
Genetic Diversity and Genetic Differentiation of Populations of Golden-Backed Carp (Cyprinus carpio var. Jinbei) in Traditional Rice Fields in Guizhou, China

Abstract
:Simple Summary
Abstract
1. Introduction
2. Sample Collection and Methods
2.1. Materials
2.2. Extraction and Detection of Genomic DNA
2.3. Library Construction and Sequencing
2.4. Bioinformatics Analysis
3. Results
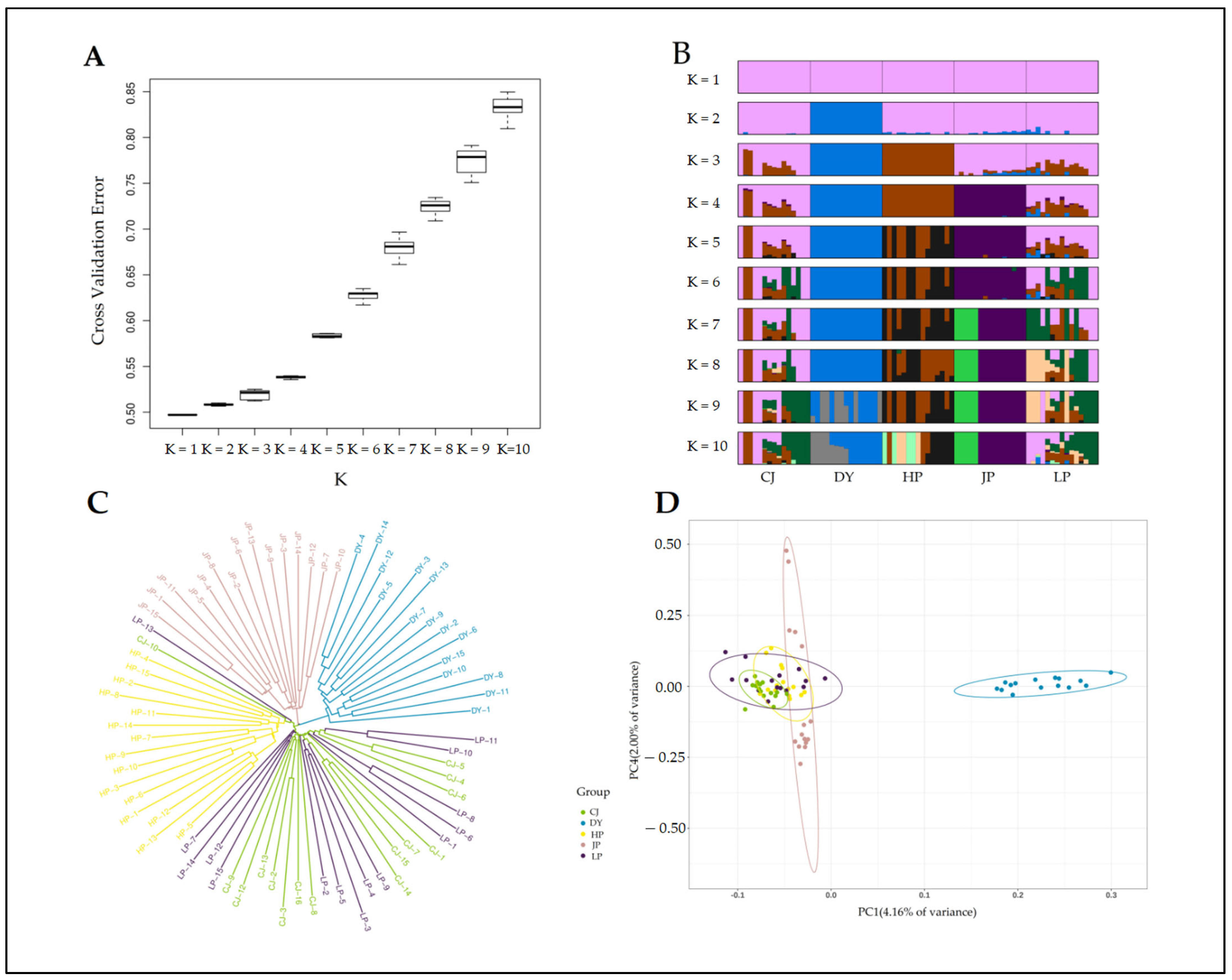
3.1. Genetic Diversity Parameters of Five Geographical Populations of Golden-Backed Carp in Guizhou, China
3.2. Genetic Differentiation of Five Populations of Golden-Backed Carp in Guizhou, China
4. Discussion
4.1. The Current Status and Reasons for the Low Genetic Diversity of Golden-Backed Carp in Five Geographical Populations in Guizhou, China
4.2. Status Quo of Genetic Differentiation among Populations of Golden-Backed Carp in Guizhou, China
4.3. Recommendations for the Conservation of Germplasm Resources of Golden-Backed Carp
- Purify the germplasm of the golden-backed carp and use pure-breeding parents for body colour for artificial reproduction.
- For the restoration of the germplasm of the golden-backed carp, the scope of breeding should be expanded as much as possible, inbreeding should be avoided, and good seeds should be regularly introduced from foreign breeding farms as breeding parents to improve the genetic diversity of offspring.
- Make full use of the globally important agricultural cultural heritage of the Congjiang Dongxiang Rice-fish-Duck Composite System area to carry out protection work.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.J.; Huai, Y.; Xu, J.F. Controlling effects and mechanisms on rice diseases, pests and weeds in rice-based coculture system. Zhejiang Agric. J. 2018, 6, 1016–1021. [Google Scholar]
- Li, J.Y.; Chang, D.; Li, B.N.; Wu, X.G.; Zhu, Z.W.; Cheng, Y.X. Benefit-cost analysis of different rice-based production systems. J. Fish. 2014, 9, 1431–1438. [Google Scholar]
- Shen, J.W.; Gu, Z.M.; Shen, H.G.; Hu, Y.Y.; Yang, Z.N.; Hu, S.R.; Liu, Y.H.; Zhang, X.Q. Soil Microorganism Species in Congjiang Dong’s Rice-Fish-Duck Complex System. Guizhou Agric. Sci. 2015, 2, 84–89. [Google Scholar]
- Zhou, X.; Xi, Y.W. A study on the restoration of rice field ecosystems by fish-rice co-cultivation. China Fish. 2016, 2, 76–79. [Google Scholar]
- Yang, H.L.; Feng, Z.M.; Lv, Y. Study on Eco-environmental Impact of Different Rice Modes. Anhui Agric. Sci. 2015, 17, 258–262, 293. [Google Scholar]
- Tang, S.K.; Liu, L.F.; Tang, L.; Wen, J. Comprehensive Economic Benefits of Rice-duck Biological Interaction. J. Agron. 2019, 10, 84–88. [Google Scholar]
- You, Z.R. Research on the Current Situation and Development Countermeasures of Rice and Fish Farming Industry in Yunnan Province. Ph.D. Thesis, Central China Normal University, Wuhan, China, 2015. [Google Scholar]
- Xu, P. Development and prospect of integrated rice-fish farming in China: A review. J. Dalian Ocean. Univ. 2021, 5, 717–726. [Google Scholar]
- Ye, M.L. A model of a pond rice field excavated in Guizhou. Guizhou Lit. Hist. Ser. 1990, 4, 32–37. [Google Scholar]
- Zhu, Y.X. Guizhou Congjiang makes good use of the world-class business card of agricultural heritage. World Herit. Site 2015, 10, 55–56. [Google Scholar]
- “13th Five-Year Plan” Report on the Development of China’s Rice and Fishery Comprehensive Breeding Industry. China Fish. 2022, 1, 43–52.
- Hammond, R.A.; Hudson, M.D. Environmental management of UK golf courses for biodiversity—Attitudes and actions. Landsc. Urban Plan. 2007, 83, 127–136. [Google Scholar] [CrossRef]
- Wu, J.; Liu, Z.M. Effect of habitat fragmentation on biodiversity. Chin. J. Ecol. 2014, 33, 1946–1952. [Google Scholar]
- Deng, C.Y. Comparative analysis of Cyt b and D-loop sequence of mitochondrial DNA from four populations of Carassius auratus. Mod. Agric. Sci. Technol. 2015, 9, 277–279. [Google Scholar]
- Wen, Y.F.; Han, W.J.; Wu, S. Plant genetic diversity and its influencing factors. J. Cent. South Univ. For. Technol. 2010, 30, 80–87. [Google Scholar]
- Wu, Z.J.; Li, Y.M. Effects of habitat fragmentation on survival of animal populations. Acta Ecol. Sin. 2003, 11, 2424–2435. [Google Scholar]
- Yu, W.H.; Wu, B.F.; Liu, Y.B. Effects of habitat fragmentation on genetic diversity of plants and animals. Chin. J. Appl. Environ. Biol. 2019, 25, 743–749. [Google Scholar]
- Ren, W.; Hu, L.; Guo, L.; Zhang, J.; Tang, L.; Zhang, E.; Zhang, J.; Luo, S.; Tang, J.; Chen, X. Preservation of the Genetic Diversity of a Local Common Carp in the Agricultural Heritage Rice-fish System. Proc. Natl. Acad. Sci. USA 2018, 115, E546–E554. [Google Scholar] [CrossRef] [Green Version]
- Gan, B.J.; Zhang, S.; Wei, L.J.; Huang, J.; Mo, J.L.; Gan, X.J.; Teng, Z.Z.; Ye, X.C. Genetic Structure Analysis of Local Variety of Common Carp Cyprinus carpio var. Jinbian Farmed in Paddy Fields in Guangxi Region. Fish. Sci. 2019, 38, 636–646. [Google Scholar]
- Ji, D.; Xu, J.S.; Yao, J.J.; An, Y.Y.; Zhao, W.; Yu, Z.K.; Zhang, C.B. Genetic Diversity and Genetic Structure Analysis of Five Geographical Populations of Jinbei Common Carp (Cyprinus carpio var. Jinbei) in Guizhou Province. Chin. J. Fish. 2021, 3, 1–10. [Google Scholar]
- Wang, S.; Liu, P.; Lv, J.; Li, Y.; Cheng, T.; Zhang, L.; Xia, Y.; Sun, H.; Hu, X.; Bao, Z. Serial Sequencing of Isolength Rad Tags for Cost-Efficient Genome-Wide Profiling of Genetic and Epigenetic Variations. Nat. Protoc. 2016, 11, 2189. [Google Scholar] [CrossRef]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Yu, C.; Li, Y.; Lam, T.W.; Yiu, S.M.; Kristiansen, K.; Wang, J. SOAP2: An improved ultrafast tool for short read alignment. Bioinformatics 2009, 25, 1966–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, X.; Dou, J.; Mao, J.; Su, H.; Jiao, W.; Zhang, L.; Hu, X.; Huang, X.; Wang, S.; Bao, Z. Radtyping: An Integrated Package for Accurate De Novo Codominant and Dominant Rad Genotyping in Mapping Populations. PLoS ONE 2013, 8, e79960. [Google Scholar] [CrossRef] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.Y.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danecek, P.; Auton, A.; Abecasis, G.; Cornelis, A.; Banks, A.E. The Variant Call Format and Vcftools. Bioinformatics 2011, 27, 2516–2518. [Google Scholar] [CrossRef] [PubMed]
- Vilella, A.J.; Severin, J.; Ureta-Vidal, A.; Heng, L.; Durbin, R.; Birney, E. EnsemblCompara GeneTrees: Complete, duplication-aware phylogenetic trees in vertebrates. Genome Res. 2009, 19, 327–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast Model-Based Estimation of Ancestry in Unrelated Individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Fornal, A.; Kowalska, K.; Zabek, T.; Piestrzynska-Kajtoch, A.; Musiał, A.D.; Ropka-Molik, K. Genetic Variability and Population Structure of Polish Konik Horse Maternal Lines Based on Microsatellite Markers. Genes 2021, 12, 546. [Google Scholar]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a Genetic Linkage Map in Man Using Restriction Fragment Length Polymorphisms. Am. J. Hum. Genet. 1980, 32, 314. [Google Scholar]
- Dong, Z.J.; Liu, N.; Fu, J.J.; Zhu, W.B.; Wang, L.M.; Su, S.Y. Genetic analysis for six wild and selection populations of common carp (Cyprinus carpio) using microsatellites. South China Fish. Sci. 2018, 14, 46–55. [Google Scholar]
- Takezaki, N.; Nei, M. Genetic distances and reconstruction of phylogenetic trees from microsatellite DNA. Genetics 1996, 144, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Hubert, S.; Hedgecock, D. Linkage maps of microsatellite DNA markers for the Pacific oyster Crassostrea gigas. Genetics 2004, 168, 351–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornuet, J.M.; Luikart, G. Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data. Genetics 1996, 144, 2001–2014. [Google Scholar] [CrossRef]
- Sinclair, E.A.; Hobbs, R.J. Sample Size Effects on Estimates of Population Genetic Structure: Implications for Ecological Restoration. Restor. Ecol. 2009, 17, 837–844. [Google Scholar] [CrossRef]
- Peter, B.M.; Slatkin, M. The effective founder effect in a spatially expanding population. Evolution 2015, 69, 721–734. [Google Scholar] [CrossRef] [Green Version]
- Nevo, E. Evolution of Genome–Phenome Diversity under Environmental Stress. Proc. Natl. Acad. Sci. USA 2001, 98, 6233–6240. [Google Scholar] [CrossRef] [Green Version]
- Quan, Y.C.; Sun, X.W.; Liang, L.Q. Genetic Polymorphism of Microsatellite DNA in Two Populations of Northern Sheatfish (Silurus soldatovi). Yi Chuan Xue Bao 2006, 33, 908–916. [Google Scholar] [CrossRef]
- Shaklee, J.B.; Tamaru, C.S.; Waples, R.S. Speciation and Evolution of Marine Fishes Studied by the Electrophoretic Analysis of Proteins. Pacif. Sci. 1982, 15, 419–432. [Google Scholar]
- Santos, S. Novel Microsatellite Markers Used for Determining Genetic Diversity and Tracing of Wild and Farmed Populations of the Amazonian Giant Fish Arapaima gigas. Genes 2021, 12, 1324. [Google Scholar]
- Qi, M.; Zhang, Y.Y.; Li, J.J.; Du, F. Genetic diversity and differentiation study of Quercus dentata with regards to its distribution in China and Japan. J. Lanzhou Univ. Nat. Sci. 2021, 57, 720–734. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Group | HW-P | He | Ho | PIC | Ne | Pi |
|---|---|---|---|---|---|---|
| CJ | 0.900 | 0.123 | 0.137 | 0.104 | 1.183 | 0.127 |
| DY | 0.883 | 0.115 | 0.118 | 0.096 | 1.177 | 0.119 |
| HP | 0.887 | 0.112 | 0.116 | 0.095 | 1.168 | 0.116 |
| JP | 0.882 | 0.117 | 0.123 | 0.099 | 1.179 | 0.122 |
| LP | 0.896 | 0.126 | 0.139 | 0.108 | 1.188 | 0.131 |
| Group | DY | HP | JP | LP | CJ |
|---|---|---|---|---|---|
| DY | 0.098 | 0.099 | 0.078 | 0.087 | |
| HP | 0.094 | 0.075 | 0.050 | 0.053 | |
| JP | 0.094 | 0.073 | 0.055 | 0.059 | |
| LP | 0.075 | 0.049 | 0.054 | 0.007 | |
| CJ | 0.084 | 0.052 | 0.057 | 0.007 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, D.; Su, X.; Yao, J.; Zhang, W.; Wang, R.; Zhang, S. Genetic Diversity and Genetic Differentiation of Populations of Golden-Backed Carp (Cyprinus carpio var. Jinbei) in Traditional Rice Fields in Guizhou, China. Animals 2022, 12, 1377. https://doi.org/10.3390/ani12111377
Ji D, Su X, Yao J, Zhang W, Wang R, Zhang S. Genetic Diversity and Genetic Differentiation of Populations of Golden-Backed Carp (Cyprinus carpio var. Jinbei) in Traditional Rice Fields in Guizhou, China. Animals. 2022; 12(11):1377. https://doi.org/10.3390/ani12111377
Chicago/Turabian StyleJi, Da, Xin Su, Junjie Yao, Wenzheng Zhang, Rongrong Wang, and Shuhai Zhang. 2022. "Genetic Diversity and Genetic Differentiation of Populations of Golden-Backed Carp (Cyprinus carpio var. Jinbei) in Traditional Rice Fields in Guizhou, China" Animals 12, no. 11: 1377. https://doi.org/10.3390/ani12111377
APA StyleJi, D., Su, X., Yao, J., Zhang, W., Wang, R., & Zhang, S. (2022). Genetic Diversity and Genetic Differentiation of Populations of Golden-Backed Carp (Cyprinus carpio var. Jinbei) in Traditional Rice Fields in Guizhou, China. Animals, 12(11), 1377. https://doi.org/10.3390/ani12111377