
Establishment of a Wisent (Bison bonasus) Germplasm Bank


 and
and 
Simple Summary
Abstract
1. Introduction
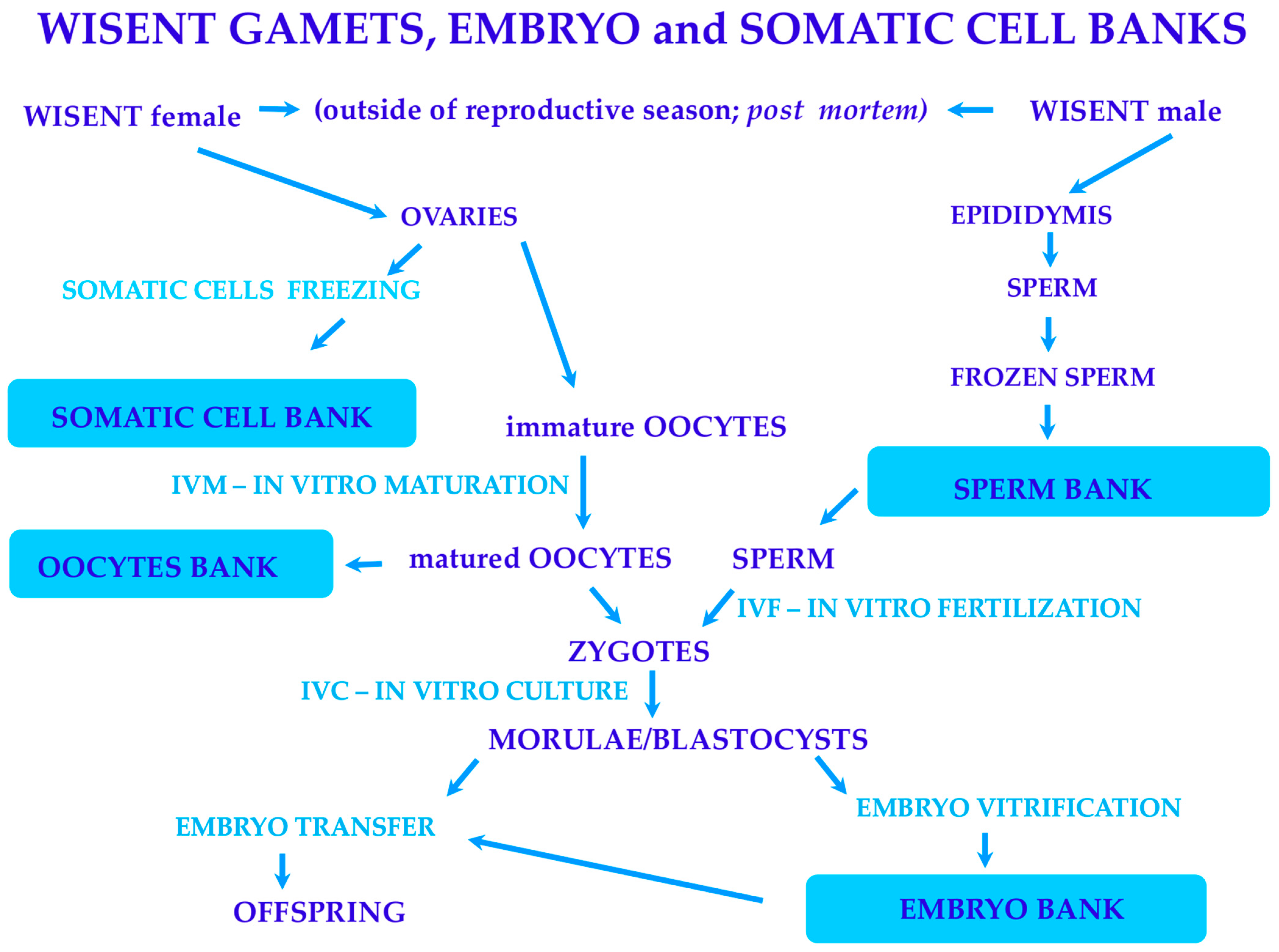
2. Materials and Methods
2.1. Ovaries Isolation
2.2. In Vitro Maturation of Oocytes
2.3. In Vitro Fertilization of Matured Oocytes
2.4. In Vitro Culture of Embryos
2.5. Vitrification
2.6. Interspecies Embryo Transfer
2.7. Monitoring Pregnancy
2.8. Statistical Analysis
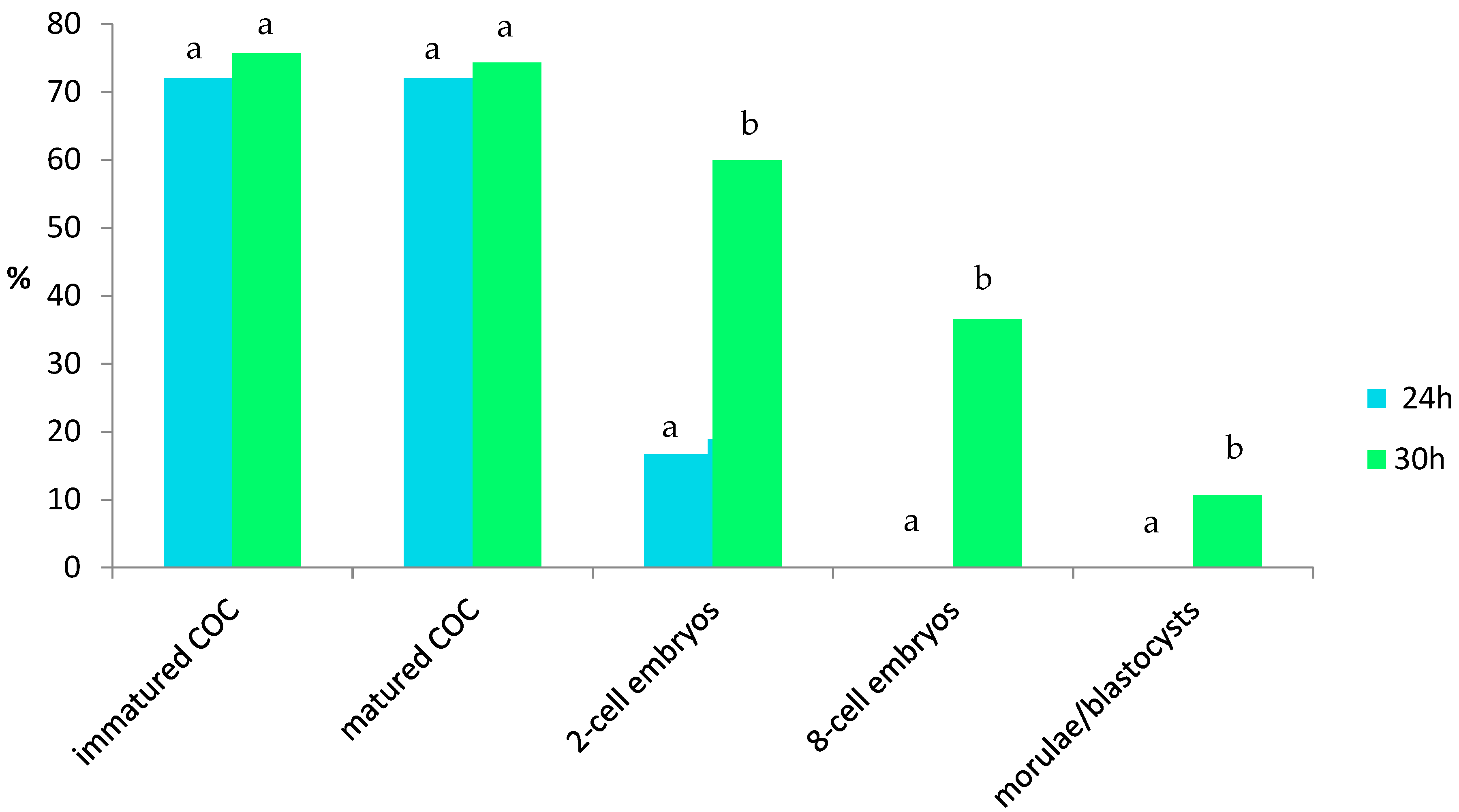
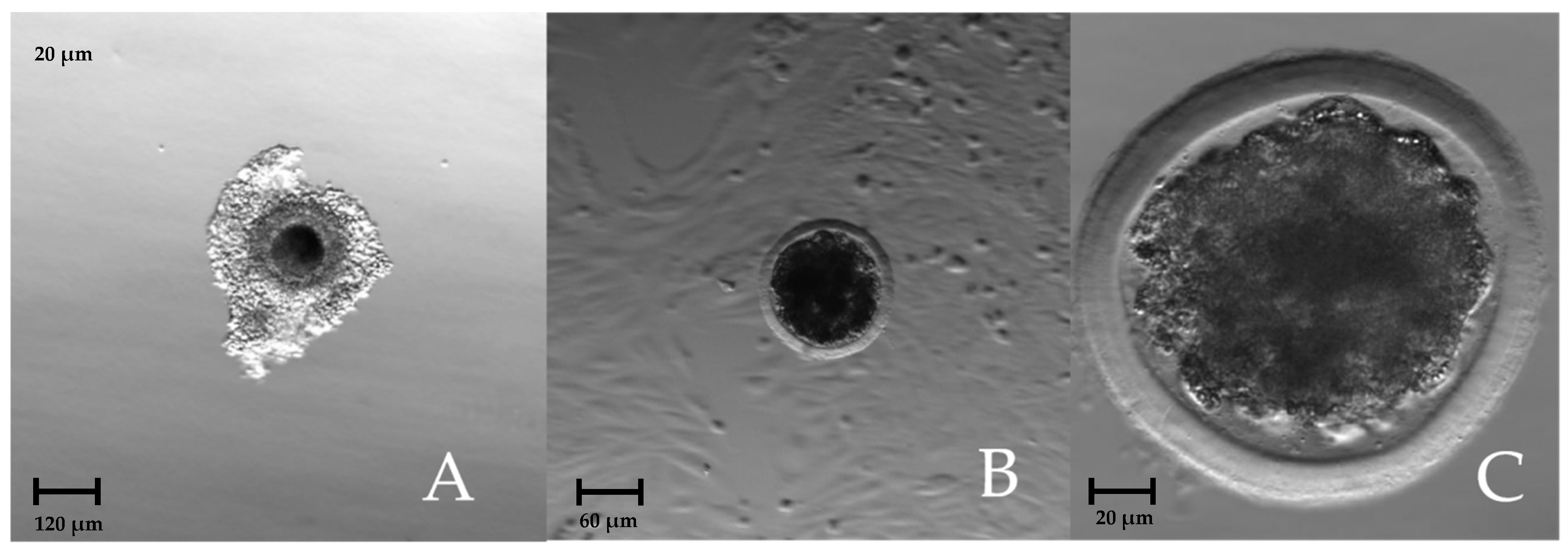
3. Results
4. Discussion
4.1. In Vitro Maturation of Wisent Oocytes
4.2. In Vitro Fertilization of Matured Wisent Oocytes
4.3. In Vitro Culture of Wisent Embryos
4.4. Vitrification of Wisent Embryos
4.5. Wisent Embryo Transfer to Cattle Recipients
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klich, D.; Olech, W.; Łopucki, R.; Danik, K. Community attitudes to the European bison Bison bonasus in areas where its reintroduction is planned and in areas with existing populations in northeastern Poland. Eur. J. Wildl. Res. 2018, 64, 61. [Google Scholar] [CrossRef]
- Pucek, Z.; Pucek, Z.; Belousova, I.P.; Krasińska, M.; Krasiński, Z.A.; Olech, W. European bison. Status Survey and Conservation Action Plan; IUCN Species Survival Commission (SSC): Gland, Switzerland; Cambridge, UK, 2004; 54p. [Google Scholar]
- Schwerk, A.; Klich, D.; Wójtowicz, E.; Olech, W. Impact of European bison Grazing (Bison bonasus (L.)) on Species and Functional Traits of Carabid Beetle Assemblages in Selected Habitats in Poland. Biology 2021, 10, 123. [Google Scholar] [CrossRef]
- Raczyński, J.; Bołbot, M. European bison Pedigree Book; Białowieża Natlional Park: Białowieża, Poland, 2020; pp. 1–83. [Google Scholar]
- Olech, W. The Genetic Variability within Bison bonasus species 90 years after bottleneck. In Restoration of Endangered and Extinct Animals; Słomski, R., Ed.; Poznan University of Life Sciences: Poznan, Poland, 2010; pp. 48–57. [Google Scholar]
- Demiaszkiewicz, A.W.; Moskwa, B.; Gralak, A.; Laskowski, Z.; Myczka, A.W.; Kołodziej-Sobocińska, M.; Kaczor, S.; Plis-Kuprianowicz, E.; Krzysiak, M.; Filip-Hutsch, K. The Nematodes Thelazia gulosa Railiet and Henry, 1910 and Thelazia skrjabini Erschov, 1928 as a Cause of Blindness in European Bison (Bison bonasus) in Poland. Acta Parasitol. 2020, 65, 963–968. [Google Scholar] [CrossRef]
- Joly, D.O.; Messier, F. The effect of bovine tuberculosis and brucellosis on reproduction and survival of wood bison in Wood Buffalo National Park. J. Anim. Ecol. 2005, 74, 543–551. [Google Scholar] [CrossRef]
- Kita, J.; Anusz, K.; Salwa, A.; Welz, M.; Orłowska, B.; Zaleska, M. Bovine Tuberculosis in European bison as Possible Zoonotic Impact in Poland. Zoonosis 2012, 7, 101–110. [Google Scholar]
- Krajewska-Wędzina, M.; Olech, W.; Kozińska, M.; Augustynowicz-Kopeć, E.; Weiner, M.; Szulowski, K. Bovine tuberculosis ourbreak in farmed American bison (Bison bison) in Poland. Pol. J. Vet. Sci. 2017, 20, 819–821. [Google Scholar]
- Plumb, G.; Kowalczyk, R.; Hernandez-Blanco, J.A. Bison bonasus. IUCN Red List. Threat. Species 2020, 1–19. [Google Scholar] [CrossRef]
- Saragusty, J.; Diecke, S.; Drukker, M.; Durrant, B.; Friedrich Ben-Nun, I.; Galli, C.; Göritz, F.; Hayashi, K.; Hermes, R.; Holtze, S.; et al. Rewinding the Process of Mammalian Extinction. Zoo Biol. 2016, 35, 280–292. [Google Scholar] [CrossRef]
- Olech, W.; Perzanowski, K. Supplementation of genetic structure through introductions of selected animals ex situ into free ranging wisent population of Bieszczady Mountains. Eur. Bison Conserv. Newsl. 2016, 9, 23–30. [Google Scholar]
- Olech, W.; Perzanowski, K. Changes of size and structure of the world population of European bison in years 2000–2015. Eur. Bison Conserv. Newsl. 2016, 9, 5–10. [Google Scholar]
- Olech, W.; Klich, D.; Perzanowski, K. Development of a new Action Plan for the European bison. Oryx 2019, 53, 214. [Google Scholar] [CrossRef]
- Choudhary, K.K.; Kavya, K.M.; Jerome, A.; Sharma, R.K. Advances in reproductive biotechnologies. Vet. World 2016, 9, 388–395. [Google Scholar] [CrossRef]
- Duszewska, A.M.; Baraniewicz-Kołek, M.; Bielecki, W.; Niżański, W.; Partyka, A.; Bernat, A.; Bihorac, A.; Olech, W. Cellular Technologies (In Vitro Production of Embryos, Germplasm Preservation and Somatic Reprogramming) for Wisent (European bison) Gene Pool Conservation. Concepts Dairy Vet. Sci. 2020, 338–340. [Google Scholar]
- Sanches, B.V.; Zangirolamo, A.F.; da Silva, N.C.; Morotti, F.; Seneda, M.M. Cryopreservation of in vitro-produced embryos: Challenges for commercial implementation. Anim. Reprod. 2017, 14, 521–527. [Google Scholar] [CrossRef]
- Silva, A.R.; Lima, G.L.; Peixoto, G.C.X.; Souza, A.L.P. Cryopreservation in mammalian conservation biology: Current applications and potential utility. Res. Rep. Biodivers. Stud. 2015, 4, 1–8. [Google Scholar] [CrossRef]
- De Souza, J.M.G.; Batista, R.I.T.P.; Melo, L.M.; Freitas, V.J.F. Reproductive biotechnologies applied to the conservation of endangered ruminant—Past, present and future. Rev. Port. Cienc. Vet. 2011, 106, 31–38. [Google Scholar]
- Bhat, G.R.; Sofi, K.A. Oocyte and embryo preservation in wild animals: An update. Cryoletters 2021, 42, 251–260. [Google Scholar]
- Marsico, T.V.; de Camargo, J.; Valente, R.S.; Sudano, M.J. Embryo competence and cryosurvival: Molecular and cellular features. Anim. Reprod. 2019, 16, 423–439. [Google Scholar] [CrossRef]
- Rienzi, L.; Gracia, C.; Maggiulli, R.; LaBarbera, A.R.; Kaser, D.J.; Ubaldi, F.M.; Vanderpoel, S.; Racowsky, C. Oocyte, embryo and blastocyst cryopreservation in ART: Systematic review and meta-analysis comparing slow-freezing versus vitrification to produce evidence for the development of global guidance. Hum. Reprod. Update 2017, 23, 139–155. [Google Scholar] [CrossRef]
- Benham, H.M.; McCollum, M.P.; Nol, P.; Frey, R.K.; Clarke, P.R.; Rhyan, J.C.; Barfield, J.P. Production of embryos and a live offspring using post mortem reproductive material from bison (Bison bison bison) originating in Yellowstone National Park, USA. Theriogenology 2021, 160, 33–39. [Google Scholar] [CrossRef]
- Barfield, J.P. In vitro production of bison embryos. Methods Mol Biol. 2019, 2006, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, M.P.; Palomino, J.M.; Anzar, M.; Mapletoft, R.J.; Adams, G.P. In vivo and in vitro maturation of oocytes collected from superstimulated wood bison (Bison bison athabascae) during the anovulatory and ovulatory seasons. Anim. Reprod. Sci. 2016, 73, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, M.P.; Palomino, J.M.; Anzar, M.; Mapletoft, R.J.; Mastromonaco, G.F.; Adams, G.P. In vitro embryo production in wood bison (Bison bison athabascae) using in vivo matured cumulus-oocyte complexes. Theriogenology 2017, 89, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Krishnakumar, S.; Whiteside, D.P.; Elkin, B.; Thundathil, J.C. Effect of reproductive seasonality on gamete quality in the North American bison (Bison bison bison). Reprod. Domest. Anim. 2015, 50, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Thundathil, J.; Whiteside, D.; Shea, B.; Ludbrook, D.; Elkin, B.; Nishi, J. Preliminary assessment of reproductive technologies in wood bison (Bison bison atha- bascae): Implications for preserving genetic diversity. Theriogenology 2007, 68, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Duszewska, A.M.; Rąpała, Ł.; Trzeciak, P.; Dąbrowski, S.; Piliszek, A. Obtaining farm animal embryos in vitro. J. Anim. Feed. Sci. 2012, 21, 217–233. [Google Scholar] [CrossRef]
- Duszewska, A.M.; Trzeciak, P.; Compa, A.; Rąpała, Ł. Selected issues concerning biotechnology of farm animals breeding—A review. Anim. Sci. Rep. 2010, 28, 295–306. [Google Scholar]
- Ferré, L.B.; Kjelland, M.E.; Strøbech, L.B.; Hyttel, P.; Mermillod, P.; Ross, P.J. Review: Recent advances in bovine in vitro embryo production: Reproductive biotechnology history and methods. Animal 2020, 14, 991–1004. [Google Scholar] [CrossRef]
- Eberhardt, M.; Niżanski, W.; Olech, W.; Prochowska, S. Assisted reproductive techniques in wisents: Achievements and further challenges. Med. Weter. Vet. Med. Sci. Pract. 2021, 77, 279–283. [Google Scholar] [CrossRef]
- Kozdrowski, R.; Niżański, W.; Dubiel, A.; Olech, W. Possibilities of using the European bison (Bison bonasus) epididymal spermatozoa collected post-mortem for cryopreservation and artificial insemination: A pilot study. Reprod. Biol. Endocrinol. 2011, 9, 31. [Google Scholar] [CrossRef]
- Seaby, R.P.; Alexander, B.; King, W.A.; Mastromonaco, G.F. In Vitro Development of Bison Embryos Using Interspecies Somatic Cell Nuclear Transfer. Reprod. Domest. Anim. 2013, 48, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Singina, G.N.; Baginov, W.A.; Dancz, S.S.; Taradaynik, T.E.; Docev, A.W.; Zinovieva, N.A. In vitro production of interspecies hybrid embryos of cattle (Bos taurus) and wisent (Bison bonasus). Agric. Biol. 2015, 51, 824–829. [Google Scholar] [CrossRef]
- Sipko, T.P.; Abilov, A.I.; Shurkhal, A.V.; Rott, N.N. Creation of bison sperm cryobank as a means of conserving genetic polymorphisms in the species. Russ. J. Genet. 1993, 29, 1516–1520. [Google Scholar]
- Sipko, T.P.; Rautian, G.S.; Udina, I.G.; Strelchenko, N.S. Conservation of genetic material from endangered and economically important species in the establishment of cryobanks. In Physiology and General Biology Reviews; Turpaev, T.M., Koltzov, N.K., Eds.; Russian Academy of Sciences: Moscow, Russia, 1997; pp. 35–98. [Google Scholar]
- Toosi, B.M.; Tribulo, A.; Lessard, C.; Mastromonaco, G.F.; McCorkell, R.B.; Adams, G.P. Superovulation and embryo transfer in wood bison (Bison bison athabascae). Theriogenology 2013, 80, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Duszewska, A.M.; Olech, W.; Trzeciak, P.; Krzysiak, M.; Rapala, L.; Nowak, Z.; Dabrowski, S. The preliminary results of in vitro production of the wisent hybrid embryos. Reprod. Fertil. Dev. 2014, 27, 219. [Google Scholar] [CrossRef]
- Duszewska, A.M.; Gręda, P.; Baraniewicz, M.; Bielecki, W.; Niżański, W.; Partyka, A.; Tracz, M.; Nowak, Z.; Chełmońska-Soyta, A.; Olech, W. Obtaining wisent early blastocyst in vitro is a basic for protection and creation of biodiversity for this threatened species. Reprod. Domest. Anim. 2018, 53, 818–821. [Google Scholar] [CrossRef]
- Riedl, J.; Wenigerkind, H.; Weppert, M.; Wolf, E. Prospects and obstacles of in-vitro-production and interspecies transfer of European bison (Bison bonasus) embryos. In Proceedings of the International Conference “Żubry w dolinie Sanu”, Muczne, Poland, 5–6 September 2018; p. 72. [Google Scholar]
- International Embryo Transfer Societes IETS. Certification and identification of embryos. In Manual of the International Embryo Transfer Society, 4th ed.; Stringfellow, D.A., Givens, M.D., Eds.; International Embryo Transfer Society: Champaign, IL, USA, 2010; pp. 86–105. [Google Scholar]
- Sekizawa, F.; Sawai, T.; Tanaka, M.; Okuda, K. Relationship between embryo collection results after superovulation treatment of Japanese Black cows and their plasma β-carotene and vitamin concentrations. J. Reprod. Dev. 2012, 58, 377–379. [Google Scholar] [CrossRef]
- Duszewska, A.; Koster, S.; Baraniewicz-Kolek, M.; Bielecki, W.; Nizanski, W.; Partyka, A.; Olech, W. The competence of wisent (Bison bonasus) sperm for in vitro fertilization. In Proceedings of the 23rd Annual Conference of the European-Society-for-Domestic-Animal-Reproduction (ESDAR), St. Petersburg, Russia, 19–22 September 2019; Volume 54, p. 125. [Google Scholar]
- Widayati, D.T. Embryo Transfer as an Assisted Reproductive Technology in Farm Animals. World Acad. Sci. Eng. Technol. 2012, 6, 70. [Google Scholar]
- Sousa, N.M.; Ayad, A.; Beckers, J.F.; Gajewski, Z. Pregnancy-associated Glycoproteins (PAG) as Pregnancy Markers in the Ruminants. J. Physiol. Pharmacol. 2006, 57, 153–171. [Google Scholar]
- Nöthling, J.O.; Cramer, K.G.M. Comparison of progesterone assay by chemiluminescence or radioimmunoassay for clinicaldecision-making in canine reproduction. J. S. Afr. Vet. Assoc. 2019, 22, e1–e6. [Google Scholar] [CrossRef]
- Tachi, S.; Tachi, C. Ultrastructural studies on maternal-embryonic cell interaction during experimentally induced implantation of rat blastocysts to the endometrium of the mouse. Dev. Biol. 1979, 68, 203–223. [Google Scholar] [CrossRef]
- Randi, R.; Fernandez-Fuertes, B.; McDonald, M.; Forde, N.; Kelly, A.K.; Amorin, H.B.; Muniz de Lima, E.; Morotti, F.; Marcondes Seneda, M.M.; Lonergan, P. Asynchronous embryo transfer as a tool to understand embryo–uterine interaction in cattle: Is a large conceptus a good thing? Reprod. Fertil. Dev. 2015, 28, 1999–2006. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Parameters | 24 h IVM | 30 h IVM |
|---|---|---|
| Number of donors | 4 | 6 |
| Number of COCs isolated/per donor | 12.50 ± 0.95 | 12.33 ± 0.5 |
| Immature COCs’ quality/per donor: | ||
| Compacted | 6.75 ± 1.25 | 7.2 ± 1.32 |
| Expanded | 1 ± 0.82 | 0.7 ± 0.81 |
| Denuded | 1.25 ± 0.95 | 1.5 ± 0.54 |
| Degenerated | 3.5 ± 0.57 | 3 ± 0.89 |
| Qualified oocytes for IVM/per donor | 9 ± 0.4 (72%) | 9.33 ± 0.61 (75.68%) |
| Matured COCs’ quality/per donor: | ||
| Compacted | 5.75 ± 0.95 a | 1 ± 1.21 b |
| Expanded | 2 ± 1.15 a | 6.7 ± 1.21 b |
| Denuded | 1.25 ± 0.95 | 1.5 ± 0.54 |
| Degenerated | 0 | 0.16 ± 0.41 |
| Qualified COsC for IVF 1 | 9 ± 0.4 (72%) | 9.16 ± 0.48 (74.32%) |
| Embryo development/per donor: | ||
| 2-cell | 1.25 ± 0.25 (18.88%) a | 5.5 ± 0.34 (59.96%) b |
| 8-cell | 0 a | 3.33 ± 0.21 (36.52%) b |
| M/B * | 0 a | 1 ± 0.26 (10.71%) b |
| Vitrified embryos/per donor | 0 a | 0.83 ± 0 (9.09%) b |
| Recipient Number | Day after Estrus | Stage of Embryo at 9 dpf */Grade | Ovary/Corpus Luteum | Estrus after ET | Pregnant by USG (41 dpf ***) | Progesterone (ng/mL) (Pregnant 1–10 ng/mL) (43 dpf) | PAG (Pregnant > 0.3) (43 dpf) | Pregnant by USG (86 dpf) |
|---|---|---|---|---|---|---|---|---|
| 1 | 9 | WB/3 | L3 | present | negative | 0.6 | 0.035 | negative |
| 2 | 8.5 | M/2 | L3 | present | negative | 0.5 | 0.02 | negative |
| 3 | 9 | WB/1 | R2 | absent | positive | 9.9 | 0.31 | negative |
| 4 | 9.25 | WB/1 | L2 | absent | positive | 6.5 | 0.32 | negative |
| 5 | 9 | WB/1 | L2 | absent | positive | 6.5 | 3.06 | negative |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duszewska, A.M.; Baraniewicz-Kołek, M.; Wojdan, J.; Barłowska, K.; Bielecki, W.; Gręda, P.; Niżański, W.; Olech, W. Establishment of a Wisent (Bison bonasus) Germplasm Bank. Animals 2022, 12, 1239. https://doi.org/10.3390/ani12101239
Duszewska AM, Baraniewicz-Kołek M, Wojdan J, Barłowska K, Bielecki W, Gręda P, Niżański W, Olech W. Establishment of a Wisent (Bison bonasus) Germplasm Bank. Animals. 2022; 12(10):1239. https://doi.org/10.3390/ani12101239
Chicago/Turabian StyleDuszewska, Anna Maria, Magdalena Baraniewicz-Kołek, Jarosław Wojdan, Katarzyna Barłowska, Wojciech Bielecki, Paweł Gręda, Wojciech Niżański, and Wanda Olech. 2022. "Establishment of a Wisent (Bison bonasus) Germplasm Bank" Animals 12, no. 10: 1239. https://doi.org/10.3390/ani12101239
APA StyleDuszewska, A. M., Baraniewicz-Kołek, M., Wojdan, J., Barłowska, K., Bielecki, W., Gręda, P., Niżański, W., & Olech, W. (2022). Establishment of a Wisent (Bison bonasus) Germplasm Bank. Animals, 12(10), 1239. https://doi.org/10.3390/ani12101239