
The Cryopreserved Sperm Traits of Various Ram Breeds: Towards Biodiversity Conservation

 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Animal Management
2.2. Semen Collection and Processing
2.3. Diluent Preparation and Freezing Process
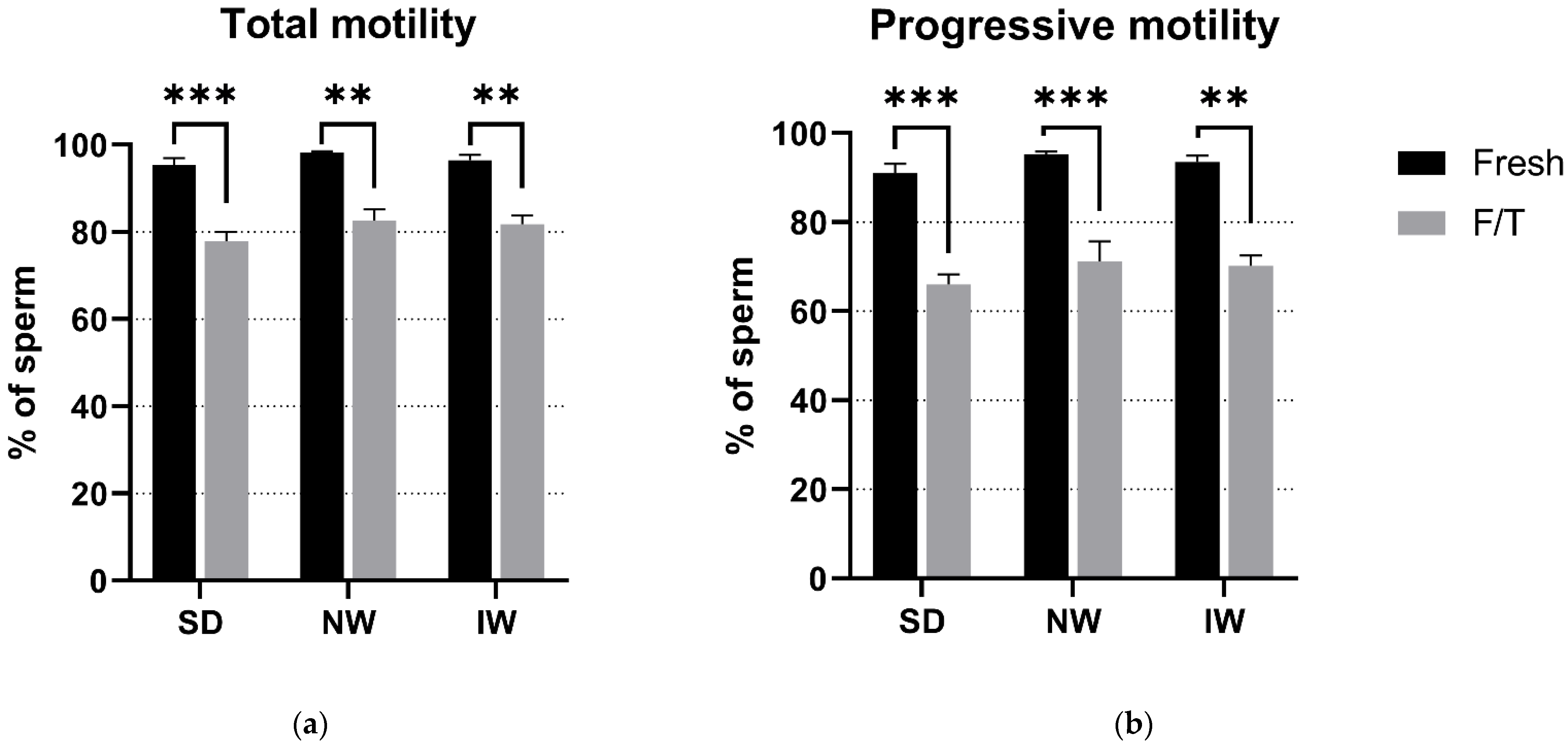
2.4. Sperm Motility Evaluation
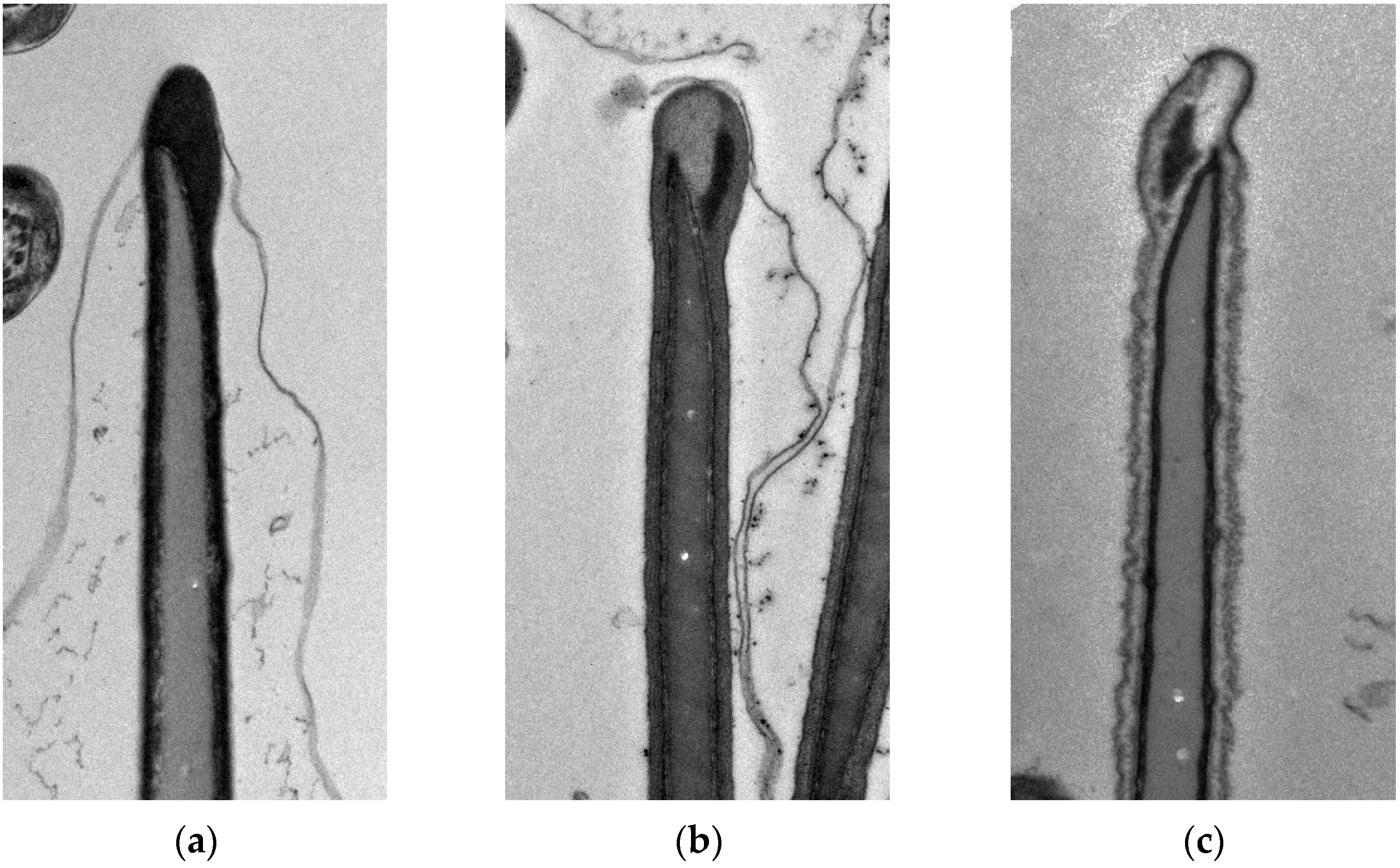
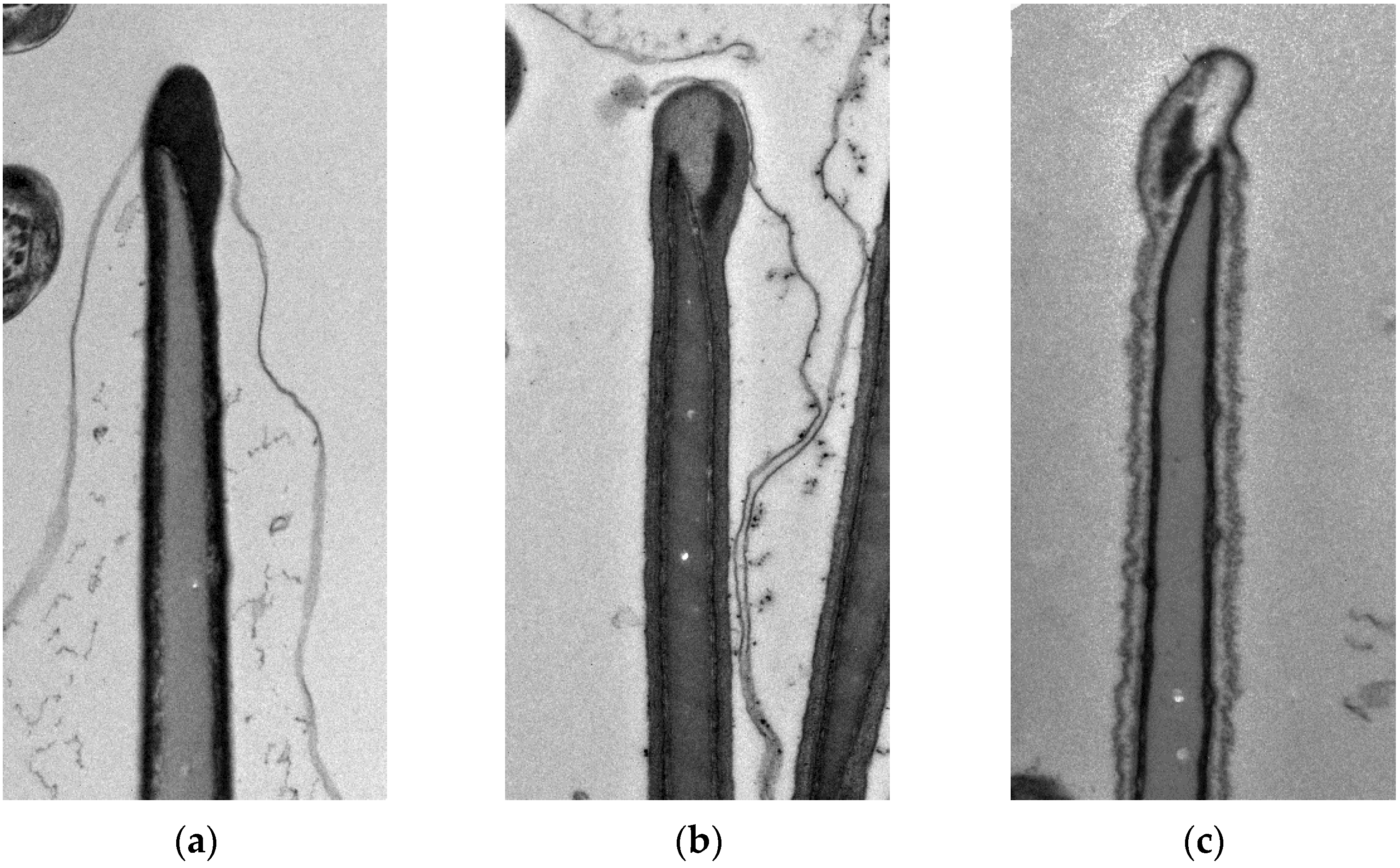
2.5. Morphological Changes
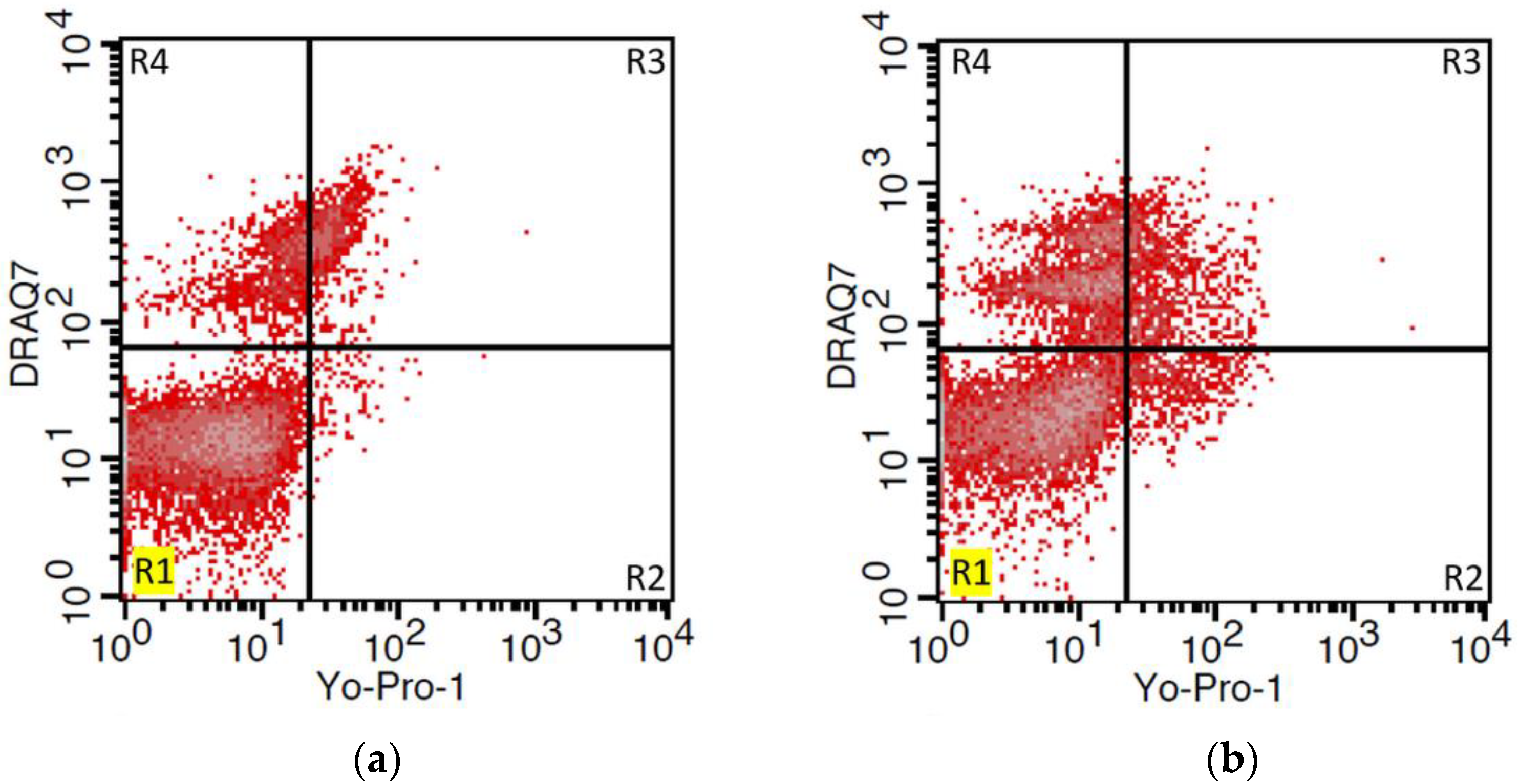
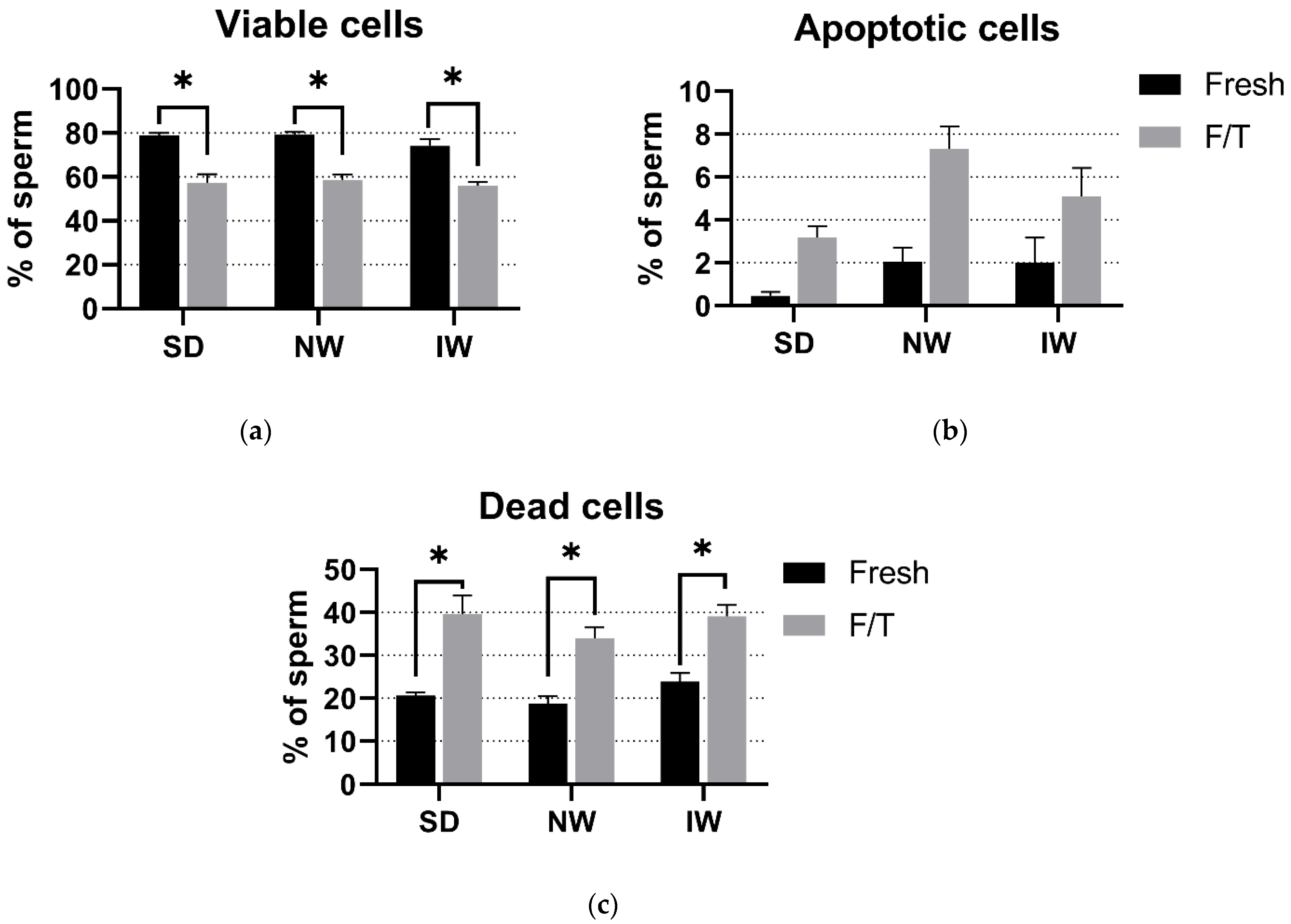
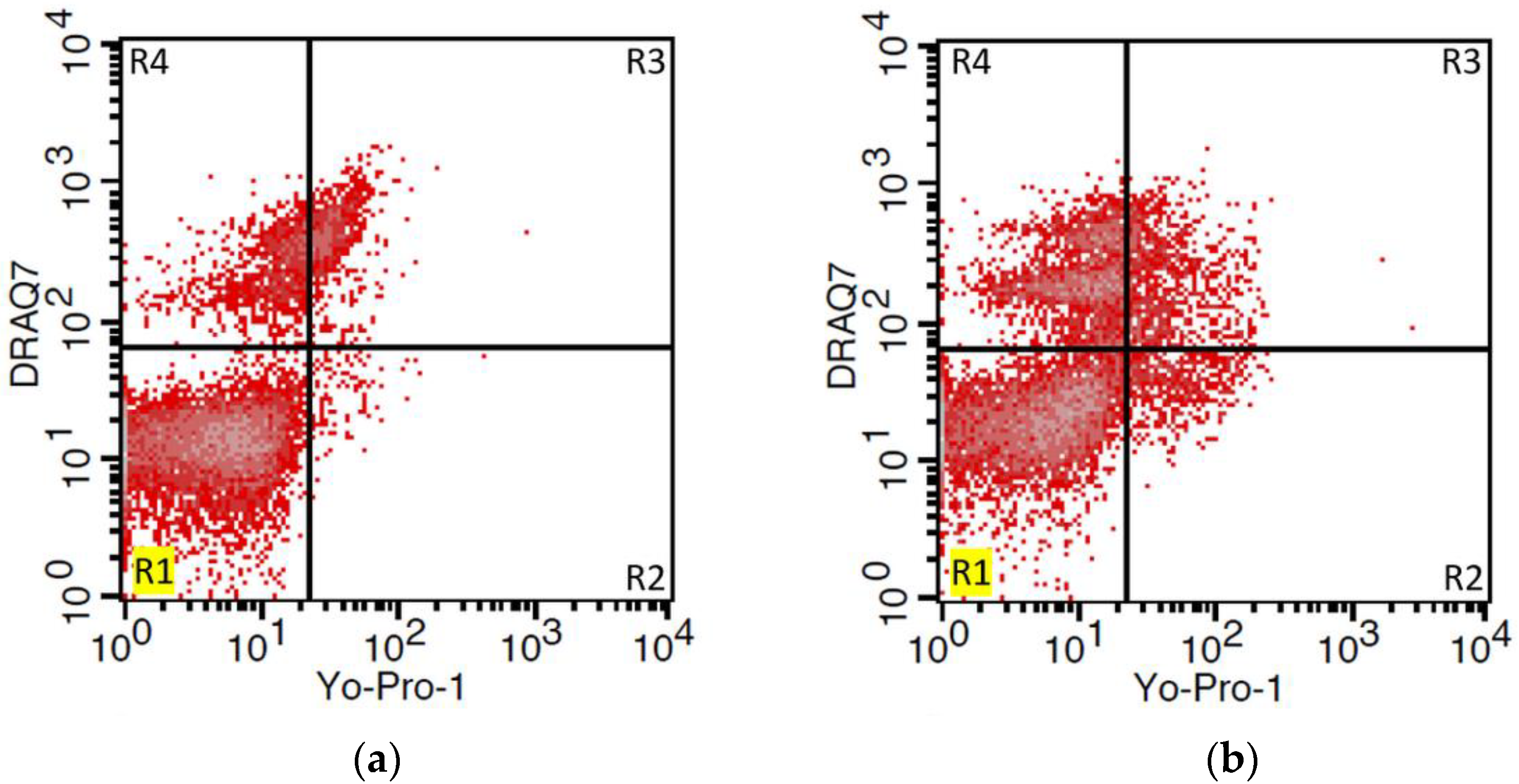
2.6. Viability and Apoptosis Assay
2.7. Test of Sperm Penetrating/Fertilizing Ability
2.8. Acrosomal Status
2.9. Statistical Analysis
3. Results
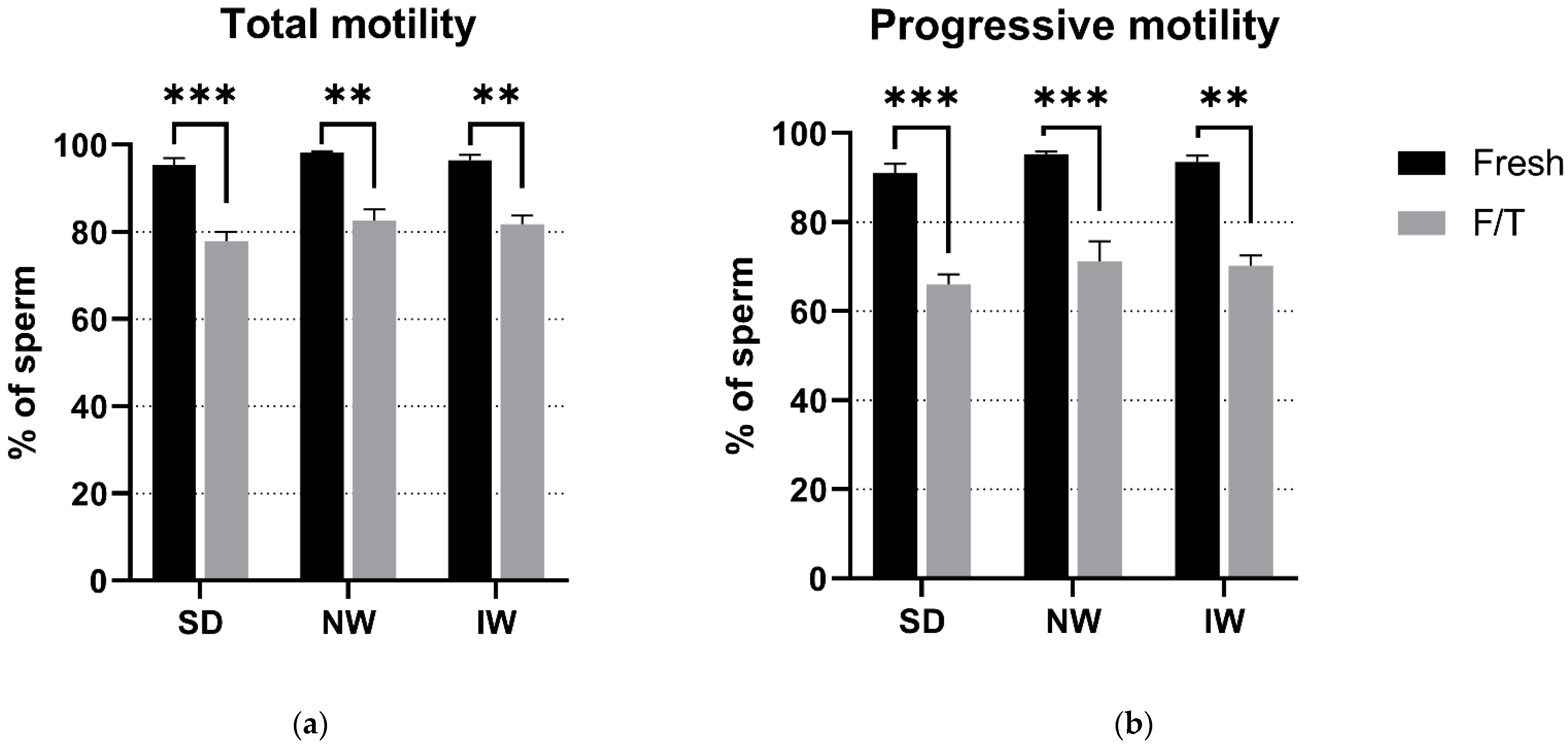
3.1. Sperm Motility Evaluation
3.2. Morphological Changes
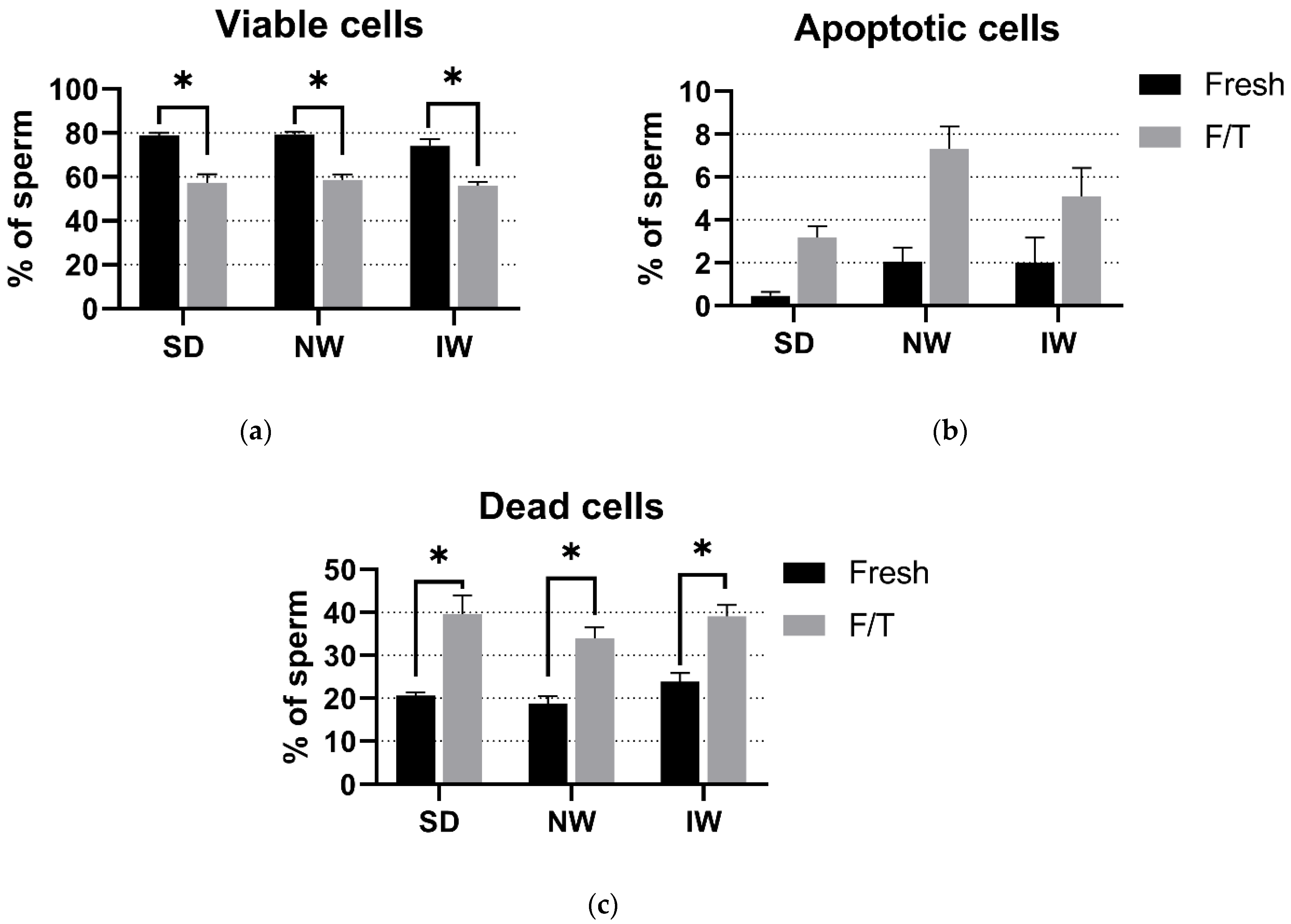
3.3. Viability and Apoptosis Assay
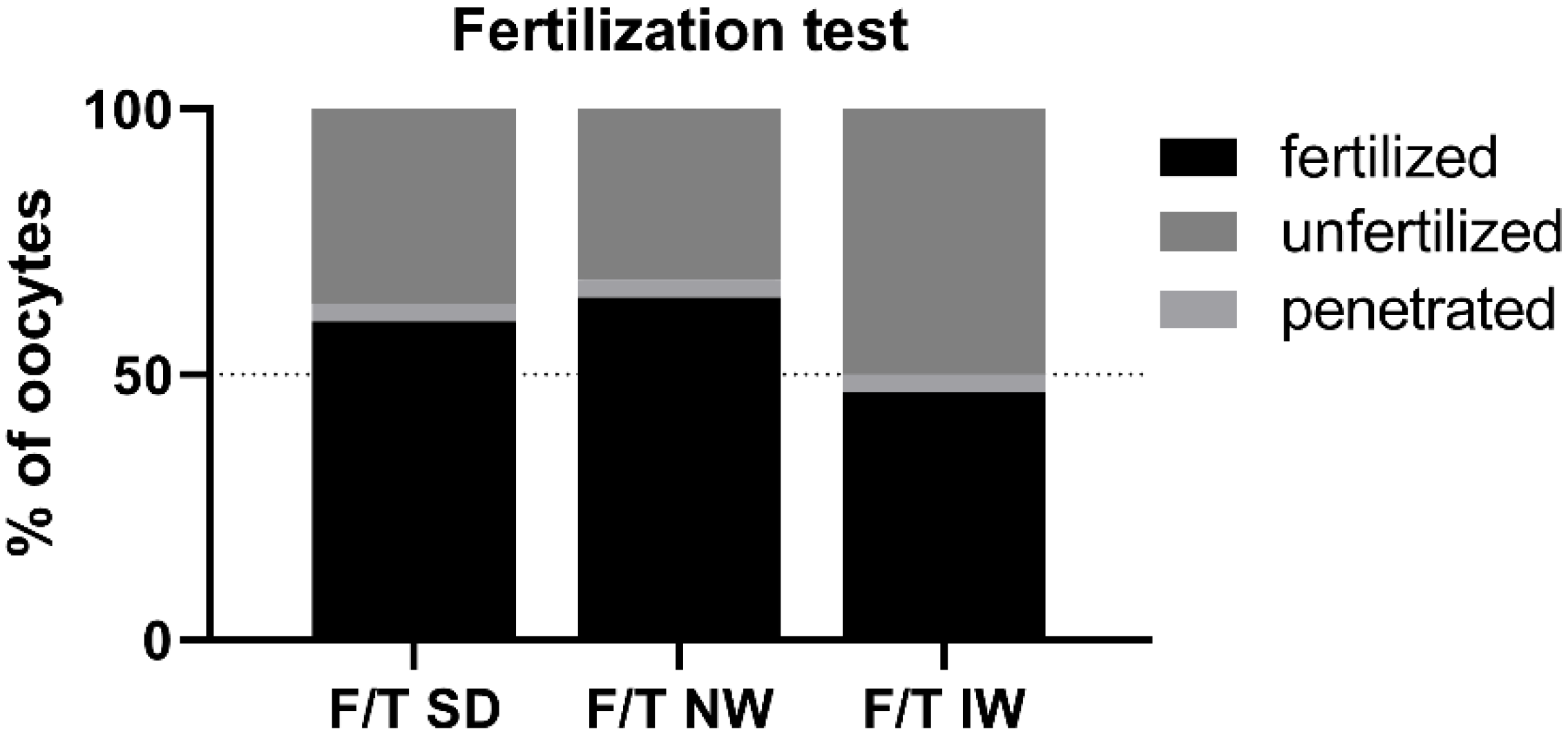
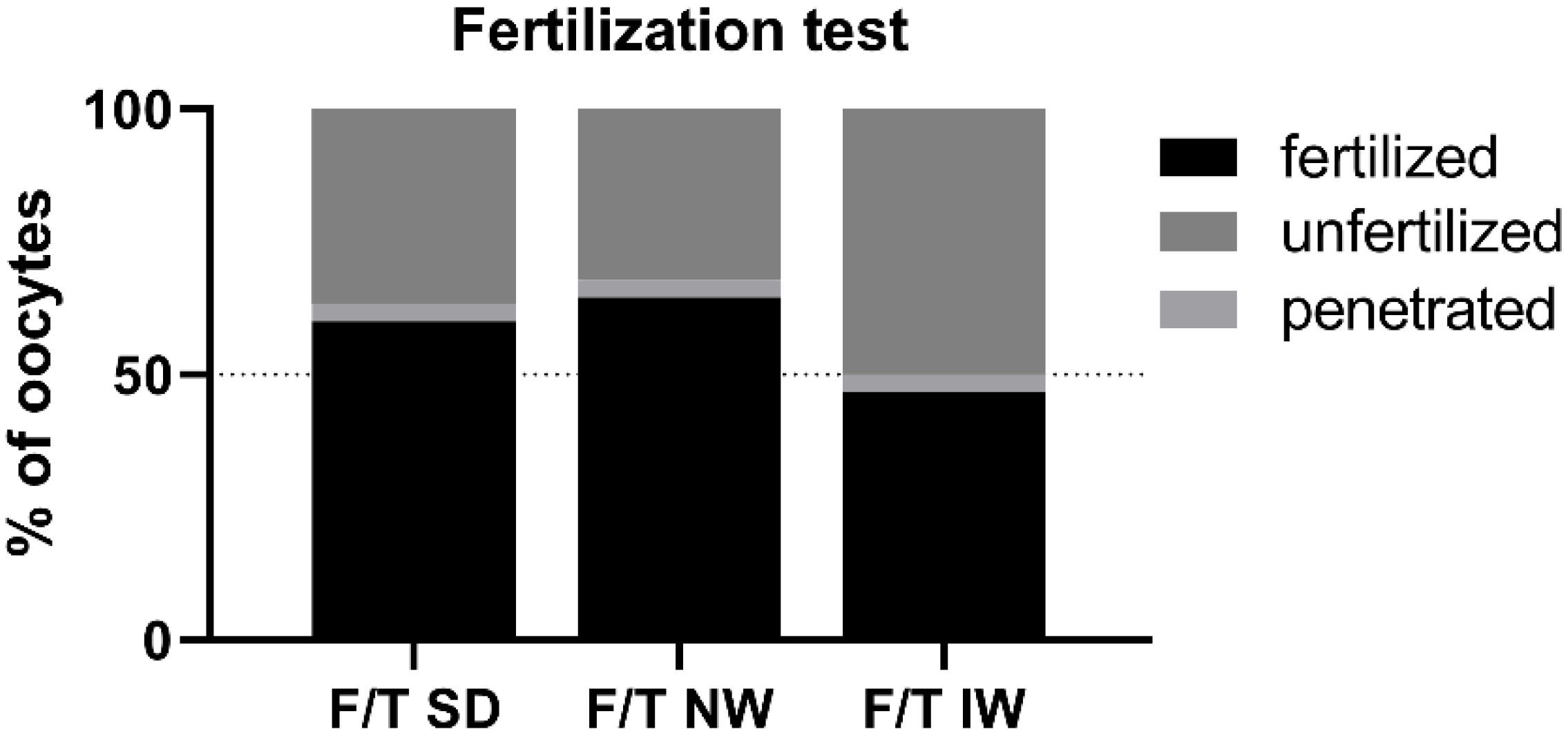
3.4. Test of Sperm Penetrating/Fertilizing Ability
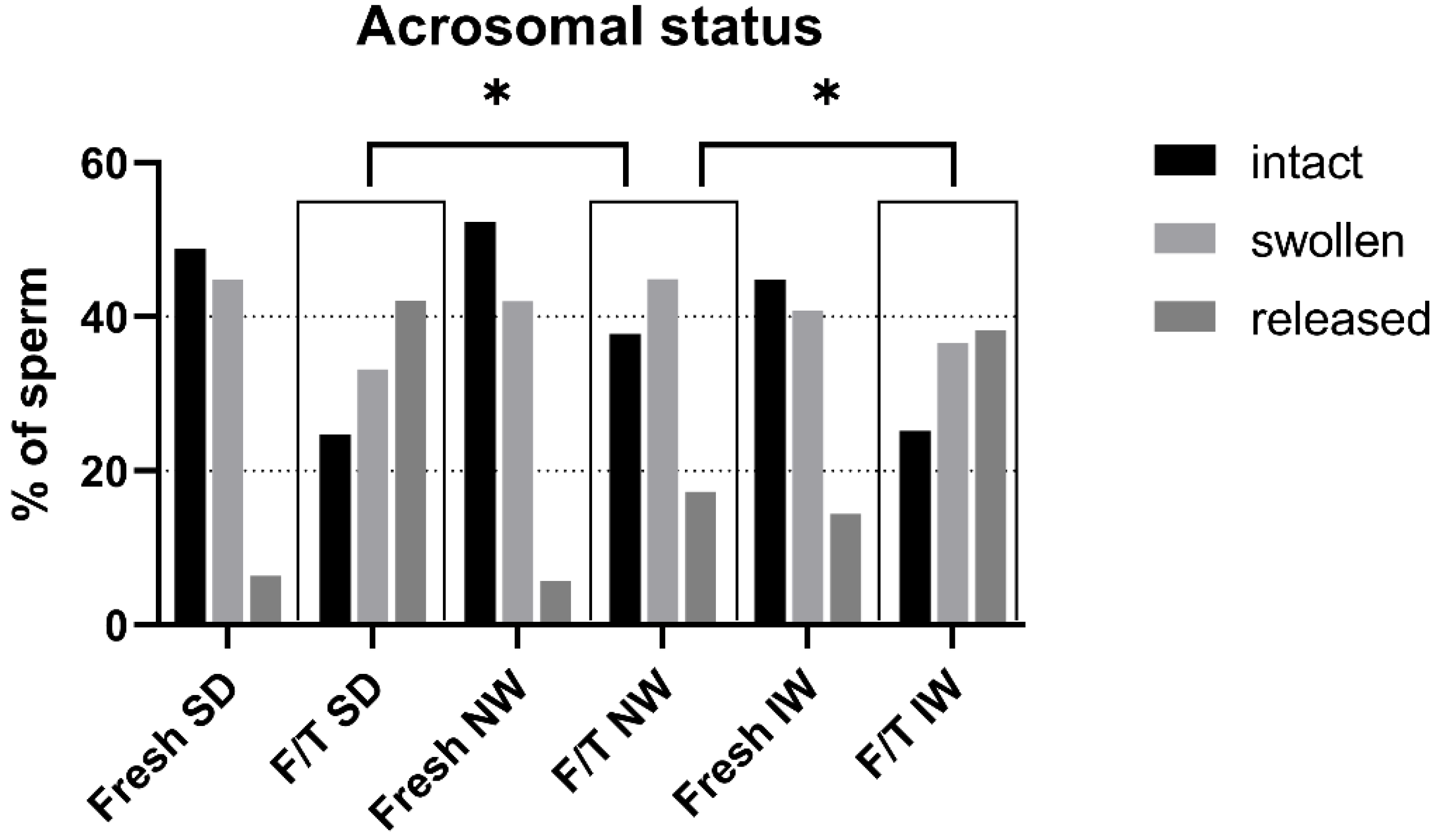
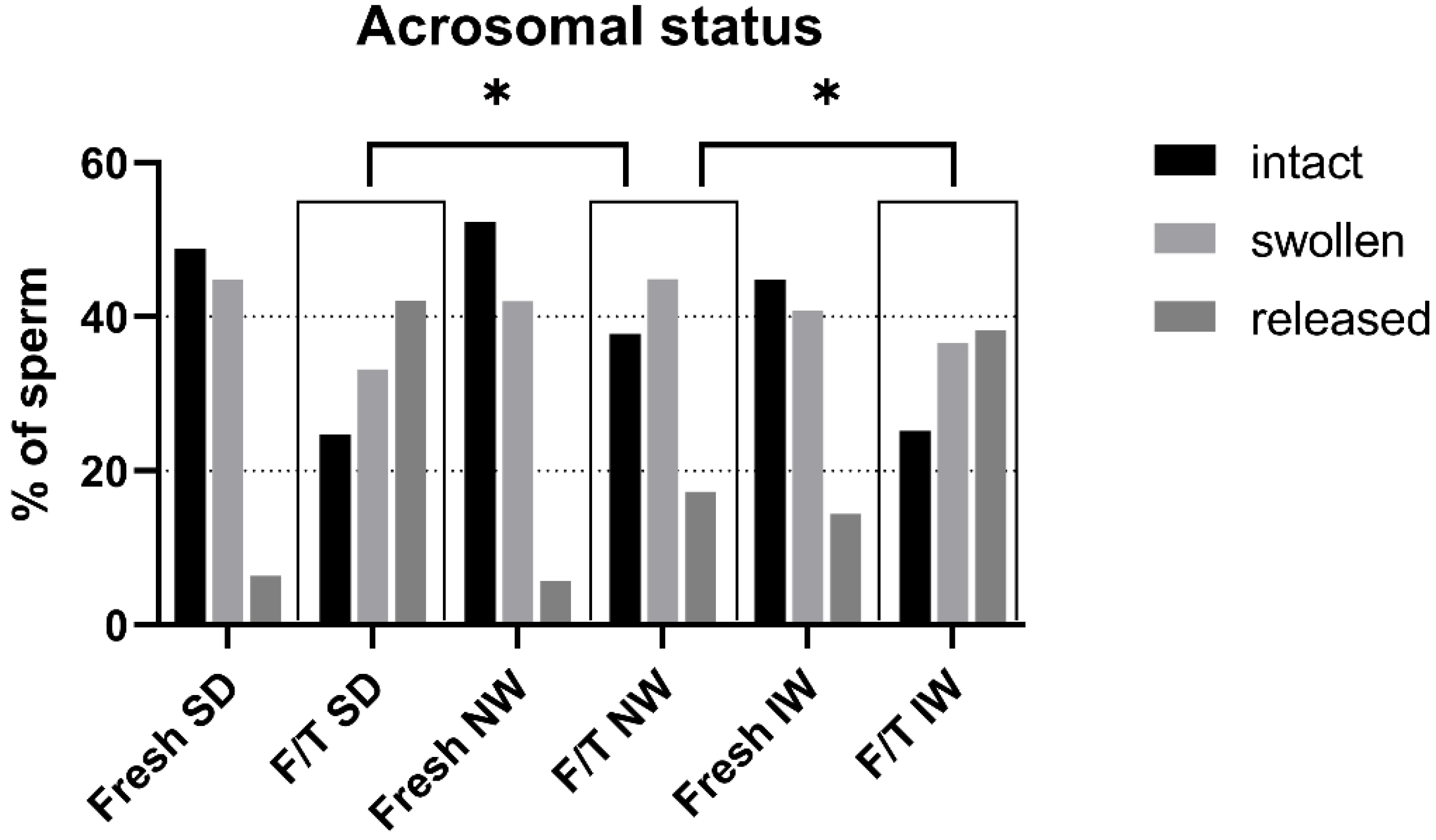
3.5. Acrosomal Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Silva, S.; Soares, A.; Batista, A.; Almeida, F.; Nunes, J.; Peixoto, C.; Guerra, M. In Vitro and In Vivo Evaluation of Ram Sperm Frozen in Tris Egg-Yolk and Supplemented with Superoxide Dismutase and Reduced Glutathione. Reprod. Domest. Anim. 2011, 46, 874–881. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Rabadán, P.; Soler, A.J.; Ramón, M.; García-Álvarez, O.; Maroto-Morales, A.; Iniesta-Cuerda, M.; Fernández-Santos, M.R.; Montoro, V.; Pérez-Guzmán, M.D.; Garde, J.J. Influence of Semen Collection Method on Sperm Cryoresistance in Small Ruminants. Anim. Reprod. Sci. 2016, 167, 103–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raspa, M.; Guan, M.; Paoletti, R.; Montoliu, L.; Ayadi, A.; Marschall, S.; Fray, M.; Scavizzi, F. Dry Ice Is a Reliable Substrate for the Distribution of Frozen Mouse Spermatozoa: A Multi-Centric Study. Theriogenology 2017, 96, 49–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gürler, H.; Malama, E.; Heppelmann, M.; Calisici, O.; Leiding, C.; Kastelic, J.P.; Bollwein, H. Effects of Cryopreservation on Sperm Viability, Synthesis of Reactive Oxygen Species, and DNA Damage of Bovine Sperm. Theriogenology 2016, 86, 562–571. [Google Scholar] [CrossRef] [PubMed]
- Küçük, N.; Aksoy, M.; Uçan, U.; Ahmad, E.; Naseer, Z.; Ceylan, A.; Serin, İ. Comparison of Two Different Cryopreservation Protocols for Freezing Goat Semen. Cryobiology 2014, 68, 327–331. [Google Scholar] [CrossRef]
- Bailey, J.L.; Blodeau, J.-F.; Cormier, N. Semen Cryopreservation in Domestic Animals: A Damaging and Capacitating Phenomenon Minireview. J. Androl. 2000, 21, 1–7. [Google Scholar] [CrossRef]
- Alcay, S.; Berk Toker, M.; Gokce, E.; Ustuner, B.; Tekin Onder, N.; Sagirkaya, H.; Nur, Z.; Kemal Soylu, M. Successful Ram Semen Cryopreservation with Lyophilized Egg Yolk-Based Extender. Cryobiology 2015, 71, 329–333. [Google Scholar] [CrossRef]
- Amirat, L.; Tainturier, D.; Jeanneau, L.; Thorin, C.; Gérard, O.; Courtens, J.L.; Anton, M. Bull Semen in Vitro Fertility after Cryopreservation Using Egg Yolk LDL: A Comparison with Optidyl®, a Commercial Egg Yolk Extender. Theriogenology 2004, 61, 895–907. [Google Scholar] [CrossRef]
- Arando, A.; Gonzalez, A.; Delgado, J.V.; Arrebola, F.A.; Perez-Marín, C.C. Storage Temperature and Sucrose Concentrations Affect Ram Sperm Quality after Vitrification. Anim. Reprod. Sci. 2017, 181, 175–185. [Google Scholar] [CrossRef]
- Tibary, A.; Manar, S. Cryo-Preservation of Sperm and Embryos in Small Ruminants. Rev. Maroc. Sci. Agron. Vétérinaires 2018, 6, 195–210. [Google Scholar]
- Luna-Orozco, J.R.; González-Ramos, M.A.; Calderón-Leyva, G.; Gaytán-Alemán, L.R.; Arellano-Rodríguez, F.; Ángel-García, O.; Véliz-Deras, F.G. Comparison of Different Diluents Based on Liposomes and Egg Yolk for Ram Semen Cooling and Cryopreservation. Iran. J. Vet. Res. 2019, 20, 126–130. [Google Scholar] [PubMed]
- Bohlool, Z.; Mohammadi, M.; Mehr, M.R.A.; Hossein-Zadeh, N.G. Effect of Different Concentrations of Trehalose and Glycerol on the Freezability of Ram Semen Using Soybean Lecithin-Based Diluents. Anim. Prod. Sci. 2015, 55, 666–671. [Google Scholar] [CrossRef]
- Emamverdi, M.; Zhandi, M.; Zare Shahneh, A.; Sharafi, M.; Akbari‐Sharif, A. Optimization of Ram Semen Cryopreservation Using a Chemically Defined Soybean Lecithin‐Based Extender. Reprod. Domest. Anim. 2013, 48, 899–904. [Google Scholar] [CrossRef] [PubMed]
- Forouzanfar, M.; Sharafi, M.; Hosseini, S.M.; Ostadhosseini, S.; Hajian, M.; Hosseini, L.; Abedi, P.; Nili, N.; Rahmani, H.R.; Nasr-Esfahani, M.H. In Vitro Comparison of Egg Yolk–Based and Soybean Lecithin–Based Extenders for Cryopreservation of Ram Semen. Theriogenology 2010, 73, 480–487. [Google Scholar] [CrossRef]
- Masoudi, R.; Sharafi, M.; Zareh Shahneh, A.; Towhidi, A.; Kohram, H.; Zhandi, M.; Esmaeili, V.; Shahverdi, A. Effect of Dietary Fish Oil Supplementation on Ram Semen Freeze Ability and Fertility Using Soybean Lecithin– and Egg Yolk–Based Extenders. Theriogenology 2016, 86, 1583–1588. [Google Scholar] [CrossRef] [Green Version]
- Najafi, A.; Najafi, M.H.; Zanganeh, Z.; Sharafi, M.; Martinez-Pastor, F.; Adeldust, H. Cryopreservation of Ram Semen in Extenders Containing Soybean Lecithin as Cryoprotectant and Hyaluronic Acid as Antioxidant. Reprod. Domest. Anim. 2014, 49, 934–940. [Google Scholar] [CrossRef]
- Sharafi, M.; Zhandi, M.; Sharif, A.A. Supplementation of Soybean Lecithin-Based Semen Extender by Antioxidants: Complementary Flowcytometric Study on Post-Thawed Ram Spermatozoa. Cell Tissue Bank. 2014, 16, 261–269. [Google Scholar] [CrossRef]
- Toker, M.B.; Alcay, S.; Gokce, E.; Ustuner, B. Cryopreservation of Ram Semen with Antioxidant Supplemented Soybean Lecithin-Based Extenders and Impacts on Incubation Resilience. Cryobiology 2016, 72, 205–209. [Google Scholar] [CrossRef]
- Üstüner, B.; Alçay, S.; Nur, Z.; Saǧirkaya, H.; Kemal Soylu, M. Effect of Egg Yolk and Soybean Lecithin on Tris-Based Extender in Post-Thaw Ram Semen Quality and in Vitro Fertility. Kafkas Univ. Vet. Fak. Derg. 2014, 20, 393–398. [Google Scholar] [CrossRef]
- Vozaf, J.; Makarevich, A.V.; Balazi, A.; Vasicek, J.; Svoradova, A.; Olexikova, L.; Chrenek, P. Cryopreservation of Ram Semen: Manual versus Programmable Freezing and Different Lengths of Equilibration. Anim. Sci. J. 2021, 92, e13670. [Google Scholar] [CrossRef]
- Chrenek, P.; Makarevič, A.V.; Kubovičová, E.; Bulla, J.; Supuka, P. Slovak National Animal Breeds; Slovak University of Agriculture: Nitra, Slovakia, 2019; ISBN 978-80-552-1982-0. [Google Scholar]
- García-Álvarez, O.; Maroto-Morales, A.; Martínez-Pastor, F.; Fernández-Santos, M.R.; Esteso, M.C.; Pérez-Guzmán, M.D.; Soler, A.J. Heterologous in Vitro Fertilization Is a Good Procedure to Assess the Fertility of Thawed Ram Spermatozoa. Theriogenology 2009, 71, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Olexikova, L.; Miranda, M.; Kulikova, B.; Baláži, A.; Chrenek, P. Cryodamage of Plasma Membrane and Acrosome Region in Chicken Sperm. Anat. Histol. Embryol. 2019, 48, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joost, S.; Bruford, M.W. The Genomic-Resources Consortium Editorial: Advances in Farm Animal Genomic Resources. Front. Genet. 2015, 6, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mara, L.; Casu, S.; Carta, A.; Dattena, M. Cryobanking of Farm Animal Gametes and Embryos as a Means of Conserving Livestock Genetics. Anim. Reprod. Sci. 2013, 138, 25–38. [Google Scholar] [CrossRef] [PubMed]
- FAO. In Vivo Conservation of Animal Genetic Resources 2013. FAO Animal Production and Health Guidelines. No. 14. Rome. Available online: https://portals.iucn.org/library/sites/library/files/documents/Bios-Cons-Gen-040.pdf (accessed on 9 February 2022).
- Rakha, B.A.; Ansari, M.S.; Akhter, S.; Hussain, I.; Blesbois, E. Cryopreservation of Indian Red Jungle Fowl (Gallus gallus murghi) Semen. Anim. Reprod. Sci. 2016, 174, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Svoradová, A.; Kuželová, L.; Vašíček, J.; Olexíková, L.; Chrenek, P. Cryopreservation of Chicken Blastodermal Cells and Their Quality Assessment by Flow Cytometry and Transmission Electron Microscopy. Biotechnol. Prog. 2018, 34, 778–783. [Google Scholar] [CrossRef]
- Curry, M.R.; Kleinhans, F.W.; Watson, P.F. Measurement of the Water Permeability of the Membranes of Boar, Ram, and Rabbit Spermatozoa Using Concentration-Dependent Self-Quenching of an Entrapped Fluorophore. Cryobiology 2000, 41, 167–173. [Google Scholar] [CrossRef]
- Gáspárdy, A.; Renkó, E.; Somoskői, B.; Bába, A.; Cseh, S. Practical Experience with Artificial Insemination (AI) Using Fresh Chilled and Frozen Semen in Mares. Acta Vet. Hung. 2020, 68, 85–90. [Google Scholar] [CrossRef]
- Barbas, J.P.; Mascarenhas, R.D. Cryopreservation of Domestic Animal Sperm Cells. Cell Tissue Bank. 2009, 10, 49–62. [Google Scholar] [CrossRef]
- Kumaresan, A.; Elango, K.; Datta, T.K.; Morrell, J.M. Cellular and Molecular Insights Into the Etiology of Subfertility/Infertility in Crossbred Bulls (Bos taurus × Bos indicus): A Review. Front. Cell Dev. Biol. 2021, 9, 696637. [Google Scholar] [CrossRef]
- Amann, R.P.; Waberski, D. Computer-Assisted Sperm Analysis (CASA): Capabilities and Potential Developments. Theriogenology 2014, 81, 5–17.e3. [Google Scholar] [CrossRef] [PubMed]
- Galarza, D.A.; López-Sebastián, A.; Woelders, H.; Blesbois, E.; Santiago-Moreno, J. Two-Step Accelerating Freezing Protocol Yields a Better Motility, Membranes and DNA Integrities of Thawed Ram Sperm than Three-Steps Freezing Protocols. Cryobiology 2019, 91, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Peris-Frau, P.; Martín-Maestro, A.; Iniesta-Cuerda, M.; Sánchez-Ajofrín, I.; Mateos-Hernández, L.; Garde, J.J.; Villar, M.; Soler, A.J. Freezing–Thawing Procedures Remodel the Proteome of Ram Sperm before and after In Vitro Capacitation. Int. J. Mol. Sci. 2019, 20, 4596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masoudi, R.; Sharafi, M.; Zareh Shahneh, A.; Towhidi, A.; Kohram, H.; Esmaeili, V.; Shahverdi, A.; Davachi, N.D. Fertility and Flow Cytometry Study of Frozen-Thawed Sperm in Cryopreservation Medium Supplemented with Soybean Lecithin. Cryobiology 2016, 73, 69–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khawar, M.B.; Gao, H.; Li, W. Mechanism of Acrosome Biogenesis in Mammals. Front. Cell Dev. Biol. 2019, 7, 195. [Google Scholar] [CrossRef] [PubMed]
- Meyers, S.A. Chapter 5—Sperm Physiology. In Equine Breeding Management and Artificial Insemination, 2nd ed.; Samper, J.C., Ed.; W.B. Saunders: Saint Louis, MO, USA, 2009; pp. 47–55. ISBN 978-1-4160-5234-0. [Google Scholar]
- Yanagimachi, R. Mammalian Sperm Acrosome Reaction: Where Does It Begin before Fertilization? Biol. Reprod. 2011, 85, 4–5. [Google Scholar] [CrossRef] [Green Version]
- Bernecic, N.C.; Donnellan, E.; O’Callaghan, E.; Kupisiewicz, K.; O’Meara, C.; Weldon, K.; Lonergan, P.; Kenny, D.A.; Fair, S. Comprehensive Functional Analysis Reveals That Acrosome Integrity and Viability Are Key Variables Distinguishing Artificial Insemination Bulls of Varying Fertility. J. Dairy Sci. 2021, 104, 11226–11241. [Google Scholar] [CrossRef]
- Parks, J.E.; Lynch, D.V. Lipid Composition and Thermotropic Phase Behavior of Boar, Bull, Stallion, and Rooster Sperm Membranes. Cryobiology 1992, 29, 255–266. [Google Scholar] [CrossRef]
- De Las Mercedes Carro, M.; Peñalva, D.A.; Antollini, S.S.; Hozbor, F.A.; Buschiazzo, J. Cholesterol and Desmosterol Incorporation into Ram Sperm Membrane before Cryopreservation: Effects on Membrane Biophysical Properties and Sperm Quality. Biochim. Biophys. Acta BBA—Biomembr. 2020, 1862, 183357. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abnormality | SD | NW | IW |
|---|---|---|---|
| F-ST | 2.0 ± 0.07 | 3.0 ± 0.2 | 2.0 ± 0.08 |
| F-KT | 3.8 ± 0.2 | 2.0 ± 0.16 | 3.0 ± 0.2 |
| F-TT | 1.0 ± 0.04 | - | 1.0 ± 0.04 |
| F-RT | 2.0 ± 0.06 | 3.0 ± 0.34 | 2.0 ± 0.1 |
| F-BT | - | 1.0 ± 0.1 | - |
| F-CD | - | - | 0.8 ± 0.04 |
| F-Total | 8.8 ± 0.4 | 9.0 ± 0.6 | 8.8 ± 0.5 |
| F/T-ST | 4.0 ± 0.17 | 5.0 ± 0.4 | 3.0 ± 0.07 |
| F/T-KT | 2.0 ± 0.08 | 2.0 ± 0.14 | 1.0 ± 0.05 |
| F/T-TT | 0.5 ± 0.05 | 1.0 ± 0.05 | 2.0 ± 0.1 |
| F/T-RT | 3.0 ± 0.2 | 3.3 ± 0.1 | 4.0 ± 0.2 |
| F/T-BT | 2.0 ± 0.15 | 2.0 ± 0.07 | 1.3 ± 0.1 |
| F/T-CD | 1.0 ± 0.03 | 1.0 ± 0.05 | 1.0 ± 0.05 |
| F/T-Total | 12.5 ± 1.0 | 14.3 ± 1.2 | 12.3 ± 0.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vozaf, J.; Svoradová, A.; Baláži, A.; Vašíček, J.; Olexiková, L.; Dujíčková, L.; Makarevich, A.V.; Jurčík, R.; Ďúranová, H.; Chrenek, P. The Cryopreserved Sperm Traits of Various Ram Breeds: Towards Biodiversity Conservation. Animals 2022, 12, 1311. https://doi.org/10.3390/ani12101311
Vozaf J, Svoradová A, Baláži A, Vašíček J, Olexiková L, Dujíčková L, Makarevich AV, Jurčík R, Ďúranová H, Chrenek P. The Cryopreserved Sperm Traits of Various Ram Breeds: Towards Biodiversity Conservation. Animals. 2022; 12(10):1311. https://doi.org/10.3390/ani12101311
Chicago/Turabian StyleVozaf, Jakub, Andrea Svoradová, Andrej Baláži, Jaromír Vašíček, Lucia Olexiková, Linda Dujíčková, Alexander V. Makarevich, Rastislav Jurčík, Hana Ďúranová, and Peter Chrenek. 2022. "The Cryopreserved Sperm Traits of Various Ram Breeds: Towards Biodiversity Conservation" Animals 12, no. 10: 1311. https://doi.org/10.3390/ani12101311
APA StyleVozaf, J., Svoradová, A., Baláži, A., Vašíček, J., Olexiková, L., Dujíčková, L., Makarevich, A. V., Jurčík, R., Ďúranová, H., & Chrenek, P. (2022). The Cryopreserved Sperm Traits of Various Ram Breeds: Towards Biodiversity Conservation. Animals, 12(10), 1311. https://doi.org/10.3390/ani12101311