. Potential for reduced methane and carbon dioxide emissions from livestock and pasture management in the tropics. Proc. Natl. Acad. Sci. USA 2010, 107, 19667–19672. [Google Scholar] [CrossRef]
- EPA. DRAFT: Global Anthropogenic Non-CO2 Greenhouse Gas Emissions 1990–2030; Publication 430-D-11-003; EPA: Washington, DC, USA, 2011. [Google Scholar]
- Smith, P.; Bustamante, M.; Ahammad, H. Climate Change 2014: Mitigation of Climate Change. Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 811–922. [Google Scholar]
- Thornton, P.K.; Jones, P.G.; Ericksen, P.J.; Challinor, A.J. Agriculture and food systems in sub-Saharan Africa in a in a +4 °C world. Philos. Trans. R. Soc. A 2011, 369, 117–136. [Google Scholar] [CrossRef]
- Boval, M.; Edouard, N.; Sauvant, D. A meta-analysis of nutrient intake, feed efficiency and performance in cattle grazing on tropical grasslands. Anim. Consort. 2015, 9, 973–983. [Google Scholar] [CrossRef]
- Capstaff, N.M.; Miller, A.J. Improving the yield and nutritional quality of forage crops. Front. Plant Sci. 2018, 9, 535. [Google Scholar] [CrossRef]
- Paul, B.K.; Koge, J.; Maass, B.L.; Notenbaert, A.; Peters, M.; Groot, J.C.J.; Tittonell, P. Tropical forage technologies can deliver multiple benefits in Sub-Saharan Africa. A meta-analysis. Agron. Sustain. Dev. 2020, 40, 22. [Google Scholar] [CrossRef]
- Boga, M.; Ayasan, T. Determination of Nutritional Value of Alfalfa Varieties and Lines by Using the In Vitro Method and Gas Production Technique. J. Hell. Vet. Med. Soc. 2022, 73, 3613–3620. [Google Scholar] [CrossRef]
- Congio, G.F.S.; Batalha, C.D.A.; Chiavegato, M.B.; Berndt, A.; Oliveira, P.P.A.; Frighetto, R.T.S.; Maxwell, T.M.R.; Gregorini, P.; Da Silva, S.C. Strategic grazing management towards sustainable intensification at tropical pasture-based dairy systems. Sci. Total Environ. 2018, 636, 872–880. [Google Scholar] [CrossRef]
- Siqueira da Silva, H.M.; Vendramini, J.M.B.; Leite de Oliveira, F.C.; Soares Filho, C.V.; Kaneko, M.; Silveira, M.L.; Dalmazo Sanchez, J.M.; Yarborough, J.K. Harvest frequency effects on herbage characteristics of ‘Mavuno’ brachiariagrass. Crop Sci. 2020, 60, 1113–1122. [Google Scholar] [CrossRef]
- Inyang, U.; Vendramini, J.M.B.; Sellers, B.; Silveira, M.L.A.; Lunpha, A.; Sollenberger, L.E.; Adesogan, A.; Paiva, L.M. Harvest frequency and stubble height affect herbage accumulation, nutritive value, and persistence of ‘Mulato II’ Brachiariagrass. Forage Grazinglands 2010, 8, 1–7. [Google Scholar] [CrossRef]
- Moyo, M.; Nsahlai, I. Consequences of increases in ambient temperature and effect of climate type on digestibility of forages by ruminants: A meta-analysis in relation to global warming. Animals 2021, 11, 172. [Google Scholar] [CrossRef]
- Pontes, L.d.S.; Baldissera, T.C.; Giostri, A.F.; Stafin, G.; dos Santos, B.R.C.; Carvalho, P.C.d.F. Effects of nitrogen fertilization and cutting intensity on the agronomic performance of warm-season grasses. Grass Forage Sci. 2017, 72, 663–675. [Google Scholar] [CrossRef]
- Woodard, K.R.; Sollenberger, L.E. Broiler litter vs. ammoniumnitrate as N source for bermudagrass hay production: Yield, nutritive value, and nitrate leaching. Crop Sci. 2011, 51, 1342–1352. [Google Scholar] [CrossRef]
- Gastal, F.; Lemaire, G. Defoliation, shoot plasticity, sward structure and herbage utilization in pasture: Review of the underlying ecophysiological processes. Agriculture 2015, 5, 1146–1171. [Google Scholar] [CrossRef]
- Kering, M.K.; Guretzky, J.; Funderburg, E.; Mosali, J. Effect of nitrogen fertilizer rate and harvest season on forage yield, quality, and macronutrient concentrations in midland Bermuda Grass. Commun. Soil Sci. Plant Anal. 2011, 42, 1958–1971. [Google Scholar] [CrossRef]
- Gardarin, A.; Garnier, É.; Carrère, P.; Cruz, P.; Andueza, D.; Bonis, A.; Colace, M.P.; Dumont, B.; Duru, M.; Farruggia, A. Plant trait-digestibility relationships across management and climate gradients in permanent grasslands. J. Appl. Ecol. 2014, 51, 1207–1217. [Google Scholar] [CrossRef]
- Hoskin, S.O.; Gee, E.K. Feeding value of pastures for horses. N. Z. Vet. J. 2004, 52, 332–341. [Google Scholar] [CrossRef]
- Herrero, M.; Wirsenius, S.; Henderson, B.; Rigolot, C.; Thornton, P.; Havlík, P.; Boer, I.d.; Gerber, P.J. Livestock and the Environment: What have we learned in the past decade? Annu. Rev. Environ. Resour. 2015, 40, 177–202. [Google Scholar] [CrossRef]
- Knapp, J.R.; Laur, G.L.; Vadas, P.A.; Weiss, W.P.; Tricarico, J.M. Invited review: Enteric methane in dairy cattle production: Quantifying the opportunities and impact of reducing emissions. J. Dairy Sci. 2014, 97, 3231–3261. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Moran, J. Tropical Dairy Farming: Feeding Management for Small Holder Dairy Farmers in the Humid Tropics; CSIRO Publishing: Collingwood, VIC, Australia, 2005. [Google Scholar]
- Benaouda, M.; González-Ronquillo, M.; Appuhamy, J.A.D.R.N.; Kebreab, E.; Molina, L.T.; Herrera-Camacho, J.; Ku-Vera, J.C.; Ángeles-Hernández, J.C.; Castelán-Ortega, O.A. Development of mathematical models to predict enteric methane emission by cattle in Latin America. Livest. Sci. 2020, 241, 104177. [Google Scholar] [CrossRef]
- Lee, M.A.; Davis, A.P.; Chagunda, M.G.G.; Manning, P. Forage quality declines with rising temperatures, with implications for livestock production and methane emissions. Biogeosciences 2017, 14, 1403–1417. [Google Scholar] [CrossRef]
- Patra, A.K. Prediction of enteric methane emission from cattle using linear and non-linear statistical models in tropical production systems. Mitig. Adapt. Strateg. Glob. Chang. 2017, 22, 629–650. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv, 2015; arXiv:1406.5823. [Google Scholar] [CrossRef]
- RCoreTeam. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Gilbert, M.; Nicolas, G.; Cinardi, G.; Van Boeckel, T.P.; Vanwambeke, S.; Wint, W.G.R.; Robinson, T.P. Global cattle distribution in 2010 (5 min of arc). Harv. Dataverse 2018. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Pro 2.8.6; Environmental Systems Research Institute: Redlands, CA, USA, 2021. [Google Scholar]
- Lean, I.J.; Golder, H.M.; Hall, M.B. Feeding, evaluating, and controlling rumen function. Vet. Clin. North Am. Food Anim. Pract. 2014, 30, 539–575. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Dairy Cattle: Seventh Revised Edition; The National Academies Press: Washington, DC, USA, 2001; p. 405. [Google Scholar]
- Van Soest, P.J. Nutritional Ecology of the Ruminant; Cornell University Press: Ithaca, NY, USA, 1994. [Google Scholar]
- Katoch, R. Factors influencing forage nutritional quality. In Nutritional Quality Management of Forages in the Himalayan Region; Springer: Singapore, 2022; pp. 99–149. [Google Scholar]
- Givens, D.I. Forage Evaluation in Ruminant Nutrition; CABI Books: London, UK, 2000. [Google Scholar]
- Ayele, S.; Duncan, A.; Larbi, A.; Khanh, T.T. Enhancing innovation in livestock value chains through networks: Lessons from fodder innovation case studies in developing countries. Sci. Public Policy 2012, 39, 333–346. [Google Scholar] [CrossRef]
- Hall, A.; Sulaiman, R.; Bezkorowajnyj, P. Reframing Technical Change: Livestock Fodder Scarcity Revisited as Innovation Capacity Scarcity—A Conceptual Framework; ILRI and UNU/MERIT: Nairobi, Kenya, 2007. [Google Scholar]
- Rao, I.M.; Peters, M.; Castro, A.; Schultze-Kraft, R.; White, D.; Fisher, M.; Miles, J.W.; Lascano, C.E.; Blümmel, M.; Bungenstab, D. LivestockPlus: The Sustainable Intensification of Forage-Based Agricultural Systems to Improve Livelihoods and Ecosystem Services in the Tropics; CIAT Publication: Cali, Colombia, 2015. [Google Scholar]
- Jégo, G.; Bélanger, G.; Tremblay, G.F.; Jing, Q.; Baron, V.S. Calibration and performance evaluation of the STICS crop model for simulating timothy growth and nutritive value. Field Crops Res. 2013, 151, 65–77. [Google Scholar] [CrossRef]
- Lee, M.A. A global comparison of the nutritive values of forage plants grown in contrasting environments. J. Plant Res. 2018, 131, 641–654. [Google Scholar] [CrossRef]
- Chapman, D.F. Using ecophysiology to improve farm efficiency: Application in temperate dairy grazing systems. Agriculture 2016, 6, 17. [Google Scholar] [CrossRef]
- Lemaire, G.; Da Silva, S.C.; Agnusdei, M.; Wade, M.; Hodgson, J. Interactions between leaf lifespan and defoliation frequency in temperate and tropical pastures: A review. Grass Forage Sci. 2009, 64, 341–353. [Google Scholar] [CrossRef]
- Ruolo, M.S.; Pérez, H.E.; Rodriguez, A.M. Chloris gayana Kunth under different defoliation regimes. Morphogenesis, sward structure and leaf area index. Grass Forage Sci. 2019, 74, 720–727. [Google Scholar] [CrossRef]
- Hughes, M.P.; Mlambo, V.; Lallo, C.H.O. Optimum nitrogen fertilization rate and nitrogen use efficiency for Brachiaria hybrid and Megathyrsus maximus varies with stage of regrowth. JSFA Rep. 2022, 2, 168–177. [Google Scholar] [CrossRef]
- Tessema, Z.; Baars, R.; Alemu, Y.; Dawit, N. In Sacco dry matter and nitrogen degradation and their relationship with in vitro dry matter digestibility of Napier grass (Pennisetum purpureum Schumach) as influenced by plant height at cutting. Aust. J. Agric. Res. 2002, 53, 7–12. [Google Scholar]
- Vendramini, J.M.B.; Sollenberger, L.E.; Soares, A.B.; Da Silva, W.L.; Sanchez, J.M.; Valente, A.L.; Aguiar, A.D.; Mullenix, M.K. Harvest frequency affects herbage accumulation and nutritive value of Brachiaria grass hybrids in Florida. Trop. Grassl.-Forrajes Trop. 2014, 2, 197–206. [Google Scholar] [CrossRef]
- Tessema, Z.K.; Mihret, J.; Solomon, M. Effect of defoliation frequency and cutting height on growth, dry-matter yield and nutritive value of Napier grass (Pennisetum purpureum (L.) Schumach). Grass Forage Sci. 2010, 65, 421–430. [Google Scholar] [CrossRef]
- Arthington, J.D.; Brown, W.F. Estimation of feeding value of four tropical forage species at two stages of maturity. J. Anim. Sci. 2005, 83, 1726–1731. [Google Scholar] [CrossRef]
- Pembleton, K.G.; Lowe, K.F.; Bahnisch, M. Utilising leaf number as an indicator for defoliation to restrict stem growth in rhodes grass (Chloris gayana) cv. Callide. Trop. Grassl.-Forrajes Trop. 2009, 43, 79–85. [Google Scholar]
- Barbosa, R.A.; Nascimento Júnior, D.d.; Euclides, V.P.B.; Silva, S.C.d.; Zimmer, A.H.; Torres Júnior, R.A.d.A. Tanzânia grass subjected to combinations of intensity and frequency of grazing. Pesqui Agropecu Bras. 2007, 42, 329–340. [Google Scholar] [CrossRef]
- Benvenutti, M.A.; Pavetti, D.R.; Poppi, D.P.; Mayer, D.G.; Gordon, I.J. Ingestive behaviour and forage intake responses of young and mature steers to the vertical differentiation of sugarcane in pen and grazing studies. J Agric. Sci. 2017, 155, 1677–1688. [Google Scholar] [CrossRef][Green Version]
- Benvenutti, M.A.; Pavetti, D.R.; Poppi, D.P.; Gordon, I.J.; Cangiano, C.A. Defoliation patterns and their implications for the management of vegetative tropical pastures to control intake and diet quality by cattle. Grass Forage Sci. 2016, 71, 424–436. [Google Scholar] [CrossRef]
- Ison, K.; Barber, D.G.; Benvenutti, M.A.; Mayer, D.G.; Findsen, C. Abstracts from the 32nd Biennial Conference of the Australian Society of Animal Production (ASAP). Anim. Prod. Sci. 2018, 58, i–cxxxi. [Google Scholar] [CrossRef]
- Ungar, R. Bite horizons and dimensions for cattle grazing herbage to high levels of depletion. Grass Forage Sci. 1999, 54, 357–364. [Google Scholar] [CrossRef]
- Benvenutti, M.A.; Findsen, C.; Savian, J.V.; Mayer, D.G.; Barber, D.G. The effect of stage of regrowth on the physical composition and nutritive value of the various vertical strata of kikuyu (Cenchrus clandestinus) pastures. Trop. Grassl.-Forrajes Trop. 2020, 8, 141–146. [Google Scholar] [CrossRef]
- Ruggieri, A.C.; Cardoso, A.d.S.; Ongaratto, F.; Casagrande, D.R.; Barbero, R.P.; Brito, L.d.F.; Azenha, M.V.; Oliveira, A.A.; Koscheck, J.F.W.; Reis, R.A. Grazing intensity impacts on herbage mass, sward structure, greenhouse gas emissions, and animal performance: Analysis of Brachiaria pastureland. Agronomy 2020, 10, 1750. [Google Scholar] [CrossRef]
- Berça, A.S.; Cardoso, A.D.S.; Longhini, V.Z.; Tedeschi, L.O.; Boddey, R.M.; Berndt, A.; Reis, R.A.; Ruggieri, A.C. Methane production and nitrogen balance of dairy heifers grazing palisade grass cv. Marandu alone or with forage peanut. J. Anim. Sci. 2019, 97, 4625–4634. [Google Scholar] [CrossRef]
- Sauvant, D.; Giger-Reverdin, S. Modélisation des interactions digestives et de la production de méthane chez les ruminants. INRAE Prod. Anim. 2009, 22, 375–384. [Google Scholar] [CrossRef]
- Neto, A.J.; Messana, J.D.; Granja-Salcedo, Y.T.; Castagnino, P.S.; Fiorentini, G.; Reis, R.A.; Berchielli, T.T. Effect of starch level in supplement with or without oil source on diet and apparent digestibility, rumen fermentation and microbial population of Nellore steers grazing tropical grass. Livest. Sci. 2017, 202, 171–179. [Google Scholar] [CrossRef]
- Hegarty, R. Nutritional management options to reduce enteric methane emissions from NSW beef and dairy herds. In Proceedings of the 24th Annual Conference of the Grassland Society of NSW, Taree, NSW, Australia, 5–6 August 2009; pp. 40–47. [Google Scholar]
- Liu, Z.; Liu, Y.; Shi, X.; Wang, J.; Murphy, J.P.; Maghirang, R. Enteric methane conversion factor for dairy and beef cattle: Effects of feed digestibility and intake level. Trans. ASABE 2017, 60, 459–464. [Google Scholar] [CrossRef]
- Eggleston, H.S.; Buendia, L.; Miwa, K.; Ngara, T.; Tanabe, K. IPCC Guidelines for National Hreenhouse Gas Nventories; IPCC National Greenhouse Gas Inventories Programme, Intergovernmental Panel on Climate Change IPCC, c/o Institute for Global Environmental Strategies IGES, 2108—11, Kamiyamaguchi: Hayama, Japan, 2006. [Google Scholar]
- Chaves, A.V.; Thompson, L.C.; Iwaasa, A.D.; Scott, S.L.; Olson, M.E.; Benchaar, C.; Veira, D.M.; McAllister, T.A. Effect of pasture type (alfalfa vs. grass) on methane and carbon dioxide production by yearling beef heifers. Can. J. Anim. Sci. 2006, 86, 409–418. [Google Scholar] [CrossRef]
- Boadi, D.A.; Wittenberg, K.M.; McCaughey, W. Effects of grain supplementation on methane production of grazing steers using the sulphur (SF6) tracer gas technique. Can. J. Anim. Sci. 2002, 82, 151–157. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Kreuzer, M.; O’Mara, F.; McAllister, T.A. Nutritional management for enteric methane abatement: A review. Aust. J. Exp. Agric. 2008, 48, 21–27. [Google Scholar] [CrossRef]
- Meister, N.C.; Cardoso, A.d.S.; Alari, F.O.; Lemos, N.L.S.; Frighetto, R.T.S.; Malheiros, E.B.; Reis, R.A.; Ruggieri, A.C. Effect of pasture management on enteric methane emissions from goats. Trop. Anim. Health Prod. 2021, 53, 94. [Google Scholar] [CrossRef] [PubMed]
- De Souza Filho, W.; Nunes, P.A.d.A.; Barro, R.S.; Kunrath, T.R.; de Almeida, G.M.; Genro, T.C.M.; Bayer, C.; de Faccio Carvalho, P.C. Mitigation of enteric methane emissions through pasture management in integrated crop-livestock systems: Trade-offs between animal performance and environmental impacts. J. Clean. Prod. 2019, 213, 968–975. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | n | Mean | SD | Minimum | Maximum |
|---|---|---|---|---|---|
| Climate | |||||
| MAT (°C) | 294 | 22.8 | 3.45 | 17.5 | 29.30 |
| MAR (mm) | 294 | 1229.5 | 603.57 | 104.5 | 3390.0 |
| MSL (m) | 294 | 516.2 | 627.61 | 15.0 | 2393.0 |
| Defoliation frequency (days) | 294 | 59.51 | 35.68 | 14.0 | 180.0 |
| Defoliation intensity (mm) | 285 | 100.1 | 50.36 | 40.0 | 200.0 |
| Forage yield (DM t ha−1) | 188 | 7.84 | 6.75 | 0.33 | 46.01 |
| Nutritive metrics (DM) | |||||
| NDF (%) | 186 | 67.32 | 5.73 | 50.91 | 79.80 |
| ADF (%) | 160 | 38.85 | 5.86 | 24.70 | 57.44 |
| ADL (%) | 30 | 5.69 | 1.42 | 2.83 | 8.20 |
| Ash (%) | 88 | 8.75 | 3.18 | 4.40 | 15.20 |
| CP (%) | 254 | 10.97 | 4.08 | 2.11 | 21.10 |
| DMD (%) | 49 | 56.01 | 7.13 | 30.20 | 70.10 |
| OMD (%) | 145 | 57.99 | 7.30 | 33.11 | 72.20 |
| ME (MJkg−1 DM) | 136 | 7.41 | 1.16 | 3.50 | 9.75 |
| OM (gkg−1) | 43 | 894.1 | 81.93 | 590.00 | 957.00 |
| CH4 production | |||||
| 1 CH4 (g CH4 animal−1 day−1) | 186 | 132.92 | 0.17 | 132.43 | 133.30 |
| Nutritive Metrics | n | Climate | p-Value | ||
|---|---|---|---|---|---|
| Arid/Dry | Tropical/ Equatorial | Warm Temperate | |||
| CP (%) | 245 | 7.15 b ± 0.97 | 11.25 a ± 0.42 | 12.02 a ± 0.76 | <0.0001 |
| NDF (%) | 175 | 63.64 ± 2.40 | 67.47 ± 0.87 | 68.50 ± 1.21 | 0.1989 |
| ADF (%) | 164 | 37.82 ± 2.27 ab | 40.97 a ± 0.82 | 35.21 ± 1.36 b | <0.0001 |
| OMD (%) | 142 | - | 58.76 ± 1.14 | 56.17 ± 1.57 | 0.1845 |
| DMD (%) | 53 | - | 54.37 ± 1.44 | 52.33 ± 3.60 | 0.6039 |
| ME (MJ kg−1 DM) | 142 | 8.67 a ± 0.55 | 7.60 b ± 0.18 | 7.19 b ± 0.25 | 0.0477 |
| ADL (%) | 34 | 4.58 ± 0.78 | 6.07 ± 0.42 | 4.50 ± 1.37 | 0.1913 |
| Ash (%) | 88 | 13.17 a ± 2.33 | 10.15 a ± 0.58 | 5.93 b ± 0.93 | <0.0001 |
| OM (g kg−1 DM) | 47 | 726.17 b ± 35.30 | 899.35 a ± 13.57 | - | <0.0001 |
| 1 CH4 (g CH4 animal−1 day−1) | 169 | 132.81 ± 0.07 | 132.93 ± 0.03 | 132.96 ± 0.04 | 0.1858 |
| Nutritive Metrics (% DM) | Defoliation Frequency | |
|---|---|---|
| r | p-Value | |
| NDF | 0.29 | <0.001 |
| ADF | 0.35 | <0.001 |
| ADL | −0.26 | 0.112 |
| Minerals | 0.19 | 0.060 |
| CP | −0.31 | <0.001 |
| DMD | −0.36 | 0.005 |
| OMD | −0.38 | <0.001 |
| ME (MJ kg−1 DM) | 0.38 | <0.001 |
| OM (g kg−1) | 0.14 | 0.334 |
| 1 CH4 (g CH4 animal−1 day−1) | 0.29 | <0.001 |
| Nutritive Metrics | Effect | Estimate | Se | DF | t Value | p-Value |
|---|---|---|---|---|---|---|
| CP | Intercept | 12.41 | 0.85 | 37 | 14.47 | <0.0001 |
| Defoliation frequency | −0.082 | 0.007 | 78 | −10.45 | <0.0001 | |
| Defoliation intensity | 0.083 | 0.087 | 78 | 0.96 | 0.3411 | |
| NDF | Intercept | 59.59 | 2.54 | 30 | 23.4 | <0.0001 |
| Defoliation frequency | 0.092 | 0.035 | 73 | 2.59 | 0.0117 | |
| Defoliation intensity | 0.031 | 0.308 | 73 | 0.1 | 0.9192 | |
| ADF | Intercept | 39.13 | 3.93 | 39 | 9.95 | <0.0001 |
| Defoliation frequency | 0.038 | 0.011 | 107 | 3.24 | 0.0016 | |
| Defoliation intensity | −0.524 | 0.316 | 107 | −1.66 | 0.1005 | |
| ME | Intercept | 6.29 | 0.43 | 40 | 14.55 | <0.0001 |
| Defoliation frequency | −0.013 | 0.004 | 87 | 2.72 | 0.0078 | |
| Defoliation intensity | 0.126 | 0.037 | 87 | 3.37 | 0.0011 | |
| OMD | Intercept | 50.59 | 2.70 | 40 | 18.71 | <0.0001 |
| Defoliation frequency | −0.083 | 0.030 | 87 | −2.72 | 0.0078 | |
| Defoliation intensity | 0.793 | 0.235 | 87 | 3.37 | 0.0011 | |
| DMD | Intercept | 47.51 | 4.86 | 12 | 9.78 | <0.0001 |
| Defoliation frequency | 0.028 | 0.082 | 31 | 0.34 | 0.7336 | |
| Defoliation intensity | 0.70 | 0.19 | 31 | 3.59 | 0.0011 | |
| 1 CH4 | Intercept | 133.18 | 0.12 | 42 | 1027.65 | <0.0001 |
| Defoliation frequency | 0.00031 | 0.00082 | 108 | 0.39 | 0.701 | |
| Defoliation intensity | −0.0254 | 0.0099 | 108 | −2.56 | 0.012 | |
| ADL | Intercept | 7.77 | 2.56 | 9 | 3.03 | 0.0142 |
| Defoliation frequency | 0.22 | 0.061 | 12 | 3.7 | 0.003 | |
| Defoliation intensity | −0.094 | 0.073 | 12 | −1.29 | 0.221 | |
| Minerals | Intercept | 14.78 | 3.19 | 19 | 4.63 | 0.0002 |
| Defoliation frequency | −0.051 | 0.0085 | 59 | −6.05 | <0.0001 | |
| Defoliation intensity | −0.44 | 0.20 | 59 | −2.2 | 0.0315 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jayasinghe, P.; Ramilan, T.; Donaghy, D.J.; Pembleton, K.G.; Barber, D.G. Comparison of Nutritive Values of Tropical Pasture Species Grown in Different Environments, and Implications for Livestock Methane Production: A Meta-Analysis. Animals 2022, 12, 1806. https://doi.org/10.3390/ani12141806
Jayasinghe P, Ramilan T, Donaghy DJ, Pembleton KG, Barber DG. Comparison of Nutritive Values of Tropical Pasture Species Grown in Different Environments, and Implications for Livestock Methane Production: A Meta-Analysis. Animals. 2022; 12(14):1806. https://doi.org/10.3390/ani12141806
Chicago/Turabian StyleJayasinghe, Priyanath, Thiagarajah Ramilan, Daniel J. Donaghy, Keith G. Pembleton, and David G. Barber. 2022. "Comparison of Nutritive Values of Tropical Pasture Species Grown in Different Environments, and Implications for Livestock Methane Production: A Meta-Analysis" Animals 12, no. 14: 1806. https://doi.org/10.3390/ani12141806
APA StyleJayasinghe, P., Ramilan, T., Donaghy, D. J., Pembleton, K. G., & Barber, D. G. (2022). Comparison of Nutritive Values of Tropical Pasture Species Grown in Different Environments, and Implications for Livestock Methane Production: A Meta-Analysis. Animals, 12(14), 1806. https://doi.org/10.3390/ani12141806