
Association of Aberrant DNA Methylation Level in the CD4 and JAK-STAT-Pathway-Related Genes with Mastitis Indicator Traits in Chinese Holstein Dairy Cattle



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
2.2. DNA and RNA Extraction, cDNA Synthesis, and Real-Time Quantitative PCR
2.3. Bisulfite Treatment of Extracted DNA and Hot Start PCR
2.4. Quantitative DNA Methylation Evaluation Using Pyrosequencing
2.5. Statistical Analysis
3. Results
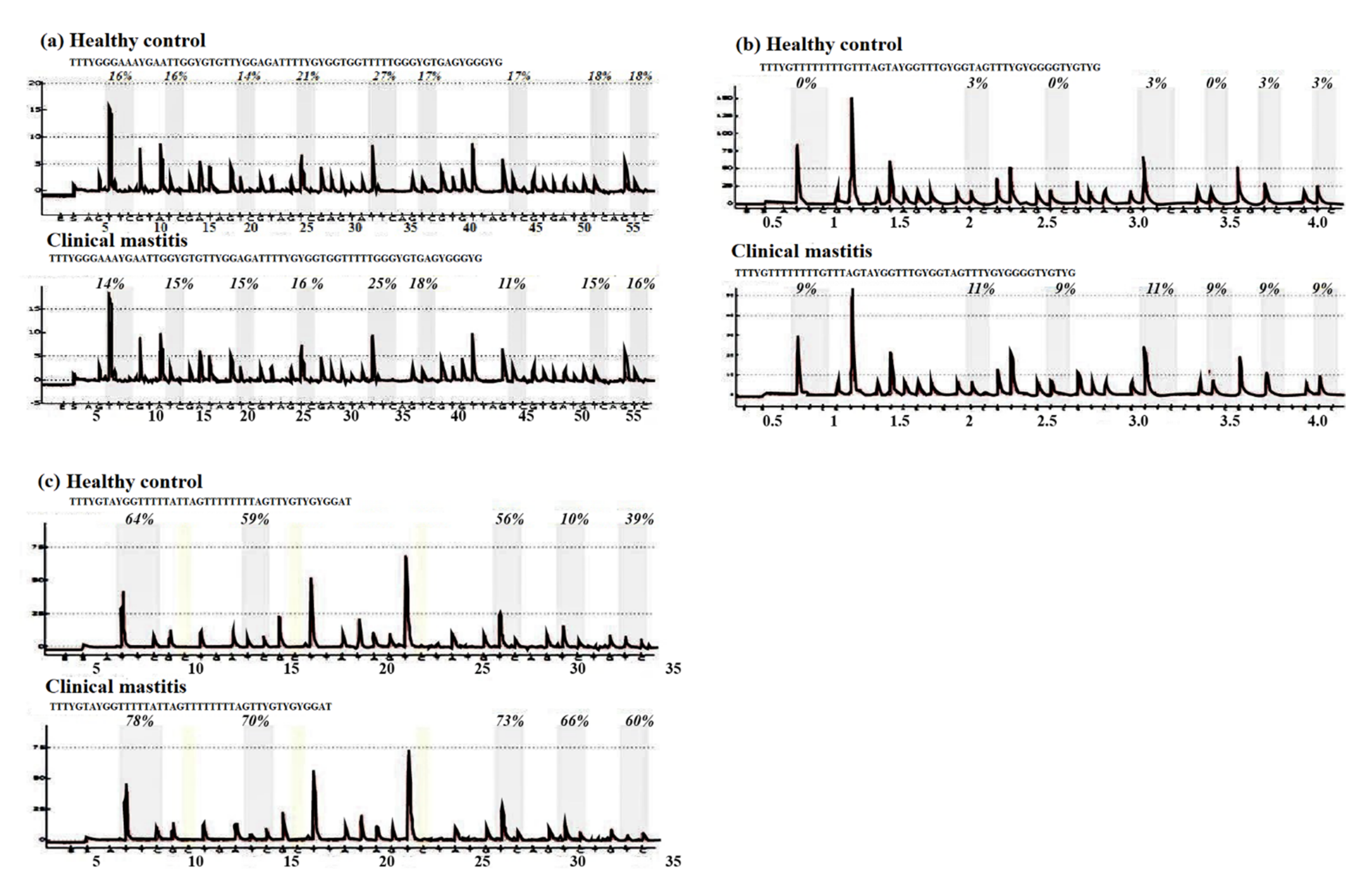
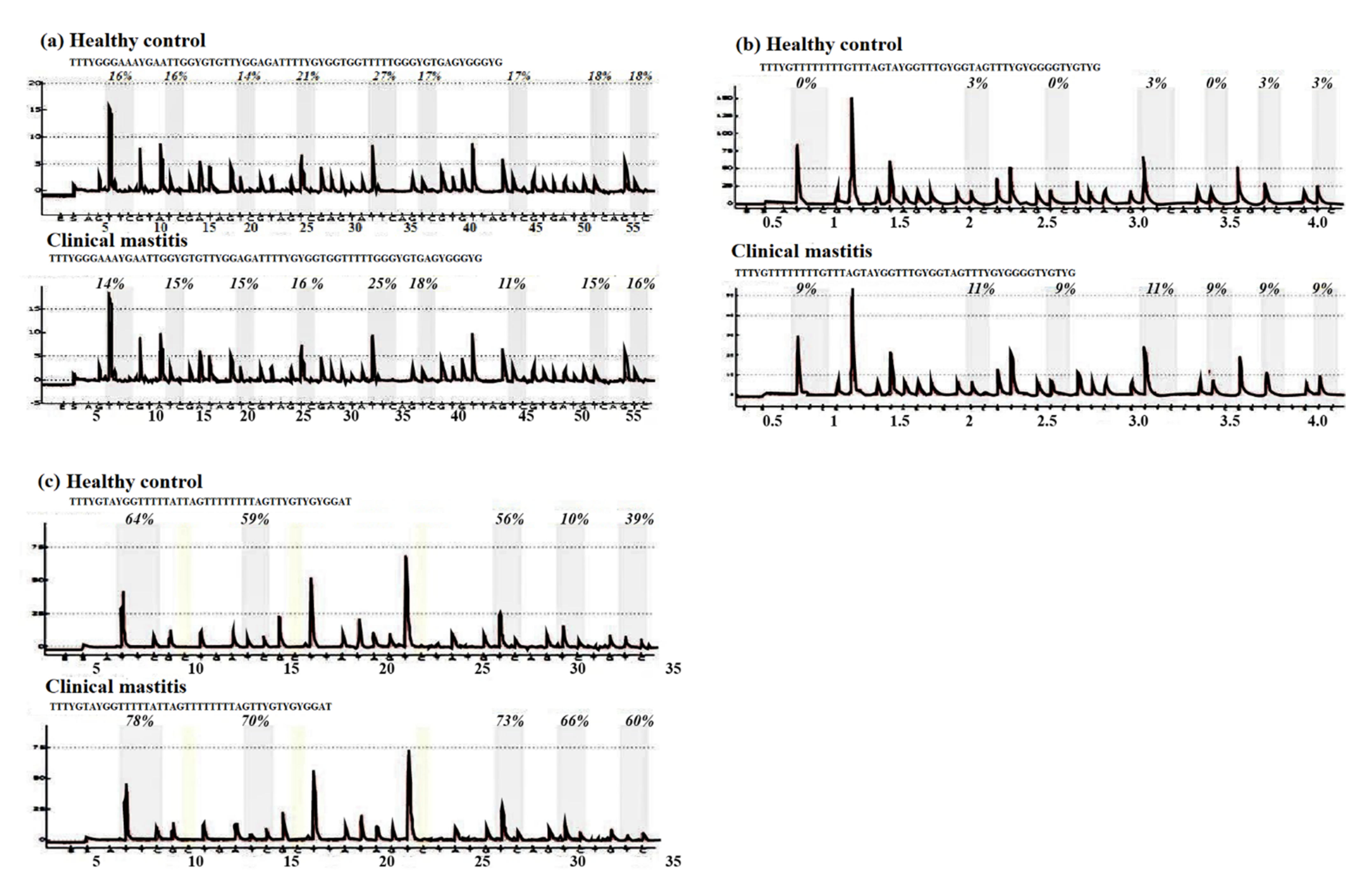
3.1. CpG Sites Methylation in the Genes under Study
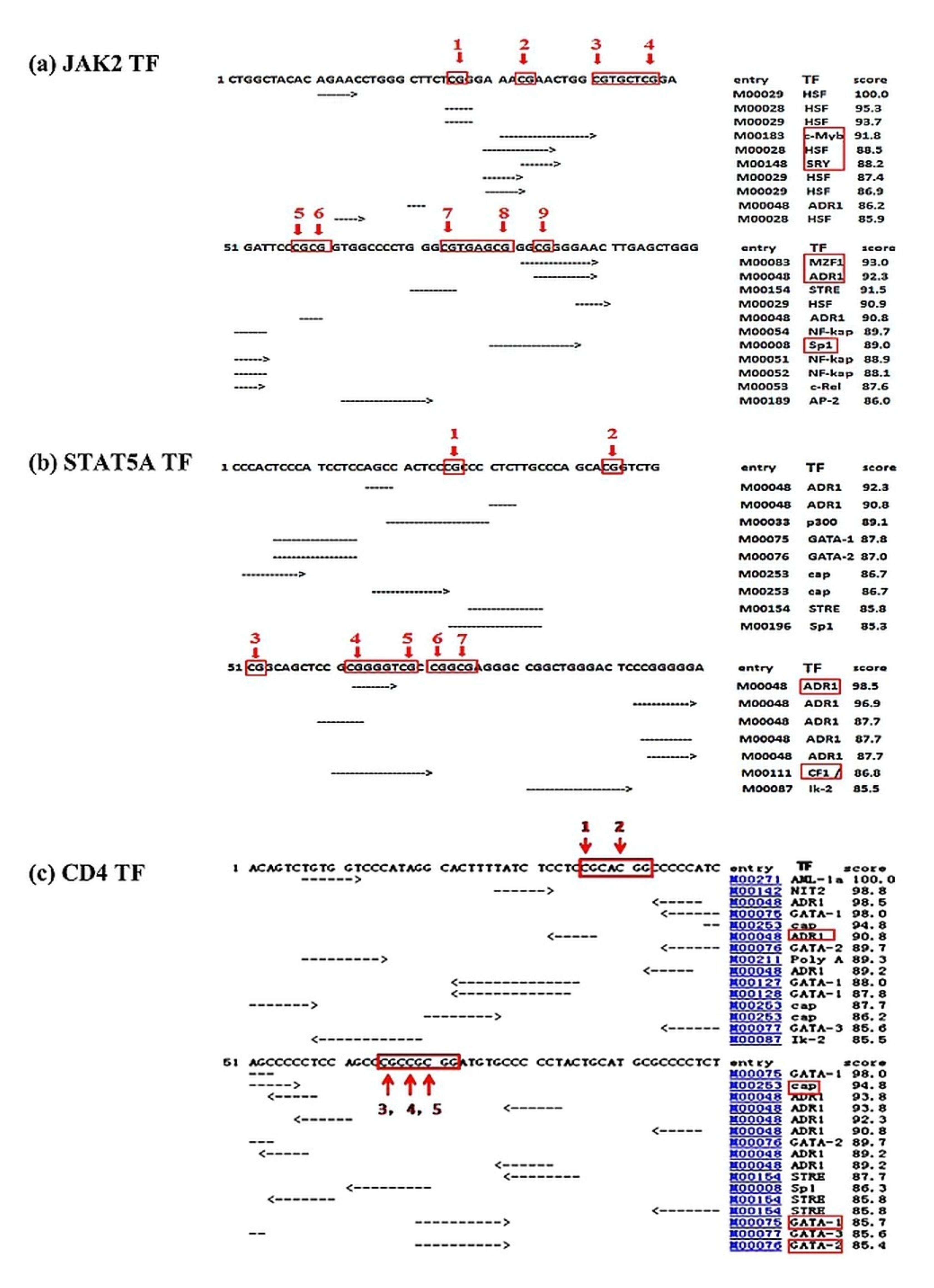
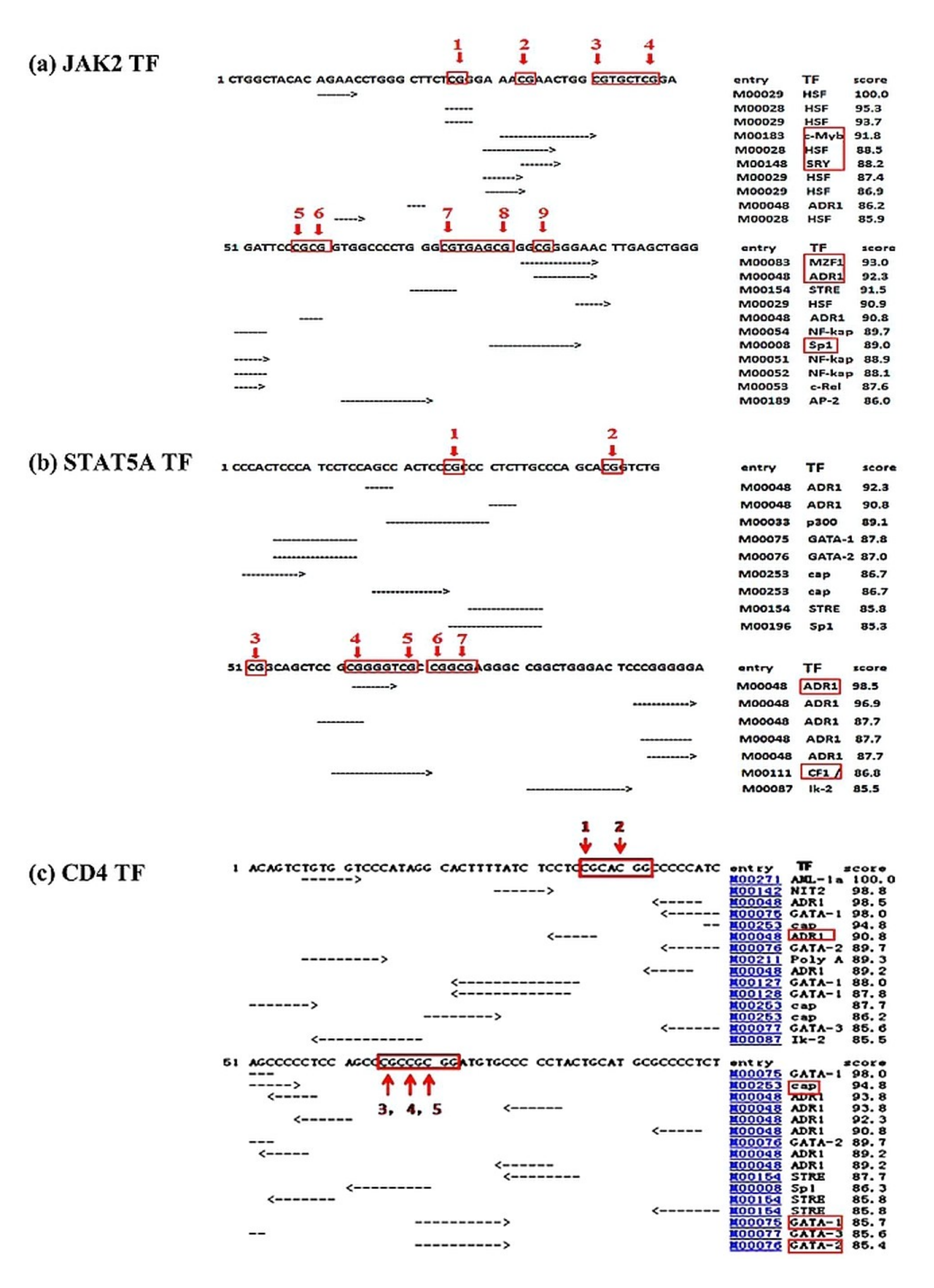
3.2. Predicted Binding Sites of TFs in the Promoter Regions of the Studied Genes
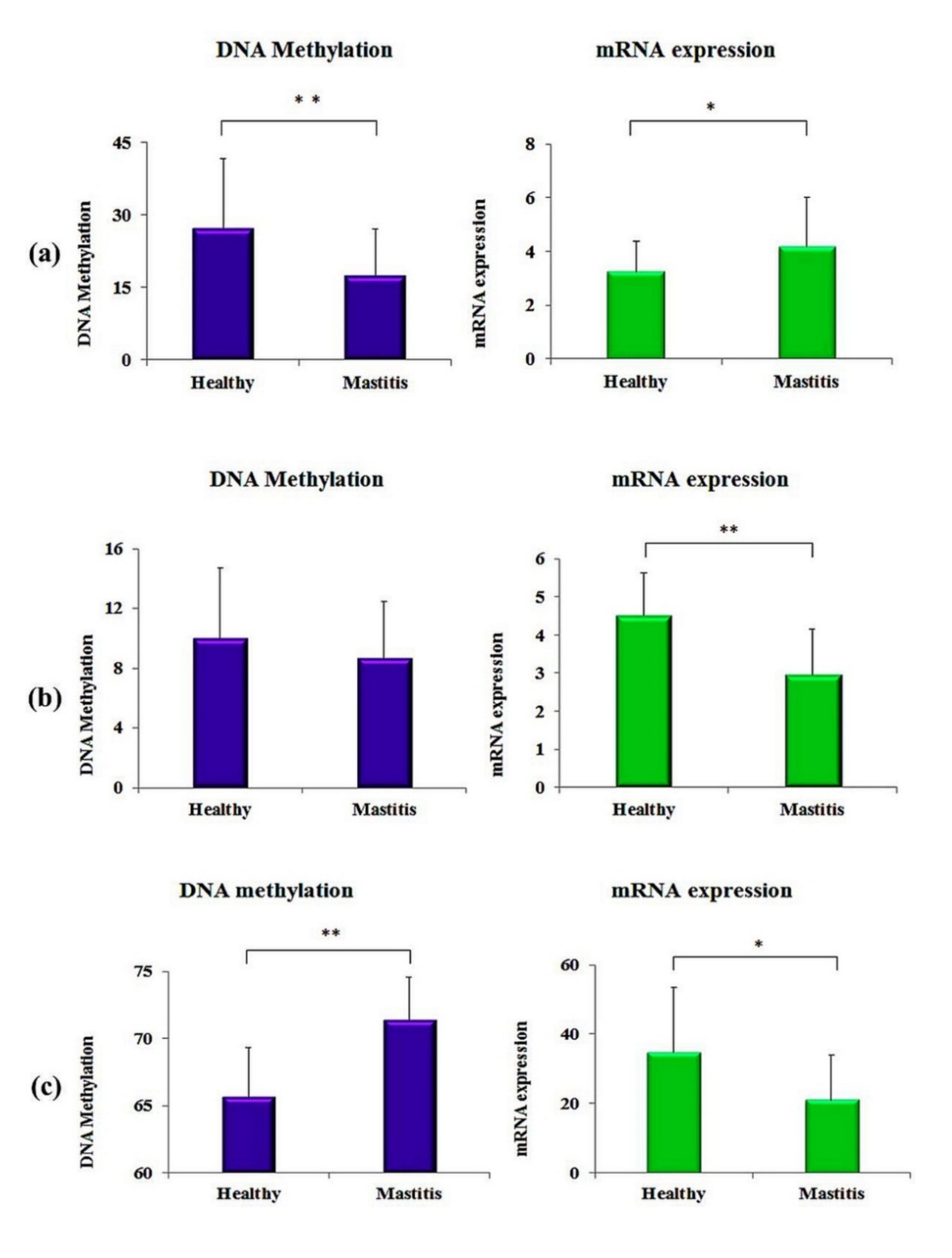
3.3. Relationship of DNA Methylation with mRNA Expression
3.4. Correlation among the CpG Sites in the Genes under Study
3.5. Methylation Correlation with Mastitis Traits in the Studied Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Usman, T.; Yu, Y.; Liu, C.; Wang, X.; Zhang, Q.; Wang, Y. Genetic Effects of Single Nucleotide Polymorphisms in JAK2 and STAT5A Genes on Susceptibility of Chinese Holsteins to Mastitis. Mol. Biol. Rep. 2014, 41, 8293–8301. [Google Scholar] [CrossRef] [PubMed]
- Hogeveen, H.; Huijps, K.; Lam, T.J. Economic aspects of mastitis: New developments. N. Z. Vet. J. 2011, 59, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Hameed, K.G.A.; Sender, G.; Korwin-Kossakowska, A. Public Health Hazard Due to Mastitis in Dairy Cows. Anim. Sci. Pap. Rep. 2007, 25, 73–85. [Google Scholar]
- Zeb, S.; Ali, N.; Niaz, S.; Rasheed, A.; Khattak, I.; Khan, N.U.; Wang, Y.; Usman, T. Association of SNPs in the Coding Regions of CD4 Gene with Mastitis Susceptibility and Production Traits in Dairy Cattle. Thai J. Vet. Med. 2020, 50, 75–80. [Google Scholar]
- Ali, N.; Niaz, S.; Khan, N.U.; Gohar, A.; Khattak, I.; Dong, Y.; Khattak, T.; Ahmad, I.; Wang, Y.; Usman, T. Polymorphisms in JAK2 Gene Are Associated with Production Traits and Mastitis Resistance in Dairy Cattle. Ann. Anim. Sci. 2020, 20, 409–423. [Google Scholar] [CrossRef]
- Usman, T.; Wang, Y.; Yu, Y.; Liu, C.; Wang, X.; Zhang, Y. Association Study of Single Nucleotide Polymorphisms in STAT5A/B with Mastitis Susceptibility in Chinese Holstein Cattle. In Proceedings of the 10th World Congress of Genetics Applied to Livestock Production, Vancouver, BC, Canada, 17–22 August 2014. [Google Scholar]
- Rasheed, A.; Ali, N.; Niaz, S.; Zeb, S.; Khattak, I.; Khan, A.; Ahmad, I.; Wang, Y.; Usman, T. Single Nucleotide Polymorphisms in the Promoter of CD4 Gene Are Associated with Production and Mastitis Traits in Dairy Cattle. Pak. Vet. J. 2020, 40, 394–396. [Google Scholar]
- Nowacka-Zawisza, M.; Wiśnik, E. DNA Methylation and Histone Modifications as Epigenetic Regulation in Prostate Cancer. Oncol. Rep. 2017, 38, 2587–2596. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, M. DNA Hypermethylation in Disease: Mechanisms and Clinical Relevance. Epigenetics 2019, 14, 1141–1163. [Google Scholar] [CrossRef] [Green Version]
- Robertson, K.D. DNA Methylation and Human Disease. Nat. Rev. Genet. 2005, 6, 597–610. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, J.; Sun, Y.; Dong, X.; Wang, Z.; Chen, J.; Dong, G. PGN and LTA from Staphylococcus aureus Induced Inflammation and Decreased Lactation through Regulating DNA Methylation and Histone H3 Acetylation in Bovine Mammary Epithelial Cells. Toxins 2020, 12, 238. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.; Kambhampati, S.; Parmar, S.; Platanias, L.C. Jak Family of Kinases in Cancer. Cancer Metastasis Rev. 2003, 22, 423–434. [Google Scholar] [CrossRef]
- Shuai, K.; Liu, B. Regulation of JAK-STAT Signalling in the Immune System. Nat. Rev. Immunol. 2003, 3, 900–911. [Google Scholar] [CrossRef]
- Usman, T.; Wang, Y.; Liu, C.; Wang, X.; Zhang, Y.; Yu, Y. Association Study of Single Nucleotide Polymorphisms in JAK 2 and STAT 5B Genes and Their Differential MRNA Expression with Mastitis Susceptibility in Chinese Holstein Cattle. Anim. Genet. 2015, 46, 371–380. [Google Scholar] [CrossRef]
- Pérez, C.; Pascual, M.; Martín-Subero, J.I.; Bellosillo, B.; Segura, V.; Delabesse, E.; Álvarez, S.; Larrayoz, M.J.; Rifón, J.; Cigudosa, J.C. Aberrant DNA Methylation Profile of Chronic and Transformed Classic Philadelphia-Negative Myeloproliferative Neoplasms. Haematologica 2013, 98, 1414–1420. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.S.; Zhang, Y.; He, Y.H.; Ma, P.P.; Fan, L.J.; Wang, Y.C.; Zhang, Y.I.; Sun, D.X.; Zhang, S.L.; Wang, C.D. Aberrant Promoter Methylation of the CD4 Gene in Peripheral Blood Cells of Mastitic Dairy Cows. Genet. Mol. Res. 2013, 12, 6228–6239. [Google Scholar] [CrossRef]
- Day, T.K.; Bianco-Miotto, T. Common Gene Pathways and Families Altered by DNA Methylation in Breast and Prostate Cancers. Endocr.-Relat. Cancer 2013, 20, R215–R232. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Costello, J. DNA Methylation: An Epigenetic Mark of Cellular Memory. Exp. Mol. Med. 2017, 49, e322. [Google Scholar] [CrossRef] [Green Version]
- Lesch, B.J.; Tothova, Z.; Morgan, E.A.; Liao, Z.; Bronson, R.T.; Ebert, B.L.; Page, D.C. Intergenerational epigenetic inheritance of cancer susceptibility in mammals. eLife 2019, 8, e39380. [Google Scholar] [CrossRef]
- Skinner, M.K.; Ben, M.M.; Sadler-Riggleman, I.; Beck, D.; Nilsson, E.; McBirney, M.; Klukovich, R.; Xie, Y.; Tang, C.; Yan, W. Alterations in sperm DNA methylation, non-coding RNA and histone retention associate with DDT-induced epigenetic transgenerational inheritance of disease. Epigenet. Chromatin 2018, 11, 8. [Google Scholar] [CrossRef] [Green Version]
- McRae, A.F.; Powell, J.E.; Henders, A.K.; Lisa, B.; Gibran, H.; Sonia, S.; Jodie, N.P.; Nicholas, G.M.; Peter, M.V.; Grant, W.M. Contribution of genetic variation to transgenerational inheritance of DNA methylation. Genome Biol. 2014, 15, 73. [Google Scholar] [CrossRef] [Green Version]
- Usman, T.; Wang, Y.; Liu, C.; He, Y.; Wang, X.; Dong, Y.; Wu, H.; Liu, A.; Ying, Y. Novel SNPs in IL-17F and IL-17A genes associated with somatic cell count in Chinese Holstein and Inner-Mongolia Sanhe cattle. J. Anim. Sci. Biotechnol. 2017, 8, 5. [Google Scholar] [CrossRef] [Green Version]
- Michael, K.; Udvardi, T.C.; Wolf-Rüdiger, S.E. Golden Rules of Quantitative RT-PCR. Plant Cell 2008, 20, 1736–1737. [Google Scholar]
- Yu, Y.; Zhang, H.; Tian, F.; Zhang, W.; Fang, H.; Song, J. An Integrated Epigenetic and Genetic Analysis of DNA Methyltransferase Genes (DNMT s) in Tumor Resistant and Susceptible Chicken Lines. PLoS ONE 2008, 3, e2672. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, H.; Tian, F.; Bacon, L.; Zhang, Y.; Zhang, W.; Song, J. Quantitative Evaluation of DNA Methylation Patterns for ALVE and TVB Genes in a Neoplastic Disease Susceptible and Resistant Chicken Model. PLoS ONE 2008, 3, e1731. [Google Scholar] [CrossRef]
- Widschwendter, M.; Apostolidou, S.; Raum, E.; Rothenbacher, D.; Fiegl, H.; Menon, U.; Stegmaier, C.; Jacobs, I.J.; Brenner, H. Epigenotyping in Peripheral Blood Cell DNA and Breast Cancer Risk: A Proof of Principle Study. PLoS ONE 2008, 3, e2656. [Google Scholar] [CrossRef]
- Wang, M.; Liang, Y.; Ibeagha-Awemu, E.M.; Li, M.; Zhang, H.; Chen, Z.; Sun, Y.; Karrow, N.A.; Yang, Z.; Mao, Y. Genome-Wide DNA methylation analysis of mammary gland tissues from Chinese Holstein cows with Staphylococcus aureus induced mastitis. Front. Genet. 2020, 11, 1295–1301. [Google Scholar] [CrossRef]
- Song, M.; He, Y.; Zhou, H.; Zhang, Y.; Li, X.; Yu, Y. Combined analysis of DNA methylome and transcriptome reveal novel candidate genes with susceptibility to bovine Staphylococcus aureus subclinical mastitis. Sci. Rep. 2016, 6, 29390. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, D.S.; Li, J.; Dawson, M.A.; Trotter, M.W.; Cheng, Y.-H.; Smith, A.M.; Mansfield, W.; Liu, P.; Kouzarides, T.; Nichols, J. LIF-Independent JAK Signalling to Chromatin in Embryonic Stem Cells Uncovered from an Adult Stem Cell Disease. Nat. Cell Biol. 2011, 13, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Sajjanar, B.; Trakooljul, N.; Wimmers, K.; Siriluck, P. DNA methylation analysis of porcine mammary epithelial cells reveals differentially methylated loci associated with immune response against Escherichia coli challenge. BMC Genom. 2019, 20, 623. [Google Scholar] [CrossRef]
- Li, W.; Sun, W.; Liu, L.; Yang, F.; Li, Y.; Chen, Y.; Fang, J.; Zhang, W.; Wu, J.; Zhu, Y. IL-32: A Host Proinflammatory Factor against Influenza Viral Replication Is Upregulated by Aberrant Epigenetic Modifications during Influenza A Virus Infection. J. Immunol. 2010, 185, 5056–5065. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, L.; Hagiwara, N.; Knowlton, A.A. Regulation of Heat Shock Protein 60 and 72 Expression in the Failing Heart. J. Mol. Cell. Cardiol. 2010, 48, 360–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.-J.; Wei, Y.-P.; Zhong, J.-C.; Chen, Z.-H.; Lu, H.; Tong, Z.-B. Sequence Characterization of the 5’-Flanking Region of the GHR Gene in Tibetan Sheep. Yi Chuan Hered 2007, 29, 963–971. [Google Scholar] [CrossRef] [PubMed]
- Stefanowicz, D.; Hackett, T.-L.; Garmaroudi, F.S.; Günther, O.P.; Neumann, S.; Sutanto, E.N.; Ling, K.-M.; Kobor, M.S.; Kicic, A.; Stick, S.M. DNA Methylation Profiles of Airway Epithelial Cells and PBMCs from Healthy, Atopic and Asthmatic Children. PLoS ONE 2012, 7, e44213. [Google Scholar]
- Zhang, Q.; Wang, H.Y.; Liu, X.; Wasik, M.A. STAT5A Is Epigenetically Silenced by the Tyrosine Kinase NPM1-ALK and Acts as a Tumor Suppressor by Reciprocally Inhibiting NPM1-ALK Expression. Nat. Med. 2007, 13, 1341–1348. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Yu, Y.; Zhang, H.; Tian, F.; Chang, S.; Cheng, H.H.; Song, J. Down-Regulation of Promoter Methylation Level of CD4 Gene after MDV Infection in MD-Susceptible Chicken Line. BMC Proc. 2011, 5, S7. [Google Scholar] [CrossRef] [Green Version]
- Landolfi, M.M.T.; Scollay, R.; Parnes, J.R. Specific Demethylation of the CD4 Gene during CD4 T Lymphocyte Differentiation. Mol. Immunol. 1997, 34, 53–61. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Clinical Mastitis | Healthy Control | |
|---|---|---|
| Number | 58 | 60 |
| SCC * | >500,000 | <200,000 |
| Parity range | 1–5 | 1–5 |
| CpG Position | Gene | Clinical Mastitis (Mean ± SE) | Healthy Control (Mean ± SE) | p Value |
|---|---|---|---|---|
| CpG site 1 | JAK2 | 15.6 ± 10.1 | 25.9 ± 15.7 | 0.05 |
| STAT5A | 10.3 ± 5.7 | 12.9 ± 12.5 | 0.40 | |
| CD4 | 82.7 ± 7.8 | 75.8 ± 5.3 | 4 × 10−3 | |
| CpG site 2 | JAK2 | 14.7 ± 6.4 | 25.5 ± 11.5 | 3 × 10−4 |
| STAT5A | 8.9 ± 10.2 | 10.1 ± 6.9 | 0.02 | |
| CD4 | 68.6 ± 3.0 | 66 ± 2.5 | 7 × 10−3 | |
| CpG site 3 | JAK2 | 13.9 ± 5.6 | 20.9 ± 9.9 | 3 × 10−3 |
| STAT5A | 8.8 ± 1.7 | 12.6 ± 8.2 | 0.15 | |
| CD4 | 77.8 ± 4 | 69.5 ± 5 | 2 × 10−5 | |
| CpG site 4 | JAK2 | 19 ± 12 | 36.5 ± 22 | 4 × 10−3 |
| STAT5A | 10.3 ± 5.6 | 10.7 ± 11.9 | 0.50 | |
| CD4 | 70.1 ± 6.3 | 62.1 ± 3.9 | 1 × 10−4 | |
| CpG site 5 | JAK2 | 25.9 ± 17.7 | 46.6 ± 28.5 | 6 × 10−3 |
| STAT5A | 5.5 ± 1.2 | 7.2 ± 1.2 | 0.02 | |
| CD4 | 57.8 ± 3.3 | 54.9 ± 5.7 | 0.05 | |
| CpG site 6 | JAK2 | 14.8 ± 6.7 | 23.9 ± 10.5 | 2 × 10−3 |
| STAT5A | 7.4 ± 1.5 | 9 ± 2.2 | 0.01 | |
| CpG site 7 | JAK2 | 15.2 ± 8 | 21.8 ± 8.5 | 0.01 |
| STAT5A | 8.5 ± 5.9 | 15.3 ± 5.5 | 0.11 | |
| CpG site 8 | JAK2 | 17.2 ± 9 | 24.3 ± 9.6 | 7 × 10−3 |
| CpG site 9 | JAK2 | 19.6 ± 8.5 | 24.1 ± 11.7 | 0.07 |
| All (Mean) | JAK2 | 17.5 ± 9.6 | 27.3 ± 14.3 | 3 × 10−3 |
| STAT5A | 8.7 ± 3.8 | 10 ± 5.7 | 0.20 | |
| CD4 | 71.4 ± 3.2 | 65.7 ± 3.7 | 5 × 10−5 |
| Traits | JAK2 Methylation | STAT5A Methylation | CD4 Methylation |
|---|---|---|---|
| SCC | −0.06 | 0.17 | 0.67 ** |
| p value | 0.55 | 0.33 | 5 × 10−4 |
| SCS | −0.05 | 0.15 | 0.71 ** |
| p value | 0.61 | 0.41 | 3 × 10−5 |
| Status | −0.13 | −0.24 | 0.66 ** |
| p value | 0.18 | 0.16 | 2 × 10−5 |
| IL-4 | 0.28 * | −0.48 * | −0.15 |
| p value | 0.03 | 0.03 | 0.51 |
| IL-6 | −0.04 | −0.51* | −0.10 |
| p value | 0.73 | 0.03 | 0.65 |
| IL-10 | −0.08 | 0.14 | −0.24 |
| p value | 0.56 | 0.56 | 0.31 |
| IL-17 | −0.19 | 0.44 * | 0.15 |
| p value | 0.15 | 0.05 | 0.53 |
| TNF-α | 0.07 | 0.08 | −0.22 |
| p value | 0.63 | 0.73 | 0.36 |
| IFN-γ | 0.11 | −0.01 | −0.54 ** |
| p value | 0.41 | 0.95 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Usman, T.; Ali, N.; Wang, Y.; Yu, Y. Association of Aberrant DNA Methylation Level in the CD4 and JAK-STAT-Pathway-Related Genes with Mastitis Indicator Traits in Chinese Holstein Dairy Cattle. Animals 2022, 12, 65. https://doi.org/10.3390/ani12010065
Usman T, Ali N, Wang Y, Yu Y. Association of Aberrant DNA Methylation Level in the CD4 and JAK-STAT-Pathway-Related Genes with Mastitis Indicator Traits in Chinese Holstein Dairy Cattle. Animals. 2022; 12(1):65. https://doi.org/10.3390/ani12010065
Chicago/Turabian StyleUsman, Tahir, Nawab Ali, Yachun Wang, and Ying Yu. 2022. "Association of Aberrant DNA Methylation Level in the CD4 and JAK-STAT-Pathway-Related Genes with Mastitis Indicator Traits in Chinese Holstein Dairy Cattle" Animals 12, no. 1: 65. https://doi.org/10.3390/ani12010065
APA StyleUsman, T., Ali, N., Wang, Y., & Yu, Y. (2022). Association of Aberrant DNA Methylation Level in the CD4 and JAK-STAT-Pathway-Related Genes with Mastitis Indicator Traits in Chinese Holstein Dairy Cattle. Animals, 12(1), 65. https://doi.org/10.3390/ani12010065