
Beclin 1, LC3 and P62 Expression in Equine Sarcoids

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tumor Samples
2.2. Immunohistochemistry
2.3. Scoring of Immunoreactivity
2.4. Western Blot (WB)
2.5. Statistical Analysis
3. Results
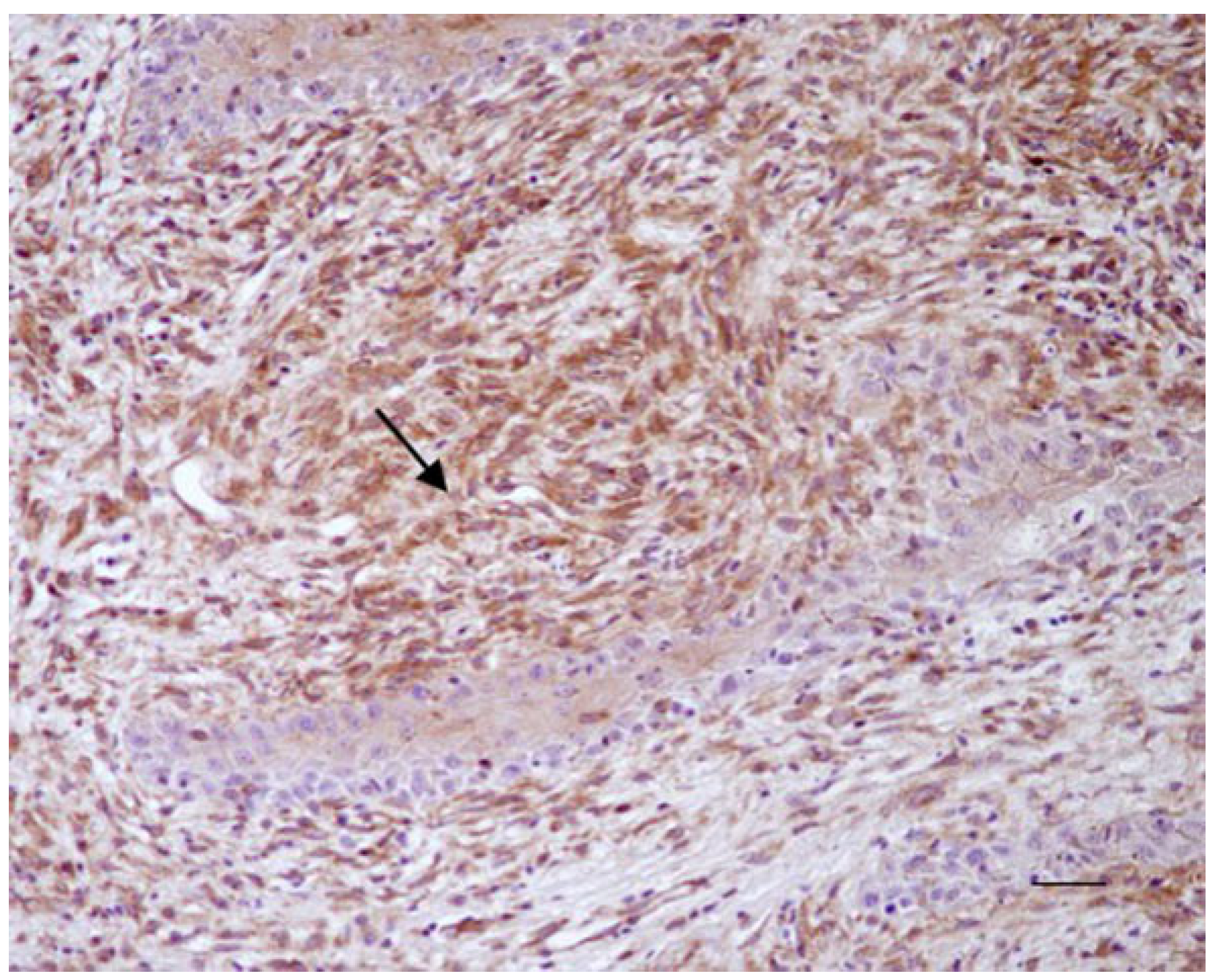
3.1. Histological and Immunohistochemical Results
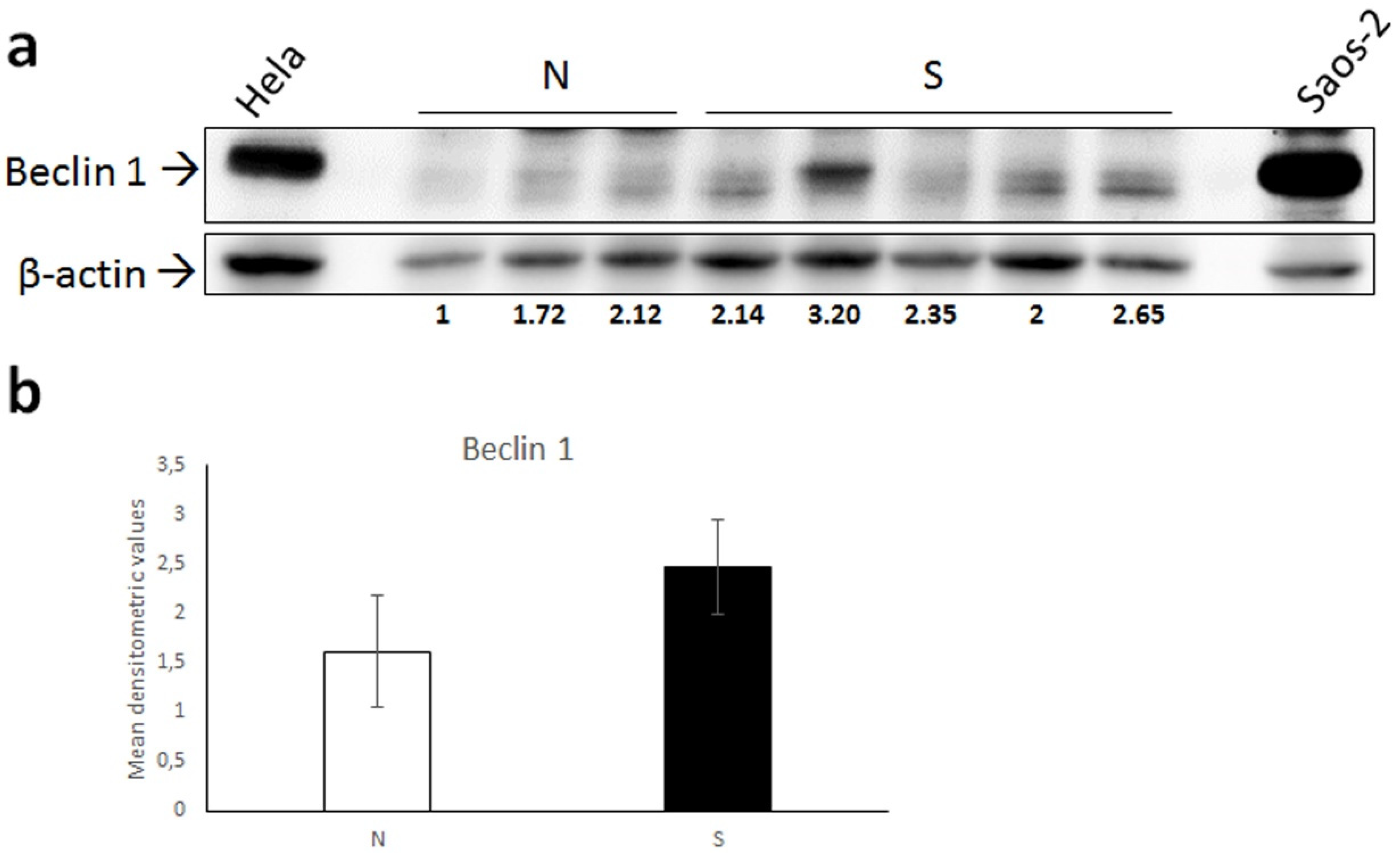
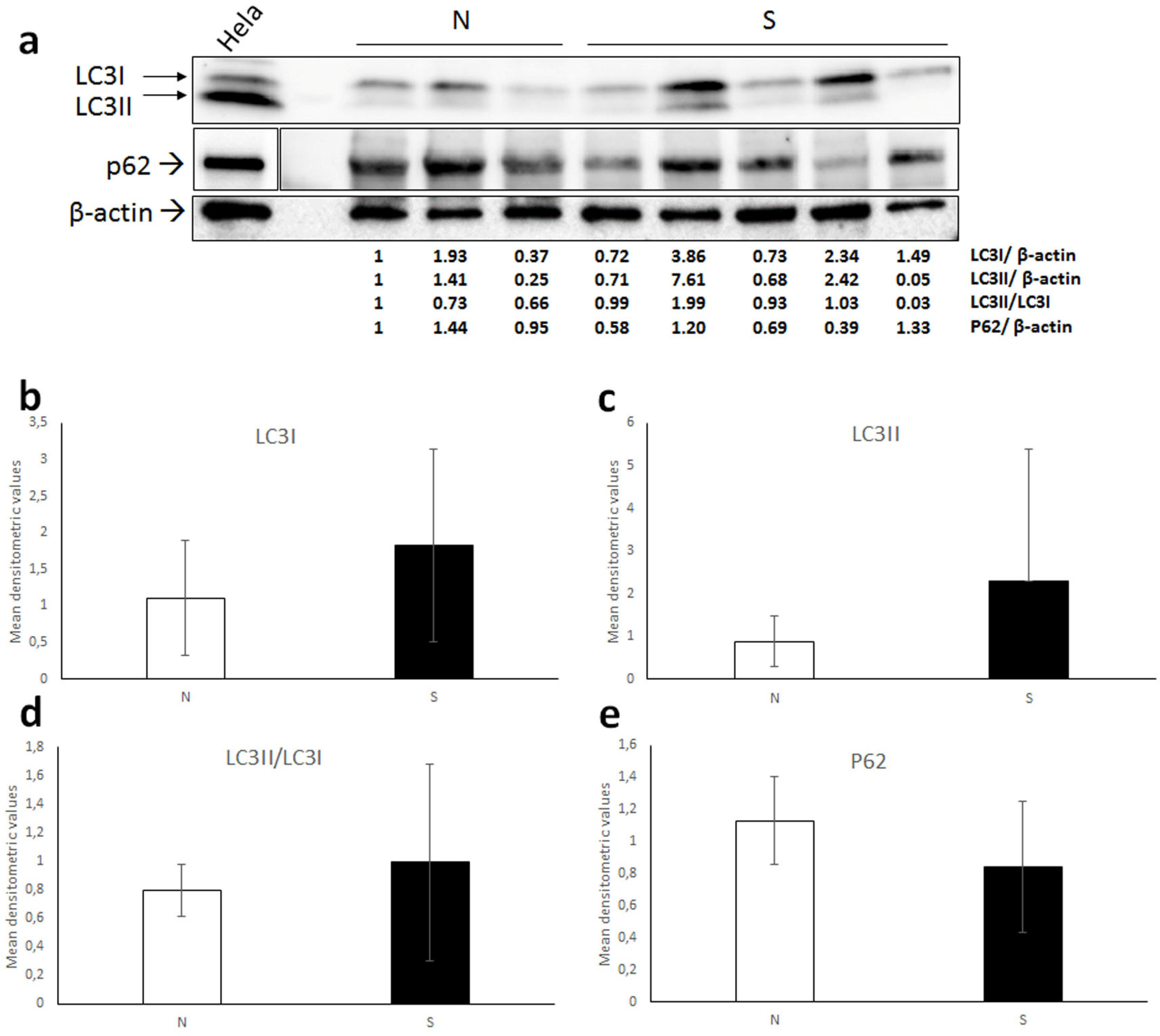
3.2. Biochemical Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
List of Abbreviations
References
- Rothman, S. How is the balance between protein synthesis and degradation achieved? Theor. Biol. Med. Model. 2010, 7, 25. [Google Scholar] [CrossRef] [Green Version]
- Klionsky, D.J.; Emr, S.D. Autophagy as a regulated pathway of cellular degradation. Science 2000, 290, 1717–1721. [Google Scholar] [CrossRef]
- Kroemer, G. Autophagy: A druggable process that is deregulated in aging and human disease. J. Clin. Investig. 2015, 125, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Dikic, I. Proteasomal and autophagic degradation systems. Ann. Rev. Biochem. 2017, 86, 193–224. [Google Scholar] [CrossRef] [PubMed]
- Parzych, K.R.; Klionsky, D.J. An Overview of Autophagy: Morphology, Mechanism, and Regulation. Antioxid. Redox Signal. 2014, 20, 460–473. [Google Scholar] [CrossRef] [Green Version]
- Tooze, S.A.; Schiavo, G. Liaisons dangereuses: Autophagy, neuronal survival and neurodegeneration. Curr. Opin. Neurobiol. 2008, 18, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Münz, C. Enhancing immunity through autophagy. Annu. Rev. Immunol. 2009, 27, 423–449. [Google Scholar] [CrossRef]
- Tukaj, C. The significance of macroautophagy in health and disease. Folia Morphol. 2013, 72, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Itakura, E.; Kishi, C.; Inoue, K.; Mizushima, N. Beclin 1 forms two distinct phosphatidylinositol 3-kinase complexes with mammalian Atg14 and UVRAG. Mol. Biol. Cell 2008, 19, 5360–5372. [Google Scholar] [CrossRef] [Green Version]
- Kabeya, Y.; Mizushima, N.; Ueno, T.; Yamamoto, A.; Kirisako, T.; Noda, T.; Kominami, E.; Ohsumi, Y.; Yoshimori, T. LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. EMBO J. 2000, 19, 5720–5728. [Google Scholar] [CrossRef]
- Pankiv, S.; Clausen, T.H.; Lamark, T.; Brech, A.; Bruun, J.A.; Outzen, H.; Øvervatn, A.; Bjørkøy, G.; Johansen, T. p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of ubiquitinated protein aggregates by autophagy. J. Biol. Chem. 2007, 282, 24131–24145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavandero, S.; Chiong, M.; Rothermel, B.A.; Hill, J.A. Autophagy in cardiovascular biology. J. Clin. Investig. 2015, 125, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Frake, R.A.; Ricketts, T.; Menzies, F.M.; Rubinsztein, D.C. Autophagy and neurodegeneration. J. Clin. Investig. 2015, 125, 65–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, E. The role for autophagy in cancer. J. Clin. Investig. 2015, 125, 42–46. [Google Scholar] [CrossRef] [Green Version]
- Jackson, W.T. Viruses and the autophagy pathway. Virology 2015, 479–480, 450–456. [Google Scholar] [CrossRef]
- Mattoscio, D.; Medda, A.; Chiocca, S. Human papilloma virus and autophagy. Int. J. Mol. Sci. 2018, 19, 1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roperto, S.; Russo, V.; De Falco, F.; Urraro, C.; Maiolino, P.; Del Piero, F.; Roperto, F. Bovine papillomavirus E5 oncoprotein expression and its association with an interactor network in aggresome-autophagy pathway. Vet. Microbiol. 2019, 233, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Roperto, S.; Russo, V.; Rosati, A.; Ceccarelli, D.M.; Munday, J.S.; Turco, M.C.; Roperto, F. Chaperone-assisted selective autophagy in healthy and papillomavirus-associated neoplastic urothelium of cattle. Vet. Microbiol. 2018, 221, 134–142. [Google Scholar] [CrossRef]
- Borzacchiello, G.; Roperto, F. Bovine papillomaviruses, papillomas and cancer in cattle. Vet. Res. 2008, 39, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunardi, M.; Alcântara, B.; Otonel, R.; Rodrigues, W.; Alfieri, A.; Alfieri, A. Bovine papillomavirus type 13 DNA in equine sarcoids. J. Clin. Microbiol. 2013, 51, 2167–2171. [Google Scholar] [CrossRef] [Green Version]
- Chambers, G.; Ellsmore, V.A.; O’Brien, P.M.; Reid, S.W.; Love, S.; Campo, M.S.; Nasir, L. Association of bovine papillomavirus with the equine sarcoid. J. Gen. Virol. 2003, 84, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Bogaert, L.; Martens, A.; Kast, W.M.; Van Marck, E.; De Cock, H. Bovine papillomavirus DNA can be detected in keratinocytesof equine sarcoid tumors. Vet. Microbiol. 2010, 146, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Marti, E.; Lazary, S.; Antczak, D.F.; Gerber, H. Report of the first international workshop on equine sarcoid. Equine Vet. J. 1993, 25, 397–407. [Google Scholar] [CrossRef]
- Ragland, W.L.; Keown, G.H.; Spencer, G.R. Equine sarcoid. Equine Vet. J. 1970, 2, 2–11. [Google Scholar] [CrossRef]
- Borzacchiello, G.; Corteggio, A. Equine sarcoid: State of the art. Ippologia 2009, 20, 7–14. [Google Scholar]
- Nasir, L.; Campo, M.S. Bovine papillomaviruses: Their role in the aetiology of cutaneous tumors of bovids and equids. Vet. Dermatol. 2008, 19, 243–254. [Google Scholar] [CrossRef]
- Nasir, L.; Brandt, S. Papillomavirus associated diseases of the horse. Vet. Microbiol. 2013, 167, 159–167. [Google Scholar] [CrossRef]
- Knottenbelt, D.C. A suggested clinical classification of the equine sarcoid. Clinical techniques in equine. Practice 2005, 4, 278–295. [Google Scholar]
- Cochrane, A.C. Models in vivo of wound healing in the horse and the role of growth factors. Vet. Dermatol. 1997, 8, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Hanson, R.R. Review: Complications of equine wound management and dermatologic surgery. Vet. Clin. N. Am. Equine Pract. 2008, 24, 663–696. [Google Scholar] [CrossRef] [PubMed]
- Martano, M.; Corteggio, A.; Restucci, B.; De Biase, M.E.; Borzacchiello, G.; Maiolino, P. Extracellular matrix remodeling in equine sarcoid: An immunohistochemical and molecular study. BMC Vet. Res. 2016, 12, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martano, M.; Power, K.; Restucci, B.; Pagano, I.; Altamura, G.; Borzacchiello, G.; Maiolino, P. Expression of vascular endothelial growth factor (VEGF) in equine sarcoid. BMC Vet. Res. 2018, 14, 266. [Google Scholar] [CrossRef] [PubMed]
- Martano, M.; Altamura, G.; Power, K.; Restucci, B.; Carella, F.; Borzacchiello, G.; Maiolino, P. Evaluation of Hypoxia-Inducible Factor-1 Alpha (HIF-1α) in Equine Sarcoid: An Immunohistochemical and Biochemical Study. Pathogens 2020, 9, 58. [Google Scholar] [CrossRef] [Green Version]
- Yun, C.W.; Lee, S.H. The Roles of Autophagy in Cancer. Int. J. Mol. Sci. 2018, 19, 3466. [Google Scholar] [CrossRef] [Green Version]
- Borzacchiello, G.; Russo, V.; Della Salda, L.; Roperto, S.; Roperto, F. Expression of platelet-derived growth factor-beta receptor and bovine papillomavirus E5 and E7 oncoproteins in equine sarcoid. J. Comp. Pathol. 2008, 139, 231–237. [Google Scholar] [CrossRef]
- He, J.M.; Liu, P.Y.; Wang, J. MicroRNA-17-5p regulates the growth, migration and invasion of the human osteosarcoma cells by modulating the expression of PTEN. J. BUON 2020, 25, 1028–1034. [Google Scholar]
- Katsuragi, Y.; Ichimura, Y.; Komatsu, M. p62/SQSTM1 functions as a signaling hub and an autophagy adaptor. FEBS J. 2015, 282, 4672–4678. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.-L.; Chen, F.-F.; Lung, J.; Lo, C.-H.; Lee, F.-H.; Lu, Y.-C.; Hung, C.-H. Prognostic significance of p62/SQSTM1 subcellular localization and LC3B in oral squamous cell carcinoma. Br. J. Cancer 2014, 111, 944–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega-Rubín-de-Celis, S. The Role of Beclin 1-Dependent Autophagy in Cancer. Biology 2020, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- Won, K.Y.; Kim, G.Y.; Kim, Y.W.; Song, J.Y.; Lim, S.J. Clinicopathologic correlation of beclin-1 and bcl-2 expression in human breast cancer. Hum. Pathol. 2010, 41, 107–112. [Google Scholar] [CrossRef]
- Jiang, Z.F.; Shao, L.J.; Wang, W.M.; Yan, X.B.; Liu, R.Y. Decreased expression of Beclin-1 and LC3 in human lung cancer. Mol. Biol. Rep. 2012, 39, 259–267. [Google Scholar] [CrossRef]
- Won, K.Y.; Kim, G.Y.; Lim, S.J.; Kim, Y.W. Decreased Beclin-1 expression is correlated with the growth of the primary tumor in patients with squamous cell carcinoma and adenocarcinoma of the lung. Hum. Pathol. 2012, 43, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.B.; Shi, Y.H.; Zhou, J.; Qiu, S.J.; Xu, Y.; Dai, Z.; Shi, G.M.; Wang, X.Y.; Ke, A.W.; Wu, B. Association of autophagy defect with a malignant phenotype and poor prognosis of hepatocellular carcinoma. Cancer Res. 2008, 68, 9167. [Google Scholar] [CrossRef] [Green Version]
- Miracco, C.; Cevenini, G.; Franchi, A.; Luzi, P.; Cosci, E.; Mourmouras, V.; Monciatti, I.; Mannucci, S.; Biagioli, M.; Toscano, M. Beclin 1 and LC3 autophagic gene expression in cutaneous melanocytic lesions. Hum. Pathol. 2010, 41, 503–512. [Google Scholar] [CrossRef]
- Ahn, C.H.; Jeong, E.; Lee, J.; Kim, M.; Kim, S.; Kim, S.; Yoo, N.; Lee, S. Expression of beclin-1, an autophagy-related protein, in gastric and colorectal cancers. Acta Pathol. Microbiol. Immunol. Scand. 2007, 115, 1344–1349. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, S.H.; Do, S.I.; Lim, S.J.; Park, Y.K.; Kim, Y.W. Clinicopathologic correlation of beclin-1 expression in pancreatic ductal adenocarcinoma. Pathol. Res. Pract. 2011, 207, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K. Pathologic Changes and Autophagy: New Insights for the Pathogenesis of Animal Diseases. Vet. Pathol. 2017, 54, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.L.; Chang, K.C.; Lo, C.C.; Chu, P.Y.; Lu, C.H. Expression of autophagy-related protein beclin-1 in malignant canine mammary tumors. BMC Vet. Res. 2013, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Akinduro, O.; Sully, K.; Patel, A.; Robinson, D.J.; Chik, A.; McPhail, G.; Braun, K.M.; Philpott, M.P.; Harwood, C.A.; Byrne, C.; et al. Constitutive Autophagy and Nucleophagy during Epidermal Differentiation. J. Investig. Dermatol. 2016, 136, 1460–1470. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.J.; Kim, H.P.; Jin, Y.; Choi, A.M.K.; Ryter, S.W. Beclin 1 deficiency is associated with increased hypoxia-induced angiogenesis. Autophagy 2011, 7, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Tan, Q.; Wang, M.; Yu, M.; Zhang, J.; Bristow, R.G.; Hill, R.P.; Tannock, I.F. Role of Autophagy as a Survival Mechanism for Hypoxic Cells in Tumors. Neoplasia 2016, 18, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Lu, Y.; Lu, C.; Zhang, L. Beclin-1 expression is a predictor of clinical outcome in patients with esophageal squamous cell carcinoma and correlated to hypoxia-inducible factor (HIF)-1alpha expression. Pathol. Oncol. Res. 2009, 15, 487–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranaghan, M.J.; Durney, M.A.; Mesleh, M.F.; McCarren, P.R.; Garvie, C.W.; Daniels, D.S.; Carey, K.L.; Skepner, A.P.; Levine, B.; Perez, J.R. The Autophagy-Related Beclin-1 Protein Requires the Coiled-Coil and BARA Domains to Form a Homodimer with Submicromolar Affinity. Biochemistry 2017, 56, 6639–6651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Principe, D.; Vona, R.; Giordani, L.; Straface, E.; Giammarioli, A.M. Defective autophagy in fibroblasts may contribute to fibrogenesis in autoimmune processes. Curr. Pharm. Des. 2011, 17, 3878–3887. [Google Scholar] [CrossRef]
- Martinez-Outschoorn, U.E.; Trimmer, C.; Lin, Z.; Whitaker-Menezes, D.; Chiavarina, B.; Zhou, J.; Wang, C.; Pavlides, S.; Martinez-Cantarin, M.P.; Capozza, F.; et al. Autophagy in cancer associated fibroblasts promotes tumor cell survival: Role of hypoxia, HIF1 induction and NFκB activation in the tumor stromal microenvironment. Cell Cycle 2010, 9, 3515–3533. [Google Scholar] [CrossRef] [PubMed]
- Everts, V.; van der Zee, E.; Creemers, L.; Beertsen, W. Phagocytosis and intracellular digestion of collagen, its role in turnover and remodelling. Histochem. J. 1996, 28, 229–245. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.H.; Xu, L.; Wang, Y.; Cao, M.Q.; Li, L.; Bai, T. Clinicopathologic correlations between human papillomavirus 16 infection and Beclin 1 expression in human cervical cancer. Int. J. Gynecol. Pathol. 2011, 30, 400–406. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Number of Cases | Staining Intensity Score * Beclin 1 | Percentage Positive Score ** Beclin 1 | Staining Intensity Score * LC3 | Percentage Positive Score ** LC3 | Staining Intensity Score * P62 | Percentage Positive Score ** P62 |
|---|---|---|---|---|---|---|---|
| Neck | 2 | ++ | 3 | +/− | 1 | ++ | 3 |
| Limb | 6 | ++ | 3 | +/− | 1 | ++ | 3 |
| 4 | + | 3 | +/− | 1 | ++ | 3 | |
| Abdomen | 4 | ++ | 3 | +/− | 1 | ++ | 3 |
| 2 | + | 3 | + | 2 | ++ | 3 | |
| Pectoral region | 5 | ++ | 3 | + | 2 | ++ | 3 |
| 3 | + | 3 | + | 2 | ++ | 3 | |
| Head | 5 | ++ | 3 | +/− | 1 | ++ | 3 |
| 1 | + | 3 | +/− | 1 | ++ | 3 | |
| (para)-genital region | 3 | ++ | 3 | + | 2 | ++ | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martano, M.; Altamura, G.; Power, K.; Liguori, P.; Restucci, B.; Borzacchiello, G.; Maiolino, P. Beclin 1, LC3 and P62 Expression in Equine Sarcoids. Animals 2022, 12, 20. https://doi.org/10.3390/ani12010020
Martano M, Altamura G, Power K, Liguori P, Restucci B, Borzacchiello G, Maiolino P. Beclin 1, LC3 and P62 Expression in Equine Sarcoids. Animals. 2022; 12(1):20. https://doi.org/10.3390/ani12010020
Chicago/Turabian StyleMartano, Manuela, Gennaro Altamura, Karen Power, Pierluigi Liguori, Brunella Restucci, Giuseppe Borzacchiello, and Paola Maiolino. 2022. "Beclin 1, LC3 and P62 Expression in Equine Sarcoids" Animals 12, no. 1: 20. https://doi.org/10.3390/ani12010020
APA StyleMartano, M., Altamura, G., Power, K., Liguori, P., Restucci, B., Borzacchiello, G., & Maiolino, P. (2022). Beclin 1, LC3 and P62 Expression in Equine Sarcoids. Animals, 12(1), 20. https://doi.org/10.3390/ani12010020