
Transboundary Animal Diseases, an Overview of 17 Diseases with Potential for Global Spread and Serious Consequences


Simple Summary
Abstract
1. Introduction
2. Methods
3. Review
3.1. African Horse Sickness
3.2. African Swine Fever
3.3. Avian Influenza
3.4. Bluetongue
3.5. Classical Swine Fever
3.6. Contagious Bovine Pleuropneumonia
3.7. Foot and Mouth Disease
3.8. Hemorrhagic Septicemia
3.9. Lumpy Skin Disease
3.10. Middle East Respiratory Syndrome
3.11. Newcastle Disease
3.12. Peste des Petits Ruminants
3.13. Rift Valley Fever
3.14. Rinderpest
3.15. Sheeppox and Goatpox
3.16. Swine Vesicular Disease
3.17. Vesicular Stomatitis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Organization for Animal Health. Animal Diseases; World Organization for Animal Health: Paris, France, 2021; Available online: www.oie.int/en/what-we-do/animal-health-and-welfare/animal-diseases/ (accessed on 16 May 2021).
- Food and Agriculture Organization of the United Nations. Transboundary Animal Diseases. 2021. Available online: www.fao.org/emergencies/emergency-types/transboundary-animal-diseases/en/ (accessed on 16 May 2021).
- Calkins, C.M.; Scasta, J.D. Transboundary Animal Diseases (Tads) Affecting Domestic and Wild African Ungulates: African Swine Fever, Foot and Mouth Disease, Rift Valley Fever (1996–2018). Res. Vet. Sci. 2020, 131, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Rossiter, P.B.; al Hammadi, N. Living with Transboundary Animal Diseases (TADs). Trop. Anim. Health Prod. 2008, 41, 999. [Google Scholar] [CrossRef] [PubMed]
- Torres-Velez, F.; Havas, K.A.; Spiegel, K.; Brown, C. Transboundary Animal Diseases as Re-Emerging Threats—Impact on One Health. Semin. Diagn. Pathol. 2019, 36, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.; Mellor, P.S.; Fall, A.G.; Garros, C.; Venter, G.J. African Horse Sickness Virus: History, Transmission, and Current Status. Annu. Rev. Entomol. 2017, 62, 343–358. [Google Scholar] [CrossRef]
- Mirchamsy, H.; Hazrati, A. A Review on Aetiology and Pathogeny of African Horsesickness. Arch. Razi. Inst. 1973, 25, 23–46. [Google Scholar]
- Roy, P.; Mertens, P.P.; Casal, I. African Horse Sickness Virus Structure. Comp. Immunol. Microbiol. Infect. Dis. 1994, 17, 243–273. [Google Scholar] [CrossRef]
- Wood, H.A. Viruses with Double-Stranded RNA Genomes. J. Gen. Virol. 1973, 20, 61–85. [Google Scholar] [CrossRef]
- Dennis, S.J.; Meyers, A.E.; Hitzeroth, I.; Rybicki, E.P. African Horse Sickness: A Review of Current Understanding and Vaccine Development. Viruses 2019, 11, 844. [Google Scholar] [CrossRef]
- Wall, G.V.; Wright, I.M.; Barnardo, C.; Erasmus, B.J.; van Staden, V.; Potgieter, A.C. African Horse Sickness Virus NS4 Protein Is an Important Virulence Factor and Interferes with JAK-STAT Signaling during Viral Infection. Virus Res. 2021, 298, 198407. [Google Scholar] [CrossRef] [PubMed]
- Zientara, S.; Weyer, C.T.; Lecollinet, S. African Horse Sickness. Rev. Sci. Tech. 2015, 34, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Zwart, L.; Potgieter, C.A.; Clift, S.J.; van Staden, V. Characterising Non-Structural Protein NS4 of African Horse Sickness Virus. PLoS ONE 2015, 10, e0124281. [Google Scholar] [CrossRef] [PubMed]
- Ngoveni, H.G.; van Schalkwyk, A.; Koekemoer, J.J.O. Evidence of Intragenic Recombination in African Horse Sickness Virus. Viruses 2019, 11, 654. [Google Scholar] [CrossRef]
- Boinas, F.; Calistrib, P.; Domingo, M.; Avilés, M.M.; López, B.M.; Sánchez, B.R.; Sánchez-Vizcaíno, J.M. Scientific Report Submitted to EFSA: Scientific Review on African Horse Sickness. EFSA Support. Publ. 2009, 6, 1–61. [Google Scholar]
- Gilkerson, J.R.; Bailey, K.E.; Diaz-Méndez, A.; Hartley, C.A. Update on Viral Diseases of the Equine Respiratory Tract. Vet. Clin. N. Am. Equine Pract. 2015, 31, 91–104. [Google Scholar] [CrossRef]
- Mellor, P.S.; Hamblin, C. African Horse Sickness. Vet. Res. 2004, 35, 445–466. [Google Scholar] [CrossRef] [PubMed]
- Van Vuuren, M.; Penzhorn, B.L. Geographic Range of Vector-Borne Infections and Their Vectors: The Role of African Wildlife. Rev. Sci. Tech. 2015, 34, 139–149. [Google Scholar] [CrossRef]
- Robin, M.; Page, P.; Archer, D.; Baylis, M. African Horse Sickness: The Potential for an Outbreak in Disease-Free Regions and Current Disease Control and Elimination Techniques. Equine Vet. J. 2016, 48, 659–669. [Google Scholar] [CrossRef]
- Becker, E.; Venter, G.J.; Greyling, T.; Molini, U.; van Hamburg, H. Evidence of African Horse Sickness Virus Infection of Equus Zebra Hartmannae in the South-Western Khomas Region, Namibia. Transbound. Emerg. Dis. 2018, 65, 278–280. [Google Scholar] [CrossRef]
- Porphyre, T.; Grewar, J.D. Assessing the Potential of Plains Zebra to Maintain African Horse Sickness in the Western Cape Province, South Africa. PLoS ONE 2019, 14, 1–24. [Google Scholar] [CrossRef]
- Castillo-Olivares, J. African Horse Sickness in Thailand: Challenges of Controlling an Outbreak by Vaccination. Equine. Vet. J. 2021, 53, 9–14. [Google Scholar] [CrossRef]
- King, S.; Rajko-Nenow, P.; Ashby, M.; Frost, L.; Carpenter, S.; Batten, C. Outbreak of African Horse Sickness in Thailand, 2020. Transbound. Emerg. Dis. 2020, 67, 1764–1767. [Google Scholar] [CrossRef]
- Lu, G.; Pan, J.; Ou, J.; Shao, R.; Hu, X.; Wang, C.; Li, S. African Horse Sickness: Its Emergence in Thailand and Potential Threat to Other Asian Countries. Transbound. Emerg. Dis. 2020, 67, 1751–1753. [Google Scholar] [CrossRef]
- Mellor, P.S.; Boned, J.; Hamblin, C.; Graham, S. Isolations of African Horse Sickness Virus from Vector Insects Made during the 1988 Epizootic in Spain. Epidemiol. Infect. 1990, 105, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.; Hooghuis, H.; Castaño, M. African Horse Sickness in Spain. Vet. Microbiol. 1992, 33, 129–142. [Google Scholar] [CrossRef]
- Thompson, G.M.; Jess, S.; Murchie, A.K. A Review of African Horse Sickness and Its Implications for Ireland. Ir. Vet. J. 2012, 65, 9. [Google Scholar] [CrossRef] [PubMed]
- Guichard, S.; Guis, H.; Tran, A.; Garros, C.; Balenghien, T.; Kriticos, D.J. Worldwide niche and future potential distribution of Culicoides imicola, a major vector of bluetongue and African horse sickness viruses. PLoS ONE 2014, 9, e0119323. [Google Scholar] [CrossRef]
- Martinez-de la Puente, J.; Navarro, J.; Ferraguti, M.; Soriguer, R.; Figuerola, J. First molecular identification of the vertebrate hosts of Culicoides imicola in Europe and a review of its blood-feeding patterns worldwide: Implications for the transmission of bluetongue disease and African horse sickness. Med. Vet. Entomol. 2017, 31, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.S.; Alvarez, J.; Bicout, D.J.; Calistri, P.; Depner, K.; Drewe, J.A.; Garin-Bastuji, B.; Rojas, J.L.G.; Schmidt, C.G.; Herskin, M.; et al. Scientific Opinion on the Assessment of the Control Measures of the Category A Diseases of Animal Health Law: African Horse Sickness. EFSA J. 2021, 19, e06403. [Google Scholar]
- Toh, X.; Wang, Y.; Rajapakse, M.P.; Lee, B.; Songkasupa, T.; Suwankitwat, N.; Kamlangdee, A.; Fernandez, C.J.; Huangfu, T. Use of Nanopore Sequencing to Characterize African Horse Sickness Virus (AHSV) from the African Horse Sickness Outbreak in Thailand in 2020. Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Faber, E.; Tshilwane, S.I.; Kleef, M.V.; Pretorius, A. Virulent African Horse Sickness Virus Serotype 4 Interferes with the Innate Immune Response in Horse Peripheral Blood Mononuclear Cells In Vitro. Infect. Genet. Evol. 2021, 91, 104836. [Google Scholar] [CrossRef]
- OIE. Application for the official recognition by the OIE of free status for African horse sickness. In Terrestrial Animal Health Code; OIE: Paris, France, 2019. [Google Scholar]
- Redmond, E.F.; Jones, D.; Rushton, J.J.E.V.J. Economic Assessment of African Horse Sickness Vaccine Impact. Equine Vet. J. 2021. [Google Scholar] [CrossRef]
- Weyer, C.T.; Grewar, J.D.; Burger, P.; Rossouw, E.; Lourens, C.; Joone, C.; le Grange, M.; Coetzee, P.; Venter, E.; Martin, D.P.; et al. African Horse Sickness Caused by Genome Reassortment and Reversion to Virulence of Live, Attenuated Vaccine Viruses, South. Africa, 2004–2014. Emerg. Infect. Dis. 2016, 22, 2087–2096. [Google Scholar]
- Wernery, U.; Joseph, S.; Raghavan, R.; Dyer, B.; Spendrup, S. African Horse Sickness Fever in Vaccinated Horses: Short Communication. J. Equine Vet. Sci. 2020, 88, 102967. [Google Scholar] [CrossRef]
- Grewar, J.D.; Sergeant, E.S.; Weyer, C.T.; van Helden, L.S.; Parker, B.J.; Anthony, T.; Thompson, P.N. Establishing Post-Outbreak Freedom from African Horse Sickness Virus in South Africa’s Surveillance Zone. Transbound. Emerg. Dis. 2019, 66, 2288–2296. [Google Scholar] [CrossRef]
- Wernery, U.; Rodriguez, M.; Raghavan, R.; Syriac, G.; Miriam Thomas, S.; Elizabeth, S.K.; Federico Ronchi, G.; Muhammed, R.; Patteril, N.A.; Joseph, S. Humoral Antibody Response of 10 Horses after Vaccination against African Horse Sickness with an Inactivated Vaccine Containing All 9 Serotypes in One Injection. Equine. Vet. J. 2021, 53, 826–833. [Google Scholar] [CrossRef]
- Lelli, R.; Molini, U.; Ronchi, G.F.; Rossi, E.; Franchi, P.; Ulisse, S.; Armillotta, G.; Capista, S.; Khaiseb, S.; di Ventura, M. Inactivated and Adjuvanted Vaccine for the Control of the African Horse Sickness Virus Serotype 9 Infection: Evaluation of Efficacy in Horses and Guinea-Pig Model. Vet. Ital. 2013, 49, 89–98. [Google Scholar]
- Dennis, S.J.; O’Kennedy, M.M.; Rutkowska, D.; Tsekoa, T.; Lourens, C.W.; Hitzeroth, I.; Meyers, A.E.; Rybicki, E.P. Safety and Immunogenicity of Plant-Produced African Horse Sickness Virus-Like Particles in Horses. Vet. Res. 2018, 49, 105. [Google Scholar]
- Rodríguez, M.; Joseph, S.; Pfeffer, M.; Raghavan, R.; Wernery, U. Immune Response of Horses to Inactivated African Horse Sickness Vaccines. BMC Vet. Res. 2020, 16, 322. [Google Scholar] [CrossRef]
- Calvo-Pinilla, E.; Gubbins, S.; Mertens, P.; Ortego, J.; Castillo-Olivares, J. The Immunogenicity of Recombinant Vaccines Based on Modified Vaccinia Ankara (MVA) Viruses Expressing African Horse Sickness Virus VP2 Antigens Depends on the Levels of Expressed VP2 Protein Delivered to the Host. Antivir. Res. 2018, 154, 132–139. [Google Scholar] [CrossRef]
- Calvo-Pinilla, E.; Marín-López, A.; Utrilla-Trigo, S.; Jiménez-Cabello, L.; Ortego, J. Reverse Genetics Approaches: A Novel Strategy for African Horse Sickness Virus Vaccine Design. Curr. Opin. Virol. 2020, 44, 49–56. [Google Scholar] [CrossRef]
- O’Hara, R.; Meyer, A.; Burroughs, J.; Pullen, L.; Martin, L.-A.; Mertens, P. Development of a Mouse Model System, Coding Assignments and Identification of the Genome Segments Controlling Virulence of African Horse Sickness Virus Serotypes 3 and 8. In African Horse Sickness; Springer: Vienna, Austria, 1998; pp. 259–279. [Google Scholar]
- Aksular, M.; Calvo-Pinilla, E.; Marín-López, A.; Ortego, J.; Chambers, A.C.; King, L.A.; Castillo-Olivares, J. A Single Dose of African Horse Sickness Virus (AHSV) VP2 Based Vaccines Provides Complete Clinical Protection in a Mouse Model. Vaccine 2018, 36, 7003–7010. [Google Scholar] [CrossRef]
- Castillo-Olivares, J.; Calvo-Pinilla, E.; Casanova, I.; Bachanek-Bankowska, K.; Chiam, R.; Maan, S.; Nieto, J.M.; Ortego, J.; Mertens, P.P. A Modified Vaccinia Ankara Virus (MVA) Vaccine Expressing African Horse Sickness Virus (AHSV) VP2 Protects against AHSV Challenge in an IFNAR −/− Mouse Model. PLoS ONE 2011, 6, e16503. [Google Scholar] [CrossRef]
- Jones, L.; Hawes, P.; Salguero, J.; Castillo-Olivares, J.J.A.M. African Horse Sickness Virus: Pathogenicity in an IFNAR (−/−) Mouse Model of Infection. Access Microbiol. 2019, 1, 15. [Google Scholar] [CrossRef]
- Ronchi, G.F.; Ulisse, S.; Rossi, E.; Franchi, P.; Armillotta, G.; Capista, S.; Peccio, A.; di Ventura, M.; Pini, A. Immunogenicity of Two Adjuvant Formulations of an Inactivated African Horse Sickness Vaccine in Guinea-Pigs and Target Animals. Vet. Ital. 2012, 48, 67–76. [Google Scholar]
- Salas, M.L.; Andrés, G.J.V.R. African Swine Fever Virus Morphogenesis. Virus Res. 2013, 173, 29–41. [Google Scholar] [CrossRef]
- Liu, S.; Luo, Y.; Wang, Y.; Li, S.; Zhao, Z.; Bi, Y.; Sun, J.; Peng, R.; Song, H.; Zhu, D.; et al. Cryo-EM Structure of the African Swine Fever Virus. Cell Host Microbe 2019, 26, 836–843.e3. [Google Scholar] [CrossRef]
- Galindo, I.; Alonso, C.J.V. African Swine Fever Virus: A Review. Viruses 2017, 9, 103. [Google Scholar] [CrossRef]
- Dixon, L.K.; Stahl, K.; Jori, F.; Vial, L.; Pfeiffer, D.U. African Swine Fever Epidemiology and Control. Annu. Rev. Anim. Biosci. 2020, 8, 221–246. [Google Scholar] [CrossRef]
- Karger, A.; Pérez-Núñez, D.; Urquiza, J.; Hinojar, P.; Alonso, C.; Freitas, F.B.; Revilla, Y.; le Potier, M.-F.; Montoya, M.J.V. An Update on African Swine Fever Virology. Viruses 2019, 11, 864. [Google Scholar] [CrossRef]
- Alejo, A.; Matamoros, T.; Guerra, M.; Andrés, G.J.J.O.V. A Proteomic Atlas of the African Swine Fever Virus Particle. J. Virol. 2018, 92, e01293-18. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Vizcaíno, J.M.; Martínez-López, B.; Martínez-Avilés, M.; Martins, C.; Boinas, F.; Vialc, L.; Michaud, V.; Jori, F.; Etter, E.; Albina, E. Scientific Review on African Swine Fever. EFSA Support. Publ. 2009, 6, 1–141. [Google Scholar] [CrossRef]
- Arias, M.; Jurado, C.; Gallardo, C.; Fernández-Pinero, J.; Sánchez, J.J.T.; Vizcaíno, M.S. Gaps in African Swine Fever: Analysis and Priorities. Transbound. Emerg. Dis. 2018, 65, 235–247. [Google Scholar] [CrossRef]
- Costard, S.; Wieland, B.; de Glanville, W.; Jori, F.; Rowlands, R.; Vosloo, W.; Roger, F.; Pfeiffer, D.U.; Dixon, L.K. African Swine Fever: How Can Global Spread Be Prevented? Philos. Trans. R. Soc. B 2009, 364, 2683–2696. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.S.; Suresh, K.P.; Vashist, V.; Prajapati, A.; Pattnaik, B.; Roy, P.J.V.W. African Swine Fever: A Permanent Threat to Indian Pigs. Vet. World 2020, 13, 2275–2285. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Liu, T.; Liu, Y.; Xiao, J.; Wang, H. Transmission of African Swine Fever in China through Legal Trade of Live Pigs. Transbound. Emerg. Dis. 2021, 68, 355–360. [Google Scholar] [CrossRef]
- Ma, J.; Chen, H.; Gao, X.; Xiao, J.; Wang, H. African Swine Fever Emerging in China: Distribution Characteristics and High-Risk Areas. Prev. Vet. Med. 2020, 175, 1–5. [Google Scholar] [CrossRef]
- Zhou, X.; Li, N.; Luo, Y.; Liu, Y.; Miao, F.; Chen, T.; Zhang, S.; Cao, P.; Li, X.; Tian, K. Emergence of African Swine Fever in China, 2018. Transbound. Emerg. Dis. 2018, 65, 1482–1484. [Google Scholar] [CrossRef]
- Wang, N.; Zhao, D.; Wang, J.; Zhang, Y.; Wang, M.; Gao, Y.; Li, F.; Wang, J.; Bu, Z.; Rao, Z. Architecture of African Swine Fever Virus and Implications for Viral Assembly. Science 2019, 366, 640–644. [Google Scholar] [CrossRef]
- Blome, S.; Franzke, K.; Beer, M. African Swine Fever—A Review of Current Knowledge. Virus Res. 2020, 287, 198099. [Google Scholar] [CrossRef]
- Wu, K.; Liu, J.; Wang, L.; Fan, S.; Li, Z.; Li, Y.; Yi, L.; Ding, H.; Zhao, M.; Chen, J. Current State of Global African Swine Fever Vaccine Development under the Prevalence and Transmission of ASF in China. Vaccines. 2020, 8, 531. [Google Scholar] [CrossRef]
- OIE. African swine fever (infection with african swince fever). In Terrestrial Animal Health Manual; OIE: Paris, France, 2019. [Google Scholar]
- Penrith, M.-L.; Vosloo, W. Review of African Swine Fever: Transmission, Spread and Control. J. S. Afr. Vet. Assoc. 2009, 80, 58–62. [Google Scholar]
- Sánchez-Cordón, P.; Montoya, M.; Reis, A.; Dixon, L. African Swine Fever: A Re-Emerging Viral Disease Threatening the Global Pig Industry. Vet. J. 2018, 233, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Hervás, J.; Gómez-Villamandos, J.; Méndez, A.; Carrasco, L.; Sierra, M. The Lesional Changes amd Pathogenesis in the Kidney in African Swine Fever. Vet. Res. Commun. 1996, 20, 285–299. [Google Scholar] [CrossRef]
- Revilla, Y.; Perez-Nunez, D.; Richt, J.A. African Swine Fever Virus Biology and Vaccine Approaches. Adv. Virus Res. 2018, 100, 41–74. [Google Scholar]
- Gaudreault, N.N.; Madden, D.W.; Wilson, W.C.; Trujillo, J.D.; Richt, J.A. African Swine Fever Virus: An. Emerging DNA Arbovirus. Front. Vet. Sci. 2020, 7, 1–17. [Google Scholar]
- Gaudreault, N.N.; Richt, J.A. Subunit Vaccine Approaches for African Swine Fever Virus. Vaccines 2019, 7, 56. [Google Scholar] [CrossRef]
- Dixon, L.K.; DChapman, A.; Netherton, C.L.; Upton, C. African Swine Fever Virus Replication and Genomics. Virus Res. 2013, 173, 3–14. [Google Scholar] [CrossRef]
- Dixon, L.; Islam, M.; Nash, R.; Reis, A. African Swine Fever Virus Evasion of Host Defences. Virus Res. 2019, 266, 25–33. [Google Scholar] [CrossRef]
- Razzuoli, E.; Franzoni, G.; Carta, T.; Zinellu, S.; Amadori, M.; Modesto, P.; Oggiano, A. Modulation of Type I Interferon System by African Swine Fever Virus. Pathogens 2020, 9, 361. [Google Scholar] [CrossRef]
- Zhuo, Y.; Guo, Z.; Ba, T.; Zhang, C.; He, L.; Zeng, C.; Dai, H. African Swine Fever Virus MGF360–12L Inhibits Type I Interferon Production by Blocking the Interaction of Importin A and NF-Κb Signaling Pathway. Virol. Sin. 2021, 36, 176–186. [Google Scholar] [CrossRef]
- Sánchez, E.G.; Pérez-Núñez, D.; Revilla, Y. Development of Vaccines against African Swine Fever Virus. Virus Res. 2019, 265, 150–155. [Google Scholar] [CrossRef]
- Teklue, T.; Sun, Y.; Abid, M.; Luo, Y.; Qiu, H.J. Current Status and Evolving Approaches to African Swine Fever Vaccine Development. Transbound. Emerg. Dis. 2020, 67, 529–542. [Google Scholar] [CrossRef]
- Bosch-Camós, L.; López, E.; Rodriguez, F. African Swine Fever Vaccines: A Promising Work Still in Progress. Porc. Health Manag. 2020, 6, 1–14. [Google Scholar] [CrossRef]
- Sang, H.; Miller, G.; Lokhandwala, S.; Sangewar, N.; Waghela, S.D.; Bishop, R.P.; Mwangi, W. Progress toward Development of Effective and Safe African Swine Fever Virus Vaccines. Front. Vet. Sci. 2020, 7, 1–9. [Google Scholar] [CrossRef]
- Rock, D. Challenges for African Swine Fever Vaccine Development—“…Perhaps the End of the Beginning”. Vet. Microbiol. 2017, 206, 52–58. [Google Scholar] [CrossRef]
- Carlson, J.; O’Donnell, V.; Alfano, M.; Salinas, L.V.; Holinka, L.G.; Krug, P.W.; Gladue, D.P.; Higgs, S.; Borca, M.V. Association of the Host Immune Response with Protection Using a Live Attenuated African Swine Fever Virus Model. Viruses 2016, 8, 291. [Google Scholar] [CrossRef]
- Neilan, J.G.; Zsak, L.; Lu, Z.; Burrage, T.G.; Kutish, G.F.; Rock, D.L. Neutralizing Antibodies to African Swine Fever Virus Proteins P30, P54, and P72 Are Not Sufficient for Antibody-Mediated Protection. Virology 2004, 319, 337–342. [Google Scholar] [CrossRef]
- Schulz, K.; Staubach, C.; Blome, S. African and Classical Swine Fever: Similarities, Differences and Epidemiological Consequences. Vet. Res. 2017, 48, 84. [Google Scholar] [CrossRef]
- Yoo, D.; Kim, H.; Lee, J.Y.; Yoo, H.S. African Swine Fever: Etiology, Epidemiological Status in Korea, and Perspective on Control. J. Vet. Sci. 2020, 21, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Dixon, L.; Sun, H.; Roberts, H. African Swine Fever. Antivir. Res. 2019, 165, 34–41. [Google Scholar] [CrossRef]
- Arias, M.; Sánchez-Vizcaíno, J.; Morilla, A.; Yoon, K.; Zimmerman, J. Trends in Emerging Viral Infections of Swine; Iowa State Press: Ames, IA, USA, 2002. [Google Scholar]
- Mur, L.; Atzeni, M.; Martínez-López, B.; Feliziani, F.; Rolesu, S.; Sanchez-Vizcaino, J. Thirty-Five-Year Presence of African Swine Fever in Sardinia: History, Evolution and Risk Factors for Disease Maintenance. Transbound Emerg. Dis. 2016, 63, e165–e177. [Google Scholar] [CrossRef]
- Mur, L.; Sánchez-Vizcaíno, J.; Fernández-Carrión, E.; Jurado, C.; Rolesu, S.; Feliziani, F.; Laddomada, A.; Martínez-López, B. Understanding African Swine Fever Infection Dynamics in Sardinia Using a Spatially Explicit Transmission Model in Domestic Pig Farms. Transbound Emerg. Dis. 2018, 65, 123–134. [Google Scholar] [CrossRef]
- Swayne, D.E. Avian Influenza; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Swayne, D.E. Understanding the Complex Pathobiology of High Pathogenicity Avian Influenza Viruses in Birds. Avian. Dis. 2007, 51 (Suppl. S1), 242–249. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.J. A Review of Avian Influenza in Different Bird Species. Vet. Microbiol. 2000, 74, 3–13. [Google Scholar] [CrossRef]
- Lee, D.H.; Bertran, K.; Kwon, J.H.; Swayne, D.E. Evolution, Global Spread, and Pathogenicity of Highly Pathogenic Avian Influenza H5Nx Clade 2.3.4.4. J. Vet. Sci. 2017, 18 (Suppl. S1), 269–280. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Criado, M.F.; Swayne, D.E. Pathobiological Origins and Evolutionary History of Highly Pathogenic Avian Influenza Viruses. Cold Spring Harb. Perspect. Med. 2021, 11, 1–22. [Google Scholar] [CrossRef]
- Peacock, T.H.P.; James, J.; Sealy, J.E.; Iqbal, M. A Global Perspective on H9N2 Avian Influenza Virus. Viruses 2019, 11, 620. [Google Scholar] [CrossRef]
- Spickler, A.R. Highly Pathogenic Avian Influenza; 2016; pp. 1–38. Available online: http://www.cfsph.iastate.edu/DiseaseInfo/factsheets.php (accessed on 16 May 2021).
- Shriner, S.A.; Root, J.J. A Review of Avian Influenza A Virus Associations in Synanthropic Birds. Viruses 2020, 12, 1209. [Google Scholar] [CrossRef]
- Root, J.; Shriner, S. Avian Influenza A Virus Associations in Wild, Terrestrial Mammals: A Review of Potential Synanthropic Vectors to Poultry Facilities. Viruses 2020, 12, 1352. [Google Scholar] [CrossRef]
- Gorsich, E.E.; Webb, C.T.; Merton, A.A.; Hoeting, J.A.; Miller, R.S.; Farnsworth, M.L.; Swafford, S.R.; DeLiberto, T.J.; Pedersen, K.; Franklin, A.B.; et al. Continental-Scale Dynamics of Avian Influenza in U.S. Waterfowl Are Driven by Demography, Migration, and Temperature. Ecol. Appl. 2021, 31, e2245. [Google Scholar]
- Khan, M.; Chaudhry, M.; Fatima, Z.; Khan, R.U.; Ahmad, B.; Ullah, R.; Khan, A. Effect of Avian Influenza H9N2 Subtype Virus Infection on Backyard Poultry Production. Sci. Lett. 2021, 9, 19–23. [Google Scholar]
- Gu, M.; Xu, L.; Wang, X.; Liu, X. Current Situation of H9N2 Subtype Avian Influenza in China. Vet. Res. 2017, 48, 49. [Google Scholar] [CrossRef]
- Kim, Y.; Biswas, P.K.; Giasuddin, M.; Hasan, M.; Mahmud, R.; Chang, Y.M.; Essen, S.; Samad, M.A.; Lewis, N.S.; Brown, I.H.; et al. Prevalence of Avian Influenza A(H5) and A(H9) Viruses in Live Bird Markets, Bangladesh. Emerg. Infect. Dis. 2018, 24, 2309–2316. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Chen, Q.; Wang, Q.; Chen, J.; Jin, T.; Wong, G.; Quan, C.; Liu, J.; Wu, J.; Yin, R.; et al. Genesis, Evolution and Prevalence of H5N6 Avian Influenza Viruses in China. Cell Host Microb. 2016, 20, 810–821. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Zhou, S.; Dong, L.; van Boeckel, T.P.; Cui, Y.; Newman, S.H.; Takekawa, J.Y.; Prosser, D.J.; Xiao, X.; Wu, Y.; et al. Avian Influenza H5N1 Viral and Bird Migration Networks in Asia. Proc. Natl. Acad. Sci. USA 2015, 112, 172–177. [Google Scholar] [CrossRef]
- Nuñez, I.A.; Ross, T.M. A Review of H5Nx Avian Influenza Viruses. Adv. Vaccines Immunother. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- OIE. Highly Pathogenic Avian Influenza (HPAI) Report N° 21: January 15 to February 04, 2021; OIE: Paris, France, 2021. [Google Scholar]
- Verhagen, J.H.; Fouchier, R.A.M.; Lewis, N. Highly Pathogenic Avian Influenza Viruses at the Wild-Domestic Bird Interface in Europe: Future Directions for Research and Surveillance. Viruses 2021, 13, 212. [Google Scholar] [CrossRef]
- Suarez, D.L.; Das, A.; Ellis, E. Review of Rapid Molecular Diagnostic Tools for Avian Influenza Virus. Avian Dis. 2007, 51, 201–208. [Google Scholar] [CrossRef]
- Beerens, N.; Koch, G.; Heutink, R.; Harders, F.; Vries, D.E.; Ho, C.; Bossers, A.; Elbers, A. Novel Highly Pathogenic Avian Influenza A (H5N6) Virus in the Netherlands, December 2017. Emerg. Infect. Dis. 2018, 24, 770–773. [Google Scholar] [CrossRef]
- OIE. Terrestrial Animal Health Manual; OIE: Paris, France, 2018. [Google Scholar]
- Liu, S.; Zhuang, Q.; Wang, S.; Jiang, W.; Jin, J.; Peng, C.; Hou, G.; Li, J.; Yu, J.; Yu, X.; et al. Control. of Avian Influenza in China: Strategies and Lessons. Transbound. Emerg. Dis. 2020, 67, 1463–1471. [Google Scholar] [CrossRef]
- Gilbert, M.; Golding, N.; Zhou, H.; Wint, G.W.; Robinson, T.P.; Tatem, A.J.; Lai, S.; Zhou, S.; Jiang, H.; Guo, D. Predicting the Risk of Avian Influenza a H7N9 Infection in Live-Poultry Markets across Asia. Nat. Commun. 2014, 5, 1–7. [Google Scholar] [CrossRef]
- St Charles, K.M.; Ssematimba, A.; Malladi, S.; Bonney, P.J.; Linskens, E.; Culhane, M.; Goldsmith, T.J.; Halvorson, D.A.; Cardona, C.J. Avian Influenza in the U.S. Commercial Upland Game Bird Industry: An Analysis of Selected Practices as Potential Exposure Pathways and Surveillance System Data Reporting. Avian Dis. 2018, 62, 307–315. [Google Scholar]
- Capua, I.; Marangon, S. Control. of Avian Influenza in Poultry. Emerg. Infect. Dis. 2006, 12, 1319–1324. [Google Scholar] [CrossRef]
- Parvin, R.; Nooruzzaman, M.; Kabiraj, C.K.; Begum, J.A.; Chowdhury, E.H.; Islam, M.R.; Harder, T. Controlling Avian Influenza Virus in Bangladesh: Challenges and Recommendations. Viruses 2020, 12, 751. [Google Scholar] [CrossRef] [PubMed]
- OIE. Avian Influenza; OIE: Paris, France, 2021. [Google Scholar]
- Peyre, M.; Fusheng, G.; Desvaux, S.; Roger, F. Avian Influenza Vaccines: A Practical Review in Relation to Their Application in the Field with a Focus on the Asian Experience. Epidemiol. Infect. 2009, 137, 1–21. [Google Scholar] [CrossRef]
- Capua, I.; Marangon, S. The Avian Influenza Epidemic in Italy, 1999–2000: A review. Avian Pathol. 2000, 29, 289–294. [Google Scholar] [CrossRef]
- Suarez, D.L. Overview of Avian Influenza DIVA Test Strategies. Biologicals 2005, 33, 221–226. [Google Scholar] [CrossRef]
- Hasan, N.H.; Ignjatovic, J.; Peaston, A.; Hemmatzadeh, F. Avian Influenza Virus and DIVA Strategies. Viral. Immunol. 2016, 29, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Samal, S.K. Innovation in Newcastle Disease Virus Vectored Avian Influenza Vaccines. Viruses 2019, 11, 300. [Google Scholar] [CrossRef] [PubMed]
- Cardona, C.J.; Halvorson, D.A.; Hall, J.; Pantin-Jackwood, M.J.; Brown, J.D. Conducting Influenza Virus Pathogenesis Studies in Avian Species. Methods Mol. Biol. 2020, 2123, 195–209. [Google Scholar] [PubMed]
- Driskell, E.A.; Jones, C.A.; Stallknecht, D.E.; Howerth, E.W.; Tompkins, S.M. Avian Influenza Virus Isolates from Wild Birds Replicate and Cause Disease in a Mouse Model of Infection. Virology 2010, 399, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Kim, Y.I.; Pascua, P.N.; Choi, Y.K. Avian Influenza A Viruses: Evolution and Zoonotic Infection. Semin. Respir. Crit. Care Med. 2016, 37, 501–511. [Google Scholar] [CrossRef]
- Koo, B.S.; Kim, H.K.; Song, D.; Na, W.; Song, M.S.; Kwon, J.J.; Wong, S.S.; Noh, J.Y.; Ahn, M.J.; Kim, D.J.; et al. Virological and Pathological Characterization of an Avian H1N1 Influenza A Virus. Arch. Virol. 2018, 163, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Yuan, B.; Xia, X.; Zhang, S.; Du, Q.; Yang, C.; Li, N.; Zhao, J.; Zhang, Y.; Zhang, R.; et al. Tree Shrew as a New Animal Model to Study the Pathogenesis of Avian Influenza (H9N2) Virus Infection. Emerg. Microbes Infect. 2018, 7, 166. [Google Scholar] [CrossRef]
- Pulit-Penaloza, J.A.; Brock, N.; Pappas, C.; Sun, X.; Belser, J.A.; Zeng, H.; Tumpey, T.M.; Maines, T.R. Characterization of Highly Pathogenic Avian Influenza H5Nx Viruses in the Ferret Model. Sci. Rep. 2020, 10, 12700. [Google Scholar] [CrossRef]
- Veldhuis Kroeze, E.; Bauer, L.; Caliendo, V.; van Riel, D. In Vivo Models to Study the Pathogenesis of Extra-Respiratory Complications of Influenza A Virus Infection. Viruses 2021, 13, 848. [Google Scholar] [CrossRef]
- Zanin, M.; Koçer, Z.A.; Poulson, R.L.; Gabbard, J.D.; Howerth, E.W.; Jones, C.A.; Friedman, K.; Seiler, J.; Danner, A.; Kercher, L.; et al. Potential for Low-Pathogenic Avian H7 Influenza A Viruses to Replicate and Cause Disease in a Mammalian Model. J. Virol. 2017, 91, 1–19. [Google Scholar] [CrossRef]
- Lei, H.; Lu, X.; Li, S.; Ren, Y. High Immune Efficacy against Different Avian Influenza H5N1 Viruses Due to Oral Administration of a Saccharomyces Cerevisiae-Based Vaccine in Chickens. Sci. Rep. 2021, 11, 8977. [Google Scholar] [CrossRef] [PubMed]
- Germeraad, E.A.; Sanders, P.; Hagenaars, T.J.; Jong, M.C.M.; Beerens, N.; Gonzales, J.L. Virus Shedding of Avian Influenza in Poultry: A Systematic Review and Meta-Analysis. Viruses 2019, 11, 812. [Google Scholar] [CrossRef]
- Mei, K.; Guo, Y.; Zhu, X.; Qu, N.; Huang, J.; Chen, Z.; Zhang, Y.; Zhao, B.; He, Z.; Liao, M.; et al. Different Pathogenicity and Transmissibility of Goose-Origin H5N6 Avian Influenza Viruses in Chickens. Viruses 2019, 11, 612. [Google Scholar] [CrossRef]
- Cardona, C.J.; Halvorson, D.A.; Hall, J.; Pantin-Jackwood, M.J.; Brown, J.D. Conducting Influenza Virus Pathogenesis Studies in Avian Species, in Animal Influenza Virus; Springer: New York, NY, USA, 2020; pp. 195–209. [Google Scholar]
- Roy, P.; Noad, R. Bluetongue Virus Assembly and Morphogenesis. In Reoviruses: Entry, Assembly and Morphogenesis; Roy, P., Ed.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 87–116. [Google Scholar]
- Schwartz-Cornil, I.; Mertens, P.; Contreras, V.; Hemati, B.; Pascale, F.; Bréard, E.; Mellor, P.; MacLachlan, N.; Zientara, S. Bluetongue Virus: Virology, Pathogenesis and Immunity. Vet. Res. 2008, 39, 46. [Google Scholar] [CrossRef] [PubMed]
- Prasad, B.V.; Yamaguchi, S.; Roy, P. Three-Dimensional Structure of Single-Shelled Bluetongue Virus. J. Virol. 1992, 66, 2135–2142. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.E.; Turner, J.; Caminade, C.; Heath, A.E.; Wardeh, M.; Kluiters, G.; Diggle, P.J.; Morse, A.P.; Baylis, M. Bluetongue Risk under Future Climates. Nat. Clim. Chang. 2019, 9, 153–157. [Google Scholar] [CrossRef]
- Rao, P.P.; Hegde, N.R.; Singh, K.P.; Putty, K.; Hemadri, D.; Maan, N.S.; Reddy, Y.N.; Maan, S.; Mertens, P.P.C. Bluetongue: Aetiology, Epidemiology, Pathogenesis, Diagnosis and Control., in Emerging and Re-Emerging Infectious Diseases of Livestock; Bayry, J., Ed.; Springer International Publishing: Cham, Switzerland, 2007; pp. 3–54. [Google Scholar]
- Walton, T.E. The History of Bluetongue and a Current Global Overview. Vet. Ital. 2004, 40, 31–38. [Google Scholar] [PubMed]
- Maclachlan, N.J.; Guthrie, A.J. Re-Emergence of Bluetongue, African Horse Sickness, and Other Orbivirus Diseases. Vet. Res. 2010, 41, 35. [Google Scholar] [CrossRef] [PubMed]
- Ries, C.; Sharav, T.; Tseren-Ochir, E.-O.; Beer, M.; Hoffmann, B. Putative Novel Serotypes ‘33’ and ‘35’ in Clinically Healthy Small Ruminants in Mongolia Expand the Group of Atypical BTV. Viruses 2021, 13, 42. [Google Scholar] [CrossRef]
- Bumbarov, V.; Golender, N.; Jenckel, M.; Wernike, K.; Beer, M.; Khinich, E.; Zalesky, O.; Erster, O. Characterization of Bluetongue Virus Serotype 28. Transbound. Emerg. Dis. 2020, 67, 171–182. [Google Scholar] [CrossRef]
- Marcacci, M.; Sant, S.; Mangone, I.; Goria, M.; Dondo, A.; Zoppi, S.; van Gennip, R.G.P.; Radaelli, M.C.; Cammà, C.; van Rijn, P.A.; et al. One after the Other: A Novel Bluetongue Virus Strain Related to Toggenburg Virus Detected in the Piedmont Region (North-Western Italy), Extends the Panel of Novel Atypical BTV Strains. Transbound. Emerg. Dis. 2018, 65, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Ries, C.; Domes, U.; Janowetz, B.; Böttcher, J.; Burkhardt, K.; Miller, T.; Beer, M.; Hoffmann, B. Isolation and Cultivation of a New Isolate of BTV-25 and Presumptive Evidence for a Potential Persistent Infection in Healthy Goats. Viruses 2020, 12, 983. [Google Scholar] [CrossRef] [PubMed]
- Rupner, R.N.; Vinodh Kumar, R.O.; Karthikeyan, R.; Sinha, D.K.; Singh, K.P.; Dubal, Z.B.; Tamta, S.; Gupta, V.K.; Singh, B.R.; Malik, Y.S.; et al. Bluetongue in India: A Systematic Review and Meta-Analysis with Emphasis on Diagnosis and Seroprevalence. Vet. Q. 2020, 40, 229–242. [Google Scholar] [CrossRef]
- Parsonson, I.M. Pathology and Pathogenesis of Bluetongue Infections; Springer: Berlin/Heidelberg, Germany, 1990. [Google Scholar]
- Maclachlan, N.J.; Drew, C.P.; Darpel, K.E.; Worwa, G. The Pathology and Pathogenesis of Bluetongue. J. Comp. Pathol. 2009, 141, 1–16. [Google Scholar] [CrossRef]
- Darpel, K.E.; Batten, C.A.; Veronesi, E.; Shaw, A.E.; Anthony, S.; Bachanek-Bankowska, K.; Kgosana, L.; Bin-Tarif, A.; Carpenter, S.; Müller-Doblies, U.U.; et al. Clinical Signs and Pathology Shown by British Sheep and Cattle Infected with Bluetongue Virus Serotype 8 Derived from the 2006 Outbreak in Northern Europen. Vet. Rec. 2007, 161, 253–261. [Google Scholar] [CrossRef]
- Sperlova, A.; Zendulkova, D. Bluetongue: A Review. Vet. Med. 2011, 56, 430–452. [Google Scholar] [CrossRef]
- Linden, A.; Gregoire, F.; Nahayo, A.; Hanrez, D.; Mousset, B.; Massart, A.L.; de Leeuw, I.; Vandemeulebroucke, E.; Vandenbussche, F.; de Clercq, K. Bluetongue Virus in Wild Deer, Belgium, 2005–2008. Emerg. Infect. Dis. 2010, 16, 833–836. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Fons, F.; Sánchez-Matamoros, A.; Gortázar, C.; Sánchez-Vizcaíno, J.M. The Role of Wildlife in Bluetongue Virus Maintenance in Europe: Lessons Learned after the Natural Infection in Spain. Virus Res. 2014, 182, 50–58. [Google Scholar] [CrossRef]
- Wilbur, L.A.; Evermann, J.F.; Levings, R.L.; Stoll, I.R.; Starling, D.E.; Spillers, C.A.; Gustafson, G.A.; McKeirnan, A.J. Abortion and Death in Pregnant Bitches Associated with a Canine Vaccine Contaminated with Bluetongue Virus. J. Am. Vet. Med. Assoc. 1994, 204, 1762–1765. [Google Scholar] [PubMed]
- Alexander, K.A.; MacLachlan, N.J.; Kat, P.W.; House, C.; O’Brien, S.J.; Lerche, N.W.; Sawyer, M.; Frank, L.G.; Holekamp, K.; Smale, L.; et al. Evidence of Natural Bluetongue Virus Infection among African Carnivores. Am. J. Trop. Med. Hyg. 1994, 51, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Maclachlan, N.J. Global Implications of the Recent Emergence of Bluetongue Virus in Europe. Vet. Clin. N. Am. Food Anim. Pract. 2010, 26, 163–171. [Google Scholar] [CrossRef] [PubMed]
- MacLachlan, N.J.; Osburn, B.I. Impact of Bluetongue Virus Infection on the International Movement and Trade of Ruminants. J. Am. Vet. Med Assoc. 2006, 228, 1346–1349. [Google Scholar] [CrossRef]
- Velthuis, A.G.J.; Mourits, M.C.M.; Saatkamp, H.W.; de Koeijer, A.A.; Elbers, A.R.W. Financial Evaluation of Different Vaccination Strategies for Controlling the Bluetongue Virus Serotype 8 Epidemic in the Netherlands in 2008. PLoS ONE 2011, 6, e19612. [Google Scholar] [CrossRef]
- Pinior, B.; Lebl, K.; Firth, C.; Rubel, F.; Fuchs, R.; Stockreiter, S.; Loitsch, A.; Köfer, J. Cost Analysis of Bluetongue Virus Serotype 8 Surveillance and Vaccination Programmes in Austria from 2005 to 2013. Vet. J. 2015, 206, 154–160. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mayo, C.; Lee, J.; Kopanke, J.; MacLachlan, N.J. A Review of Potential Bluetongue Virus Vaccine Strategies. Vet. Microbiol. 2017, 206, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, P.; van Vuuren, M.; Venter, E.H.; Stokstad, M. A Review of Experimental Infections with Bluetongue Virus in the Mammalian Host. Virus Res. 2014, 182, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Martinelle, L.; Pozzo, F.D.; Thiry, E.; de Clercq, K.; Saegerman, C. Reliable and Standardized Animal Models to Study the Pathogenesis of Bluetongue and Schmallenberg Viruses in Ruminant Natural Host Species with Special Emphasis on Placental Crossing. Viruses 2019, 11, 753. [Google Scholar] [CrossRef] [PubMed]
- Maclachlan, N.J.; Crafford, J.E.; Vernau, W.; Gardner, I.A.; Goddard, A.; Guthrie, A.J.; Venter, E.H. Experimental Reproduction of Severe Bluetongue in Sheep. Vet. Pathol. 2008, 45, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Pini, A. Study on the Pathogenesis of Bluetongue: Replication of the Virus in the Organs of Infected Sheep. Onderstepoort J. Vet. Res. 1976, 43, 159–164. [Google Scholar]
- Gibbs, E.P.J.; Greiner, E.C. The Epidemiology of Bluetongue. Comp. Immunol. Microbiol. Infect. Dis. 1994, 17, 207–220. [Google Scholar] [CrossRef]
- Melville, L.F.; Hunt, N.T.; Davis, S.S.; Weir, R.P. Bluetongue Virus Does Not Persist in Naturally Infected Cattle. Vet. Ital. 2004, 40, 502–507. [Google Scholar]
- Dal Pozzo, F.; Saegerman, C.; Thiry, E. Bovine Infection with Bluetongue Virus with Special Emphasis on European Serotype 8. Vet. J. 2009, 182, 142–151. [Google Scholar] [CrossRef]
- Barratt-Boyes, S.M.; MacLachlan, N.J. Dynamics of Viral Spread in Bluetongue Virus Infected Calves. Vet. Microbiol. 1994, 40, 361–371. [Google Scholar] [CrossRef]
- Caporale, M.; Wash, R.; Pini, A.; Savini, G.; Franchi, P.; Golder, M.; Patterson-Kane, J.; Mertens, P.; di Gialleonardo, L.; Armillotta, G.; et al. Determinants of Bluetongue Virus Virulence in Murine Models of Disease. J. Virol. 2011, 85, 11479–11489. [Google Scholar] [CrossRef]
- Stokstad, M.; Coetzee, P.; Myrmel, M.; Mutowembwa, P.; Venter, E.H.; Larsen, S. Refined Experimental Design May Increase the Value of Murine Models for Estimation of Bluetongue Virus Virulence. Lab. Anim. 2021, 55, 53–64. [Google Scholar] [CrossRef]
- Narayan, O.; Johnson, R.T. Effects of Viral Infection on Nervous System Development. I. Pathogenesis of Bluetongue Virus Infection in Mice. Am. J. Pathol. 1972, 68, 1–14. [Google Scholar] [PubMed]
- Calvo-Pinilla, E.; Rodríguez-Calvo, T.; Anguita, J.; Sevilla, N.; Ortego, J. Establishment of a Bluetongue Virus Infection Model in Mice that Are Deficient in the Alpha/Beta Interferon Receptor. PLoS ONE 2009, 4, e5171. [Google Scholar]
- Ortego, J.; de la Poza, F.; Marín-López, A. Interferon α/β Receptor Knockout Mice as a Model to Study Bluetongue Virus Infection. Virus Res. 2014, 182, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Darpel, K.E.; Monaghan, P.; Simpson, J.; Anthony, S.J.; Veronesi, E.; Brooks, H.W.; Elliott, H.; Brownlie, J.; Takamatsu, H.-H.; Mellor, P.S.; et al. Involvement of the Skin During Bluetongue Virus Infection and Replication in the Ruminant Host. Vet. Res. 2012, 43, 40. [Google Scholar] [CrossRef]
- Umeshappa, C.S.; Singh, K.P.; Channappanavar, R.; Sharma, K.; Nanjundappa, R.H.; Saxena, M.; Singh, R.; Sharma, A.K. A Comparison of Intradermal and Intravenous Inoculation of Bluetongue Virus Serotype 23 in Sheep for Clinico-Pathology, and Viral and Immune Responses. Vet. Immunol. Immunopathol. 2011, 141, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Blome, S.; Staubach, C.; Henke, J.; Carlson, J.; Beer, M. Classical Swine Fever-An Updated Review. Viruses 2017, 9, 86. [Google Scholar] [CrossRef] [PubMed]
- Moennig, V.; Becher, P.; Beer, M. Classical Swine Fever. Dev. Biol. 2013, 135, 167–174. [Google Scholar]
- Moennig, V.; Floegel-Niesmann, G.; Greiser-Wilke, I. Clinical Signs and Epidemiology of Classical Swine Fever: A Review of New Knowledge. Vet. J. 2003, 165, 11–20. [Google Scholar] [CrossRef]
- Penrith, M.L.; Vosloo, W.; Mather, C. Classical Swine Fever (Hog Cholera): Review of Aspects Relevant to Control. Transbound. Emerg. Dis. 2011, 58, 187–196. [Google Scholar] [CrossRef]
- Summerfield, A.; Ruggli, N. Immune Responses Against Classical Swine Fever Virus: Between Ignorance and Lunacy. Front. Vet. Sci. 2015, 2, 10. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, J.; Yang, Q.; Anwar, M.N.; Yu, S.; Qiu, H.J. Complex Virus-Host Interactions Involved in the Regulation of Classical Swine Fever Virus Replication: A Minireview. Viruses 2017, 9, 86. [Google Scholar]
- Zhang, L.; Qin, Y.; Chen, M. Viral Strategies for Triggering and Manipulating Mitophagy. Autophagy 2018, 14, 1665–1673. [Google Scholar] [CrossRef] [PubMed]
- Zhu, E.; Chen, W.; Qin, Y.; Ma, S.; Fan, S.; Wu, K.; Li, W.; Fan, J.; Yi, L.; Ding, H.; et al. Classical Swine Fever Virus Infection Induces Endoplasmic Reticulum Stress-Mediated Autophagy to Sustain Viral Replication In Vivo and In Vitro. Front. Microbiol. 2019, 10, 2545. [Google Scholar] [PubMed]
- Leifer, I.; Ruggli, N.; Blome, S. Approaches to Define the Viral Genetic Basis of Classical Swine Fever Virus Virulence. Virology 2013, 438, 51–55. [Google Scholar] [CrossRef]
- Ma, S.M.; Mao, Q.; Yi, L.; Zhao, M.Q.; Chen, J.D. Apoptosis, Autophagy, and Pyroptosis: Immune Escape Strategies for Persistent Infection and Pathogenesis of Classical Swine Fever Virus. Pathogens 2019, 8, 239. [Google Scholar] [CrossRef] [PubMed]
- Vuono, E.A.; Ramirez-Medina, E.; Azzinaro, P.; Berggren, K.A.; Rai, A.; Pruitt, S.; Silva, E.; Velazquez-Salinas, L.; Borca, M.V.; Gladue, D.P. SERTA Domain Containing Protein 1 (SERTAD1) Interacts with Classical Swine Fever Virus Structural Glycoprotein E2, Which Is Involved in Virus Virulence in Swine. Viruses 2020, 12, 421. [Google Scholar] [CrossRef]
- Liu, C.C.; Liu, Y.Y.; Cheng, Y.; Zhang, Y.N.; Zhang, J.; Liang, X.D.; Gao, Y.; Chen, H.; Baloch, A.S.; Yang, Q.; et al. The ESCRT-I Subunit Tsg101 Plays Novel Dual Roles in Entry and Replication of Classical Swine Fever Virus. J. Virol. 2021, 95, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Dahle, J.; Liess, B. A Review on Classical Swine Fever Infections in Pigs: Epizootiology, Clinical Disease and Pathology. Comp. Immunol. Microbiol. Infect. Dis. 1992, 15, 203–211. [Google Scholar] [CrossRef]
- Fatima, M.; Luo, Y.; Zhang, L.; Wang, P.-Y.; Song, H.; Fu, Y.; Li, Y.; Sun, Y.; Li, S.; Bao, Y.-J. Genotyping and Molecular Characterization of Classical Swine Fever Virus Isolated in China during 2016–2018. Viruses 2021, 13, 664. [Google Scholar] [CrossRef]
- Stoian, A.M.M.; Petrovan, V.; Constance, L.A.; Olcha, M.; Dee, S.; Diel, D.G.; Sheahan, M.A.; Rowland, R.R.R.; Patterson, G.; Niederwerder, M.C. Stability of Classical Swine Fever Virus and Pseudorabies Virus in Animal Feed Ingredients Exposed to Transpacific Shipping Conditions. Transbound. Emerg. Dis. 2020, 67, 1623–1632. [Google Scholar] [CrossRef]
- Greiser-Wilke, I.; Blome, S.; Moennig, V. Diagnostic Methods for Detection of Classical Swine Fever Virus--Status Quo and New Developments. Vaccine 2007, 25, 5524–5530. [Google Scholar] [CrossRef] [PubMed]
- De Smit, A.J. Laboratory Diagnosis, Epizootiology, and Efficacy of Marker Vaccines in Classical Swine Fever: A Review. Vet. Q. 2000, 22, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Madera, R.; Li, Y.; McVey, D.S.; Drolet, B.S.; Shi, J. Recent Advances in the Diagnosis of Classical Swine Fever and Future Perspectives. Pathogens 2020, 9, 658. [Google Scholar] [CrossRef] [PubMed]
- Beemer, O.; Remmenga, M.; Gustafson, L.; Johnson, K.; Hsi, D.; Antognoli, M.C. Assessing the Value of PCR Assays in Oral Fluid Samples for Detecting African Swine Fever, Classical Swine Fever, and Foot-and-Mouth Disease in U.S. Swine. PLoS ONE 2019, 14, e0219532. [Google Scholar] [CrossRef] [PubMed]
- Chander, V.; Nandi, S.; Ravishankar, C.; Upmanyu, V.; Verma, R. Classical Swine Fever in Pigs: Recent Developments and Future Perspectives. Anim. Health Res. Rev. 2014, 15, 87–101. [Google Scholar] [CrossRef] [PubMed]
- OIE. Classical swine fever (infection with classical swine fever). In Terrestrial Animal Health Manual; OIE: Paris, France, 2019. [Google Scholar]
- Pineda, P.; Santa, C.; Deluque, A.; Peña, M.; Casal, J. Evaluation of the Sensitivity of the Classical Swine Fever Surveillance System in Two Free Zones in Colombia. Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef]
- Meyer, D.; Petrov, A.; Becher, P. Inactivation of Classical Swine Fever Virus in Porcine Serum Samples Intended for Antibody Detection. Pathogens 2019, 8, 286. [Google Scholar] [CrossRef]
- Bohórquez, J.A.; Muñoz-González, S.; Pérez-Simó, M.; Muñoz, I.; Rosell, R.; Coronado, L.; Domingo, M.; Ganges, L. Foetal Immune Response Activation and High. Replication Rate during Generation of Classical Swine Fever Congenital Infection. Pathogens 2020, 9, 285. [Google Scholar]
- Muñoz-González, S.; Ruggli, N.; Rosell, R.; Pérez, L.J.; Frías-Leuporeau, M.T.; Fraile, L.; Montoya, M.; Cordoba, L.; Domingo, M.; Ehrensperger, F.; et al. Postnatal Persistent Infection with Classical Swine Fever Virus and Its Immunological Implications. PLoS ONE 2015, 10, e0125692. [Google Scholar] [CrossRef] [PubMed]
- Kleiboeker, S.B. Swine Fever: Classical Swine Fever and African Swine Fever. Vet. Clin. N. Am. Food Anim. Pract. 2002, 18, 431–451. [Google Scholar] [CrossRef]
- Gómez-Villamandos, J.C.; Carrasco, L.; Bautista, M.J.; Sierra, M.A.; Quezada, M.; Hervas, J.; Mde, L.C.; Ruiz-Villamor, E.; Salguero, F.J.; Sónchez-Cordón, P.J.; et al. African Swine Fever and Classical Swine Fever: A Review of the Pathogenesis. Dtsch. Tierarztl. Wochenschr. 2003, 110, 165–169. [Google Scholar] [PubMed]
- Coronado, L.; Bohórquez, J.A.; Muñoz-González, S.; Perez, L.J.; Rosell, R.; Fonseca, O.; Delgado, L.; Perera, C.L.; Frías, M.T.; Ganges, L. Investigation of Chronic and Persistent Classical Swine Fever Infections under Field Conditions and Their Impact on Vaccine Efficacy. BMC Vet. Res. 2019, 15, 247. [Google Scholar] [CrossRef]
- Barman, N.N.; Khatoon, E.; Bora, M.; Deori, L.; Gogoi, S.M.; Kalita, D. Investigation of Congenital Tremor Associated with Classical Swine Fever Virus Genotype 2.2 in an Organized Pig Farm in North-Eastern India. Virus Dis. 2021, 32, 1–10. [Google Scholar]
- Beer, M.; Goller, K.V.; Staubach, C.; Blome, S. Genetic Variability and Distribution of Classical Swine Fever Virus. Anim. Health Res. Rev. 2015, 16, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Paton, D.J.; McGoldrick, A.; Greiser-Wilke, I.; Parchariyanon, S.; Song, J.Y.; Liou, P.P.; Stadejek, T.; Lowings, J.P.; Björklund, H.; Belák, S. Genetic Typing of Classical Swine Fever Virus. Vet. Microbiol. 2000, 73, 137–157. [Google Scholar] [CrossRef]
- Hao, G.; Zhang, H.; Chen, H.; Qian, P.; Li, X. Comparison of the Pathogenicity of Classical Swine Fever Virus Subgenotype 2.1c and 2.1d Strains from China. Pathogens 2020, 9, 821. [Google Scholar]
- Postel, A.; Austermann-Busch, S.; Petrov, A.; Moennig, V.; Becher, P. Epidemiology, Diagnosis and Control of Classical Swine Fever: Recent Developments and Future Challenges. Transbound. Emerg. Dis. 2018, 65 (Suppl. S1), 248–261. [Google Scholar] [CrossRef]
- Malik, Y.S.; Bhat, S.; Kumar, O.R.V.; Yadav, A.K.; Sircar, S.; Ansari, M.I.; Sarma, D.K.; Rajkhowa, T.K.; Ghosh, S.; Dhama, K. Classical Swine Fever Virus Biology, Clinicopathology, Diagnosis, Vaccines and a Meta-Analysis of Prevalence: A Review from the Indian Perspective. Pathogens 2020, 9, 500. [Google Scholar] [CrossRef]
- Li, Y.C.; Chiou, M.T.; Lin, C.N. Serodynamic Analysis of the Piglets Born from Sows Vaccinated with Modified Live Vaccine or E2 Subunit Vaccine for Classical Swine Fever. Pathogens 2020, 9, 427. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.; Wang, Z.; Li, X.; Zhang, S.; Paudyal, N.; Zhang, X.; Li, X.; Fang, W. E2 and E(Rns) of Classical Swine Fever Virus C-Strain Play Central Roles in Its Adaptation to Rabbits. Virus Genes 2019, 55, 238–242. [Google Scholar] [CrossRef]
- Cao, T.; Zhang, S.; Li, X.; Xu, Y.; Wang, Z.; Chen, C.; Paudyal, N.; Li, X.; Sun, J.; Fang, W. Classical Swine Fever Virus C-Strain with Eight Mutation Sites Shows Enhanced Cell Adaptation and Protects Pigs from Lethal Challenge. Arch. Virol. 2019, 164, 1619–1628. [Google Scholar] [CrossRef]
- Xie, L.; Han, Y.; Ma, Y.; Yuan, M.; Li, W.; Li, L.F.; Li, M.; Sun, Y.; Luo, Y.; Li, S.; et al. P108 and T109 on E2 Glycoprotein Domain I Are Critical for the Adaptation of Classical Swine Fever Virus to Rabbits but Not for Virulence in Pigs. J. Virol. 2020, 94, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Coronado, L.; Perera, C.L.; Rios, L.; Frías, M.T.; Pérez, L.J. A Critical Review about Different Vaccines against Classical Swine Fever Virus and Their Repercussions in Endemic Regions. Vaccines 2021, 9, 154. [Google Scholar] [CrossRef]
- Wei, Q.; Liu, Y.; Zhang, G. Research Progress and Challenges in Vaccine Development against Classical Swine Fever Virus. Viruses 2021, 13, 445. [Google Scholar] [CrossRef]
- Lamothe-Reyes, Y.; Bohórquez, J.A.; Wang, M.; Alberch, M.; Pérez-Simó, M.; Rosell, R.; Ganges, L. Early and Solid Protection Afforded by the Thiverval Vaccine Provides Novel Vaccination Alternatives Against Classical Swine Fever Virus. Vaccines 2021, 9, 464. [Google Scholar] [CrossRef]
- Jelsma, T.; Post, J.; Born, E.v.d.; Segers, R.; Kortekaas, J.J.V. Assessing the Protective Dose of a Candidate DIVA Vaccine against Classical Swine Fever. Vaccines 2021, 9, 483. [Google Scholar] [CrossRef] [PubMed]
- Villanueva-Cabezas, J.P.; Wangchuk, J. Classical Swine Fever: Challenges for the Emerging Swine Sector in Bhutan. Trop. Anim. Health Prod. 2020, 52, 2731–2735. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Rajak, K.K.; Kumar, A.; Yadav, S.K. Classical Swine Fever in India: Current Status and Future Perspective. Trop. Anim. Health Prod. 2018, 50, 1181–1191. [Google Scholar] [CrossRef]
- Huang, Y.L.; Deng, M.C.; Wang, F.I.; Huang, C.C.; Chang, C.Y. The Challenges of Classical Swine Fever Control: Modified Live and E2 Subunit Vaccines. Virus Res. 2014, 179, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Je, S.H.; Kwon, T.; Yoo, S.J.; Lee, D.U.; Lee, S.; Richt, J.A.; Lyoo, Y.S. Classical Swine Fever Outbreak after Modified Live LOM Strain Vaccination in Naive Pigs, South Korea. Emerg. Infect. Dis. 2018, 24, 798–800. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.; Kim, J.A.; Kang, W.M.; Yang, H.S.; Park, C.; Jeong, K.; Moon, S.U.; Park, C.K.; Lyoo, Y.S.; Lee, C. Endemic Outbreaks Due to the Re-Emergence of Classical Swine Fever after Accidental Introduction of Modified Live LOM Vaccine on Jeju Island, South Korea. Transbound. Emerg. Dis. 2019, 66, 634–639. [Google Scholar] [CrossRef]
- Blome, S.; Moß, C.; Reimann, I.; König, P.; Beer, M. Classical Swine Fever Vaccines-State-of the-Art. Vet. Microbiol. 2017, 206, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Blome, S.; Wernike, K.; Reimann, I.; König, P.; Moß, C.; Beer, M. A Decade of Research into Classical Swine Fever Marker Vaccine CP7_E2alf (Suvaxyn® CSF Marker): A Review of Vaccine Properties. Vet. Res. 2017, 48, 51. [Google Scholar] [CrossRef] [PubMed]
- Sordo-Puga, Y.; Suárez-Pedroso, M.; Naranjo-Valdéz, P.; Pérez-Pérez, D.; Santana-Rodríguez, E.; Sardinas-Gonzalez, T.; Mendez-Orta, M.K.; Duarte-Cano, C.A.; Estrada-Garcia, M.P.; Rodríguez-Moltó, M.P. Porvac® Subunit Vaccine E2-CD154 Induces Remarkable Rapid Protection against Classical Swine Fever Virus. Vaccines 2021, 9, 167. [Google Scholar] [CrossRef]
- Park, Y.; Oh, Y.; Wang, M.; Ganges, L.; Bohórquez, J.A.; Park, S.; Gu, S.; Park, J.; Lee, S.; Kim, J.; et al. A Novel E2 Glycoprotein Subunit Marker Vaccine Produced in Plant Is Able to Prevent Classical Swine Fever Virus Vertical Transmission after Double Vaccination. Vaccines 2021, 9, 418. [Google Scholar]
- Tong, W.; Zheng, H.; Li, G.X.; Gao, F.; Shan, T.L.; Zhou, Y.J.; Yu, H.; Jiang, Y.F.; Yu, L.X.; Li, L.W.; et al. Recombinant Pseudorabies Virus Expressing E2 of Classical Swine Fever Virus (CSFV) Protects against Both Virulent Pseudorabies Virus and CSFV. Antivir. Res. 2020, 173, 104652. [Google Scholar] [CrossRef]
- Kumar, R.; Kumar, V.; Kekungu, P.; Barman, N.N.; Kumar, S. Evaluation of Surface Glycoproteins of Classical Swine Fever Virus as Immunogens and Reagents for Serological Diagnosis of Infections in Pigs: A Recombinant Newcastle Disease Virus Approach. Arch. Virol. 2019, 164, 3007–3017. [Google Scholar] [CrossRef]
- Ganges, L.; Crooke, H.R.; Bohórquez, J.A.; Postel, A.; Sakoda, Y.; Becher, P.; Ruggli, N. Classical Swine Fever Virus: The Past, Present and Future. Virus Res. 2020, 289, 198151. [Google Scholar] [CrossRef]
- Backer, J.; Brouwer, H.; van Schaik, G.; Van Roermund, H. Using Mortality Data for Early Detection of Classical Swine Fever in The Netherlands. Prev. Vet. Med. 2011, 99, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Backer, J.A.; Hagenaars, T.J.; van Roermund, H.J.; de Jong, M.C. Modelling the Effectiveness and Risks of Vaccination Strategies to Control Classical Swine Fever Epidemics. J. R. Soc. Interface 2009, 6, 849–861. [Google Scholar] [CrossRef]
- Fan, J.; Liao, Y.; Zhang, M.; Liu, C.; Li, Z.; Li, Y.; Li, X.; Wu, K.; Yi, L.; Ding, H.; et al. Anti-Classical Swine Fever Virus Strategies. Microorganisms 2021, 9, 761. [Google Scholar] [CrossRef]
- Artois, M.; Depner, K.; Guberti, V.; Hars, J.; Rossi, S.; Rutili, D. Classical Swine Fever (Hog Cholera) in Wild Boar in Europe. Rev. Sci. Tech. l’OIE 2002, 21, 287–303. [Google Scholar] [CrossRef] [PubMed]
- Isoda, N.; Baba, K.; Ito, S.; Ito, M.; Sakoda, Y.; Makita, K. Dynamics of Classical Swine Fever Spread in Wild Boar in 2018–2019, Japan. Pathogens 2020, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Jurado, C.; Bosch, J.; Sánchez-Vizcaíno, J.M.; Isoda, N.; Sakoda, Y. Role of Wild Boar in the Spread of Classical Swine Fever in Japan. Pathogens 2019, 8, 206. [Google Scholar] [CrossRef] [PubMed]
- Moennig, V. The Control of Classical Swine Fever in Wild Boar. Front. Microbiol. 2015, 6, 1211. [Google Scholar] [CrossRef]
- Shimizu, Y.; Hayama, Y.; Murato, Y.; Sawai, K.; Yamaguchi, E.; Yamamoto, T. Epidemiology of Classical Swine Fever in Japan—A Descriptive Analysis of the Outbreaks in 2018–2019. Front. Vet. Sci. 2020, 7, 573480. [Google Scholar] [CrossRef]
- Shimizu, Y.; Hayama, Y.; Murato, Y.; Sawai, K.; Yamaguchi, E.; Yamamoto, T. Epidemiological Analysis of Classical Swine Fever in Wild Boars in Japan. BMC Vet. Res. 2021, 17, 188. [Google Scholar] [CrossRef]
- Kameyama, K.I.; Nishi, T.; Yamada, M.; Masujin, K.; Morioka, K.; Kokuho, T.; Fukai, K. Experimental Infection of Pigs with a Classical Swine Fever Virus Isolated in Japan for the First Time in 26 Years. J. Vet. Med. Sci. 2019, 81, 1277–1284. [Google Scholar] [CrossRef]
- Jelsma, T.; Wijnker, J.J.; Smid, B.; Verheij, E.; van der Poel, W.H.M.; Wisselink, H.J. Salt Inactivation of Classical Swine Fever Virus and African Swine Fever Virus in Porcine Intestines Confirms the Existing In Vitro Casings Model. Vet. Microbiol. 2019, 238, 108424. [Google Scholar] [CrossRef]
- Badasara, S.K.; Mohan, M.; Sah, V.; Kumari, P.; Upmanyu, V.; Dhar, P.; Tiwari, A.K.; Chander, V.; Gupta, V.K. Replacement of Animal Model for Propagation of Classical Swine Fever Challenge Virus by Adaption in the PK-15 Cell Line. J. Anim. Res. 2017, 7, 581. [Google Scholar] [CrossRef]
- Sordo, Y.; Suárez, M.; Caraballo, R.; Sardina, T.; Brown, E.; Duarte, C.; Lugo, J.; Gil, L.; Perez, D.; Oliva, A.; et al. Humoral and Cellular Immune Response in Mice Induced by the Classical Swine Fever Virus E2 Protein Fused to the Porcine CD154 Antigen. Biologicals 2018, 52, 67–71. [Google Scholar] [CrossRef]
- Di Teodoro, G.; Marruchella, G.; Di Provvido, A.; D’Angelo, A.R.; Orsini, G.; Di Giuseppe, P.; Sacchini, F.; Scacchia, M. Contagious Bovine Pleuropneumonia: A Comprehensive Overview. Vet. Pathol. 2020, 57, 476–489. [Google Scholar] [CrossRef]
- Amanfu, W. Contagious Bovine Pleuropneumonia, in Transboundary Animal Diseases in Sahelian Africa and Connected Regions; Kardjadj, M., Ed.; Springer Nature: Cham, Switzerland, 2019; pp. 423–437. [Google Scholar]
- Dupuy, V.; Manso-Silván, L.; Barbe, V.; Thebault, P.; Dordet-Frisoni, E.; Citti, C.; Poumarat, F.; Blanchard, A.; Breton, M.; Sirand-Pugnet, P.; et al. Evolutionary History of Contagious Bovine Pleuropneumonia Using Next Generation Sequencing of Mycoplasma Mycoides Subsp. Mycoides “Small Colony”. PLoS ONE 2012, 7, e46821. [Google Scholar] [CrossRef]
- Jores, J.; Baldwin, C.; Blanchard, A.; Browning, G.F.; Colston, A.; Gerdts, V.; Goovaerts, D.; Heller, M.; Juleff, N.; Labroussaa, F.; et al. Contagious Bovine and Caprine Pleuropneumonia: A Research Community’s Recommendations for the Development of Better Vaccines. NPJ Vaccines 2020, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Amanfu, W. Contagious Bovine Pleuropneumonia (Lung Sickness) in Africa. Onderstepoort J. Vet. Res. 2009, 76, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Dedieu-Engelmann, L. Contagious Bovine Pleuropneumonia: A Rationale for the Development of a Mucosal Sub-Unit Vaccine. Comp. Immunol. Microbiol. Infect. Dis. 2008, 31, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Thiaucourt, F.; Yaya, A.; Wesonga, H.; Huebschle, O.J.; Tulasne, J.J.; Provost, A. Contagious Bovine Pleuropneumonia: A Reassessment of the Efficacy of Vaccines Used in Africa. Ann. N. Y. Acad. Sci. 2006, 916, 71–80. [Google Scholar] [CrossRef]
- Olorunshola, I.D.; Peters, A.R.; Scacchia, M.; Nicholas, R.A.J. Contagious Bovine Pleuropneumonia—Never Out of Africa? CAB Rev. 2017, 12, 1–7. [Google Scholar] [CrossRef]
- Mamo, Y.; Bitew, M.; Teklemariam, T.; Soma, M.; Gebre, D.; Abera, T.; Benti, T.; Deneke, Y. Contagious Bovine Pleuropneumonia: Seroprevalence and Risk Factors in Gimbo District, Southwest Ethiopia. Vet. Med. Int. 2018, 2018, 1–7. [Google Scholar] [CrossRef]
- Zerbo, L.H.; Dahourou, L.D.; Sidi, M.; Ouoba, L.B.; Ouandaogo, S.H.; Bazimo, G.; Sie, B.N.; Traore, K.Z.A.; Tapsoba, M.; Ouedraogo, A.; et al. Seroprevalence and Determinants of Contagious Bovine Pleuropneumonia in Cattle in Burkina Faso. Trop. Anim. Health Prod. 2021, 53, 1–8. [Google Scholar] [CrossRef]
- Nicholas, R.; Bashiruddin, J.; Ayling, R.; Miles, R. Contagious Bovine Pleuropneumonia: A Review of Recent Developments. Vet. Bull. 2000, 70, 827–838. [Google Scholar]
- Abdela, N.; Yune, N. Seroprevalence and Distribution of Contagious Bovine Pleuropneumonia in Ethiopia: Update and Critical Analysis of 20 Years (1996–2016) Reports. Front. Vet. Sci. 2017, 4, 100. [Google Scholar] [CrossRef] [PubMed]
- Yansambou, M.S.; Diallo, A.A.; Idi, M.; Gagara, H.; Haido, A.M.; Alambedji, R.B. Serological Prevalence of Contagious Bovine Pleuropneumonia in Niger in 2017. Front. Vet. Sci. 2018, 5, 238. [Google Scholar] [CrossRef] [PubMed]
- Thiaucourt, F.; van der Lugt, J.J.; Provost, A. Contagious Bovine Pleuropneumonia. In Infectious Diseases of Livestock; Coetzer, J.A., Tustin, R.C., Eds.; Oxford University Press Southern Africa: Cape Town, South Africa, 2004; pp. 2045–2057. [Google Scholar]
- Masiga, W.; Domenech, J.; Windsor, R. Manifestation and Epidemiology of Contagious Bovine Pleuropneumonia in Africa. Rev. Sci. Tech. 1996, 15, 1283–1308. [Google Scholar] [CrossRef] [PubMed]
- Windsor, R.; Masiga, W. Investigations into the Role of Carrier Animals in the Spread of Contagious Bovine Pleuropneumonia. Res. Vet. Sci. 1977, 23, 224–229. [Google Scholar] [CrossRef]
- Lesnoff, M.; Bonnet, P.; Workalemahu, A. A Mathematical Model of Contagious Bovine Pleuropneumonia (CBPP) within-Herd Outbreaks for Economic Evaluation of Local Control Strategies: An Illustration from a Mixed Crop-Livestock System in Ethiopian Highlands. Anim. Res. 2004, 53, 429–438. [Google Scholar] [CrossRef]
- Windsor, R.S. The Eradication of Contagious Bovine Pleuropneumonia from South Western Africa: A Plan for Action. Ann. New York Acad. Sci. 2006, 916, 326–332. [Google Scholar] [CrossRef]
- Le Goff, C.; Thiaucourt, F. A Competitive ELISA for the Specific Diagnosis of Contagious Bovine Pleuropneumonia (CBPP). Vet. Microbiol. 1998, 60, 179–191. [Google Scholar] [CrossRef]
- Ayling, R.D.; Bisgaard-Frantzen, S.; March, J.B.; Godinho, K.; Nicholas, R.A.J. Assessing the In Vitro Effectiveness of Antimicrobials against Mycoplasma Mycoides Subsp. Mycoides Small-Colony Type to Reduce Contagious Bovine Pleuropneumonia Infection. Antimicrob. Agents Chemother. 2005, 49, 5162–5165. [Google Scholar] [CrossRef] [PubMed]
- Huebschle, O.J.; Ayling, R.D.; Godinho, K.; Lukhele, O.; Tjipura-Zaire, G.; Rowan, T.G.; Nicholas, R.A. Danofloxacin (Advocin™) Reduces the Spread of Contagious Bovine Pleuropneumonia to Healthy In-Contact Cattle. Res. Vet. Sci. 2006, 81, 304–309. [Google Scholar] [CrossRef]
- Danbirni, S.; Kaltungo, B.Y.; Dandu, B.B.; Mohammed, F.U.; Abubakar, U.B.; Ibrahim, S.; Usman, A.; Sackey, A.K.B. Clinical Management Protocol of an Acute Contagious Bovine Pleuropneumonia in a 6-Year-Old Bunaji Cow in Sakaru Village of Soba Local Gov-Ernment Area, Kaduna State, Nigeria. Niger. J. Anim. Prod. 2020, 47, 51–56. [Google Scholar] [CrossRef]
- Tulasne, J.J.; Litamoi, J.K.; Morein, B.; Dedieu, L.; Palya, V.J.; Yami, M.; Abusugra, I.; Sylla, D.; Bensaid, A. Contagious Bovine Pleuropneumonia Vaccines: The Current Situation and the Need for Improvement. Rev. Sci. Tech. 1996, 15, 1373–1396. [Google Scholar] [CrossRef] [PubMed]
- Sacchini, F.; Luciani, M.; Salini, R.; Scacchia, M.; Pini, A.; Lelli, R.; Naessens, J.; Poole, J.; Jores, J. Plasma lLevels of TNF-alpha, IFN-gamma, IL-4 and IL-10 during a Course of Experimental Contagious Bovine Pleuropneumonia. BMC Vet. Res. 2012, 8, 44. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, R.A.J.; Ayling, R.D.; Tjipura-Zaire, G.; Rowan, T. Treatment of Contagious Bovine Pleuropneumonia. Vet. Rec. 2012, 171, 510–511. [Google Scholar] [CrossRef]
- Hamsten, C.; Tjipura-Zaire, G.; McAuliffe, L.; Huebschle, O.J.B.; Scacchia, M.; Ayling, R.D.; Persson, A. Protein-Specific Analysis of Humoral Immune Responses in a Clinical Trial for Vaccines against Contagious Bovine Pleuropneumonia. Clin. Vaccine Immunol. 2010, 17, 853–861. [Google Scholar] [CrossRef]
- Jores, J.; Mariner, J.C.; Naessens, J. Development of an Improved Vaccine for Contagious Bovine Pleuropneumonia: An African Perspective on Challenges and Proposed Actions. Vet. Res. 2013, 44, 122. [Google Scholar] [CrossRef]
- Westberg, J.; Persson, A.; Holmberg, A.; Goesmann, A.; Lundeberg, J.; Johansson, K.-E.; Pettersson, B.; Uhlén, M. The Genome Sequence of Mycoplasma Mycoides Subsp. Mycoides SC Type Strain PG1T, the Causative Agent of Contagious Bovine Pleuropneumonia (CBPP). Genome Res. 2004, 14, 221–227. [Google Scholar] [CrossRef]
- Windsor, R.S.; Wood, A. Contagious Bovine Pleuropneumonia: The Costs of Control in Central/Southern Africa. Ann. New York Acad. Sci. 1998, 849, 299–306. [Google Scholar] [CrossRef]
- Ter Laak, E. Contagious Bovine Pleuropneumonia A Review. Vet. Q. 1992, 14, 104–110. [Google Scholar] [CrossRef]
- Nicholas, R.; Bashiruddin, J. Mycoplasma Mycoides Subspecies Mycoides (Small Colony Variant): The Agent of Contagious Bovine Pleuropneumonia and Member of the “Mycoplasma Mycoides Cluster”. J. Comp. Pathol. 1995, 113, 1–27. [Google Scholar] [CrossRef]
- Alhaji, N.B.; Ankeli, P.I.; Ikpa, L.T.; Babalobi, O.O. Contagious Bovine Pleuropneumonia: Challenges and Prospects Regarding Diagnosis and Control Strategies in Africa. Vet. Med. Res. Rep. 2020, 11, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Marobela-Raborokgwe, C.; Modise, B.; Kgotlele, T.; Masoba, K.; Keokilwe, L.; Dipuo, K. The Experience of Contagious Bovine Pleuropneumonia Ring Trials in Botswana. Rev. Sci. Tech. 2018, 37, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Rweyemamu, M.; Litamoi, J.; Palya, V.; Sylla, D. Contagious Bovine Pleuropneumonia Vaccines: The Need for Improvements. Rev. Sci. Tech. 1995, 14, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Ssematimba, A.; Jores, J.; Mariner, J.C. Mathematical Modelling of the Transmission Dynamics of Contagious Bovine Pleuropneu-Monia Reveals Minimal Target Profiles for Improved Vaccines and Diagnostic Assays. PLoS ONE 2015, 10, e0116730. [Google Scholar] [CrossRef]
- Kusiluka, L.; Sudi, F. Review of Successes and Failures of Contagious Bovine Pleuropneumonia Control Strategies in Tanzania. Prev. Vet. Med. 2003, 59, 113–123. [Google Scholar] [CrossRef]
- Marobela-Raborokgwe, C. Contagious Bovine Pleuropneumonia in Botswana: Experience with Control, Eradication, Prevention and Surveillance. Vet. Ital. 2011, 47, 397–405. [Google Scholar]
- Muuka, G.; Songolo, N.; Kabilika, S.; Hang’ombe, B.M.; Nalubamba, K.S.; Muma, J.B. Challenges of Controlling Contagious Bovine Pleuropneumonia in Sub-Saharan Africa: A Zambian Perspective. Trop. Anim. Health Prod. 2013, 45, 9–15. [Google Scholar] [CrossRef]
- Onono, J.; Wieland, B.; Suleiman, A.; Rushton, J. Policy Analysis for Delivery of Contagious Bovine Pleuropneumonia Control Strategies in Sub-Saharan Africa. Rev. Sci. Tech. 2017, 36, 195–205. [Google Scholar] [CrossRef]
- Tambi, N.E.; Maina, W.O.; Ndi, C. An Estimation of the Economic Impact of Contagious Bovine Pleuropneumonia in Africa. Rev. Sci. Tech. 2006, 25, 999–1011. [Google Scholar] [CrossRef]
- Aligaz, A.A.; Munganga, J.M.W. Modelling the Transmission Dynamics of Contagious Bovine Pleuropneumonia in the Presence of Antibiotic Treatment with Limited Medical Supply. Math. Model. Anal. 2021, 26, 1–20. [Google Scholar] [CrossRef]
- Mariner, J.; McDermott, J.; Heesterbeek, J.; Thomson, G.; Martin, S. A Model of Contagious Bovine Pleuropneumonia Transmission Dynamics in East Africa. Prev. Vet. Med. 2006, 73, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Mariner, J.; McDermott, J.; Heesterbeek, J.; Thomson, G.; Roeder, P.; Martin, S. A Heterogeneous Population Model for Contagious Bovine Pleuropneumonia Transmission and Control in Pastoral Communities of East Africa. Prev. Vet. Med. 2006, 73, 75–91. [Google Scholar] [CrossRef][Green Version]
- Aligaz, A.A.; Munganga, J.M.W. Mathematical Modelling of the Transmission Dynamics of Contagious Bovine Pleuropneumonia with Vaccination and Antibiotic Treatment. J. Appl. Math. 2019, 2019, 1–10. [Google Scholar] [CrossRef]
- Aye, R.; Weldearegay, Y.B.; Lutta, H.O.; Chuma, F.; Pich, A.; Jores, J.; Meens, J.; Naessens, J. Identification of Targets of Monoclonal Antibodies that Inhibit Adhesion and Growth in Mycoplasma Mycoides Subspecies Mycoides. Vet. Immunol. Immunopathol. 2018, 204, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Gaurivaud, P.; Lakhdar, L.; le Grand, D.; Poumarat, F.; Tardy, F. Comparison of In Vivo and In Vitro Properties of Capsulated and Noncapsulated Variants of Mycoplasma Mycoides Subsp. Mycoides Strain Afade: A Potential New Insight into the Biology of Contagious Bovine Pleuropneumonia. FEMS Microbiol. Lett. 2014, 359, 42–49. [Google Scholar] [CrossRef]
- Di Teodoro, G.; Marruchella, G.; Di Provvido, A.; Orsini, G.; Ronchi, G.F.; D’Angelo, A.R.; D’Alterio, N.; Sacchini, F.; Scacchia, M. Respiratory Explants as a Model to Investigate Early Events of Contagious Bovine Pleuropneumonia Infection. Vet. Res. 2018, 49, 5. [Google Scholar] [CrossRef]
- Waite, E.; March, J. Capsular Polysaccharide Conjugate Vaccines against Contagious Bovine Pleuropneumonia: Immune Responses and Protection in Mice. J. Comp. Pathol. 2002, 126, 171–182. [Google Scholar] [CrossRef]
- Mwirigi, M.; Nkando, I.; Olum, M.; Attah-Poku, S.; Ochanda, H.; Berberov, E.; Potter, A.; Gerdts, V.; Perez-Casal, J.; Wesonga, H.; et al. Capsular Polysaccharide from Mycoplasma Mycoides Subsp. Mycoides Shows Potential for Protection against Contagious Bovine Pleuropneumonia. Vet. Immunol. Immunopathol. 2016, 178, 64–69. [Google Scholar] [CrossRef]
- Gull, T.; French, R.A.; Gorton, T.S.; Burrage, T.G.; Prozesky, L.; Geary, S.J.; Adams, L.G. Models of Contagious Bovine Pleuro-pneumonia: Evaluation of Two Novel Strains. Open Vet. Sci. J. 2013, 7, 23–33. [Google Scholar] [CrossRef]
- Nkando, I.G.; Wesonga, H.O.; Kuria, J.K.; McKeever, D. Assessing the Effectiveness of Intubation as a Challenge Model in Conta-Gious Bovine Pleuropneumonia Vaccine Experiments. Trop. Anim. Health Prod. 2010, 42, 1743–1747. [Google Scholar] [CrossRef] [PubMed]
- Sacchini, F.; Liljander, A.M.; Heller, M.; Poole, E.J.; Posthaus, H.; Schieck, E.; Jores, J. Reproduction of Contagious Bovine Pleu-Ropneumonia via Aerosol-Based Challenge with Mycoplasma Mycoides Subsp. Mycoides. Acta Vet. Scand. 2020, 62, 62. [Google Scholar] [CrossRef] [PubMed]
- Mahy, B.W.J. Introduction and History of Foot-and-Mouth Disease Virus; Springer: Berlin/Heidelberg, Germany, 2005; pp. 1–8. [Google Scholar]
- Grubman, M.J.; Baxt, B. Foot-and-Mouth Disease. Clin. Microbiol. Rev. 2004, 17, 465–493. [Google Scholar] [CrossRef]
- Jamal, S.M.; Belsham, G.J. Foot-and-Mouth Disease: Past, Present and Future. Vet. Res. 2013, 44, 116. [Google Scholar] [CrossRef]
- Segundo, F.D.-S.; Medina, G.N.; Stenfeldt, C.; Arzt, J.; Santos, T.D.L. Foot-and-Mouth Disease Vaccines. Vet. Microbiol. 2017, 206, 102–112. [Google Scholar] [CrossRef]
- Alexandersen, S.; Mowat, N. Foot-and-Mouth Disease: Host Range and Pathogenesis. Curr. Top. Microbiol. 2005, 288, 9–42. [Google Scholar]
- Thomson, G.; Vosloo, W.; Bastos, A. Foot and Mouth Disease in Wildlife. Virus Res. 2003, 91, 145–161. [Google Scholar] [CrossRef]
- Brito, B.P.; Rodriguez, L.L.; Hammond, J.M.; Pinto, J.; Perez, A.M. Review of the Global Distribution of Foot-and-Mouth Disease Virus from 2007 to 2014. Transbound. Emerg. Dis. 2017, 64, 316–332. [Google Scholar] [CrossRef]
- Knight-Jones, T.J.D.; Robinson, L.; Charleston, B.; Rodriguez, L.L.; Gay, C.G.; Sumption, K.J.; Vosloo, W. Global Foot-and-Mouth Disease Research Update and Gap Analysis: 2—Epidemiology, Wildlife and Economics. Transbound. Emerg. Dis. 2016, 63 (Suppl. S1), 14–29. [Google Scholar] [CrossRef]
- Gibbs, E.P.; A Herniman, K.; Lawman, M.J.; Sellers, R.F. Foot-and-Mouth Disease in British Deer: Transmission of Virus to Cattle, Sheep and Deer. Vet. Rec. 1975, 96, 558–563. [Google Scholar] [CrossRef]
- Kittelberger, R.; Nfon, C.; Swekla, K.; Zhang, Z.; Hole, K.; Bittner, H.; Salo, T.; Goolia, M.; Embury-Hyatt, C.; Bueno, R.; et al. Foot-and-Mouth Disease in Red Deer—Experimental Infection and Test. Methods Perform. 2017, 64, 213–225. [Google Scholar]
- Paton, D.J.; Gubbins, S.; King, D.P. Understanding the Transmission of Foot-and-Mouth Disease Virus at Different Scales. Curr. Opin. Virol. 2018, 28, 85–91. [Google Scholar] [CrossRef]
- Donaldson, A.I.; Gloster, J.; Harvey, L.D.; Deans, D.H. Use of Prediction Models to Forecast and Analyse Airborne Spread during the Foot-and-Mouth Disease Outbreaks in Brittany, Jersey and the Isle of Wight in 1981. Vet. Rec. 1982, 110, 53–57. [Google Scholar] [CrossRef]
- Knight-Jones, T.; Rushton, J. The Economic Impacts of Foot and Mouth Disease—What Are They, how Big Are They and where Do They Occur? Prev. Vet. Med. 2013, 112, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Davies, G. The Foot and Mouth Disease (FMD) Epidemic in the United Kingdom 2001. Comp. Immunol. Microbiol. Infect. Dis. 2002, 25, 331–343. [Google Scholar] [CrossRef]
- Doel, T.R. Natural and Vaccine Induced Immunity to FMD. In Foot-and-Mouth Disease Virus; Mahy, B.W.J., Ed.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 103–131. [Google Scholar]
- Sutmoller, P.; Gaggero, A. Foot-and Mouth Diseases Carriers. Vet. Rec. 1965, 77, 968–969. [Google Scholar] [CrossRef] [PubMed]
- Ferris, N.; King, D.; Reid, S.; Hutchings, G.; Shaw, A.; Paton, D.; Goris, N.; Haas, B.; Hoffmann, B.; Brocchi, E.; et al. Foot-and-Mouth Disease Virus: A First Inter-Laboratory Comparison Trial to Evaluate Virus Isolation and RT-PCR Detection Methods. Vet. Microbiol. 2006, 117, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Sobrino, F.; Dávila, M.; Ortín, J.; Domingo, E. Multiple Genetic Variants Arise in the Course of Replication of Foot-and-Mouth Disease Virus in Cell Culture. Virology 1983, 128, 310–318. [Google Scholar] [CrossRef]
- Gray, A.R.; Wood, B.A.; Henry, E.; Azhar, M.; King, D.P.; Mioulet, V. Evaluation of Cell Lines for the Isolation of Foot-and-Mouth Disease Virus and Other Viruses Causing Vesicular Disease. Front. Vet. Sci. 2020, 7, 426. [Google Scholar] [CrossRef]
- Kopliku, L.; Relmy, A.; Romey, A.; Gorna, K.; Zientara, S.; Bakkali-Kassimi, L.; Blaise-Boisseau, S. Establishment of Persistent Foot-and-Mouth Disease Virus (FMDV) Infection in MDBK Cells. Arch. Virol. 2015, 160, 2503–2516. [Google Scholar] [CrossRef]
- O’Donnell, V.; Pacheco, J.; Larocco, M.; Gladue, D.; Pauszek, S.; Smoliga, G.; Krug, P.; Baxt, B.; Borca, M.; Rodriguez, L. Virus–Host Interactions in Persistently FMDV-Infected Cells Derived from Bovine Pharynx. Virology 2014, 468–470, 185–196. [Google Scholar] [CrossRef] [PubMed]
- De la Torre, J.C.; Dávila, M.; Sobrino, F.; Ortín, J.; Domingo, E. Establishment of Cell Lines Persistently Infected with Foot-and-Mouth Disease Virus. Virology 1985, 145, 24–35. [Google Scholar] [CrossRef]
- Hägglund, S.; Laloy, E.; Näslund, K.; Pfaff, F.; Eschbaumer, M.; Romey, A.; Relmy, A.; Rikberg, A.; Svensson, A.; Huet, H.; et al. Model of Persistent Foot-and-Mouth Disease Virus Infection in Multilayered Cells Derived from Bovine Dorsal Soft Palate. Transbound. Emerg. Dis. 2019, 67, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Habiela, M.; Seago, J.; Perez-Martin, E.; Waters, R.; Windsor, M.; Salguero, F.J.; Wood, J.; Charleston, B.; Juleff, N. Laboratory Animal Models to Study Foot-and-Mouth Disease: A Review with Emphasis on Natural and Vaccine-Induced Immunity. J. Gen. Virol. 2014, 95, 2329–2345. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.J.; Wolfson, J.S.; Swartz, M.N.; Hooper, D.C. Pasteurella Multocida Infections. Report Of 34 Cases and Review of the Literature. Medicine 1984, 63, 133–154. [Google Scholar] [CrossRef] [PubMed]
- Shivachandra, S.B.; Viswas, K.N.; Kumar, A.A. A Review of Hemorrhagic Septicemia in Cattle and Buffalo. Anim. Health Res. Rev. 2011, 12, 67–82. [Google Scholar] [CrossRef]
- Biberstein, E.L. Our Understanding of the Pasteurellaceae. Can. J. Vet. Res. 1990, 54, S78–S82. [Google Scholar]
- Wilson, B.A.; Ho, M. Pasteurella Multocida: From Zoonosis to Cellular Microbiology. Clin. Microbiol. Rev. 2013, 26, 631–655. [Google Scholar] [CrossRef]
- Raffi, F.; Barrier, J.; Baron, D.; Drugeon, H.B.; Nicolas, F.; Courtieu, A.L. Pasteurella Multocida Bacteremia: Report of Thirteen Cases over Twelve Years and Review of the Literature. Scand. J. Infect. Dis. 1987, 19, 385–393. [Google Scholar] [CrossRef]
- Hunt, M.L.; Boucher, D.J.; Boyce, J.D.; Adler, B. In Vivo-Expressed Genes of Pasteurella multocida. Infect. Immun. 2001, 69, 3004–3012. [Google Scholar] [CrossRef]
- Hurtado, R.; Maturrano, L.; Azevedo, V.; Aburjaile, F. Pathogenomics Insights for Understanding Pasteurella Multocida Adaptation. Int. J. Med. Microbiol. 2020, 310, 151417. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Wang, X.; Zhou, R.; Chen, H.; Wilson, B.A.; Wu, B. Pasteurella Multocida: Genotypes and Genomics. Microbiol. Mol. Biol. Rev. 2019, 83, 83. [Google Scholar] [CrossRef]
- Harper, M.; Boyce, J.; Adler, B. Pasteurella Multocida Pathogenesis: 125 Years after Pasteur. FEMS Microbiol. Lett. 2006, 265, 1–10. [Google Scholar] [CrossRef]
- Tankaew, P.; Srisawat, W.; Singhla, T.; Tragoolpua, K.; Kataoka, Y.; Sawada, T.; Sthitmatee, N. Comparison of Two Indirect ELISA Coating Antigens for the Detection of Dairy Cow Antibodies Against Pasteurella Multocida. J. Microbiol. Methods 2018, 145, 20–27. [Google Scholar] [CrossRef]
- Harper, M.; Cox, A.; Adler, B.; Boyce, J. Pasteurella Multocida Lipopolysaccharide: The Long and the Short of It. Vet. Microbiol. 2011, 153, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.B.; Dawkins, H.J.; Spencer, T.L. Electrophoretic Profiles of Pasteurella Multocida Isolates from Animals with Hemor-Rhagic Septicemia. Am. J. Vet. Res. 1991, 52, 1644–1648. [Google Scholar]
- De Alwis, M.C. Haemorrhagic Septicaemia—A General Review. Br. Vet. J. 1992, 148, 99–112. [Google Scholar] [CrossRef]
- Eriksen, L.; Aalbaek, B.; Leifsson, P.S.; Basse, A.; Christiansen, T.; Eriksen, E.; Rimler, R.B. Hemorrhagic Septicemia in Fallow Deer (Dama Dama) Caused by Pasteurella Multocida Multocida. J. Zoo Wildl. Med. 1999, 30, 285–292. [Google Scholar] [PubMed]
- Khairani, K.O.; Nydam, D.; Felippe, M.J.; McDonough, P.; Barry, J.; Mahmud, R.; Haryono, M.; Radcliffe, R.W. Surveillance for Hemorrhagic Septicemia in Buffalo (Bubalus Bubalis) as an Aid to Range Expansion of the Javan Rhinoceros (Rhinoceros Sondaicus) in Ujung Kulon National Park, Indonesia. J. Wildl. Dis. 2018, 54, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Kock, R.A.; Orynbayev, M.; Robinson, S.; Zuther, S.; Singh, N.J.; Beauvais, W.; Morgan, E.R.; Kerimbayev, A.; Khomenko, S.; Martineau, H.M.; et al. Saigas on the Brink: Multidisci-Plinary Analysis of the Factors Influencing Mass Mortality Events. Sci. Adv. 2018, 4, eaao2314. [Google Scholar] [CrossRef]
- Tankaew, P.; Singh-La, T.; Titaram, C.; Punyapornwittaya, V.; Vongchan, P.; Sawada, T.; Sthitmatee, N. Evaluation of an In-House Indirect ELISA for Detection of Antibody against Haemorrhagic Septicemia in Asian Elephants. J. Microbiol. Methods 2017, 134, 30–34. [Google Scholar] [CrossRef]
- Marza, A.D.; Abdullah, F.F.J.; Ahmed, I.M.; Chung, E.L.; Ibrahim, H.H.; Zamri-Saad, M.; Omar, A.R.; Bakar, M.Z.; Sa-haree, A.A.; Haron, A.; et al. Involvement of Nervous System in Cattle and Buffaloes Due to Pasteurella Multocida B:2 Infection: A Review of Clinicopathological and Pathophysiological Changes. J. Adv. Vet. Anim. Res. 2015, 2, 252–262. [Google Scholar] [CrossRef]
- Jilo, K.; Belachew, T.; Birhanu, W.; Habte, D.; Yadeta, W.; Giro, A. Pasteurellosis Status in Ethiopia: A Comprehensive Review. J. Trop. Dis. Public Health 2020, 8, 1–5. [Google Scholar]
- Fereidouni, S.; Freimanis, G.L.; Orynbayev, M.; Ribeca, P.; Flannery, J.; King, D.P.; Zuther, S.; Beer, M.; Hoper, D.; Kydyrmanov, A.; et al. Mass Die-Off. of Saiga Antelopes, Kazakhstan, 2015. Emerg. Infect. Dis. 2019, 25, 1169–1176. [Google Scholar] [CrossRef]
- Robinson, S.; Milner-Gulland, E.J.; Grachev, Y.; Salemgareyev, A.; Orynbayev, M.; Lushchekina, A.; Morgan, E.; Beauvais, W.; Singh, N.; Khomenko, S.; et al. Opportunistic Bacteria and Mass Mortality in Ungulates: Lessons from an Extreme Event. Ecosphere 2019, 10, e02671. [Google Scholar] [CrossRef]
- Khan, A.; Saleemi, M.K.; Khan, M.Z.; Gul, S.T.; Irfan, M.; Qamar, M.S. Hemorrhagic Septicemia in Buffalo (Bubalus bubalis) Calves under Sub-Tropical Conditions in Pakistan. Pak. J. Zool. 2011, 43, 295–302. [Google Scholar]
- Townsend, K.M.; Frost, A.J.; Lee, C.W.; Papadimitriou, J.M.; Dawkins, H.J. Development of PCR Assays for Species- and Type-Specific Identification of Pasteurella Multocida Isolates. J. Clin. Microbiol. 1998, 36, 1096–1100. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, A.M.; Bennett, M.D. Development of Loop-Mediated Isothermal Amplification-Based Diagnostic Assays for Detection of Pasteurella Multocida and Hemorrhagic Septicemia-Associated P Multocida Serotype B:2. Am. J. Vet. Res. 2017, 78, 134–143. [Google Scholar] [CrossRef]
- Zhao, G.; He, H.; Wang, H. Use of a Recombinase Polymerase Amplification Commercial Kit for Rapid Visual Detection of Pasteurella Multocida. BMC Vet. Res. 2019, 15, 154. [Google Scholar] [CrossRef]
- Bote, Y.; Legesse, K.; Tassew, A.; Zeit, E.D. Isolation, Identification and Antimicrobial Susceptibility of Pasteurella Multocida from Cattle with Hemorrhagic Septicemia in Assosa and Bambasi Districts, Benishangul Gumuz Regional State, Ethiopia. Int. J. Anim. Res. 2017, 1, 67–75. [Google Scholar] [CrossRef][Green Version]
- Cuevas, I.; Carbonero, A.; Cano-Terriza, D.; García-Bocanegra, I.; Amaro, M.Á.; Borge, C. Antimicrobial Resistance of Pasteurella Multocida Type B Isolates Associated with Acute Septicemia in Pigs and Cattle in Spain. BMC Vet. Res. 2020, 16, 222. [Google Scholar] [CrossRef]
- Mostaan, S.; Ghasemzadeh, A.; Sardari, S.; Shokrgozar, M.A.; Brujeni, G.N.; Abolhassani, M.; Ehsani, P.; Karam, M.R.A. Pasteurella Multocida Vaccine Candidates: A Systematic Review. Avicenna J. Med. Biotechnol. 2020, 12, 140–147. [Google Scholar]
- Verma, R.; Jaiswal, T. Haemorrhagic Septicaemia Vaccines. Vaccine 1998, 16, 1184–1192. [Google Scholar] [CrossRef]
- Hussain Shah, A.; Kamboh, A.A.; Rajput, N.; Korejo, N.A. Optimization of Physico-chemical Conditions for the Growth of Pasteurella multocida under In Vitro. J. Agric. Soc. Sci. 2008, 4, 176–179. [Google Scholar]
- Varshney, R.; Varshney, R.; Chaturvedi, V.K.; Rawat, M.; Saminathan, M.; Singh, V.; Singh, R.; Sahoo, M.; Gupta, P.K. De-Velopment of Novel Iron-Regulated Pasteurella Multocida B: 2 Bacterin and Refinement of Vaccine Quality in Terms of Minimum Variation in Particle Size and Distribution Vis-a-Vis Critical Level of Iron in Media. Microb. Pathog. 2020, 147, 1–9. [Google Scholar] [CrossRef]
- Gulia, D.; Aly, S.S. Modeling Vaccination Programs in Outbreaks of Hemorrhagic Septicemia in India. J. Anim. Res. 2020, 10, 19–24. [Google Scholar]
- Yap, S.K.; Zakaria, Z.; Othman, S.S.; Omar, A.R. In Vitro Treatment of Lipopolysaccharide Increases Invasion of Pasteurella Mul-Tocida Serotype B:2 into Bovine Aortic Endothelial Cells. J. Vet. Sci. 2018, 19, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Puspitasari, Y.; Annas, S.; Adza-Rina, M.N.; Zamri-Saad, M. In-Vitro Phagocytosis and Intracellular Killing of Pasteurella Mul-Tocida B:2 by Phagocytic Cells of Buffaloes. Microb. Pathog. 2019, 131, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Puspitasari, Y.; Salleh, A.; Zamri-Saad, M. Ultrastructural Changes in Endothelial Cells of Buffaloes Following In-Vitro Exposure to Pasteurella Multocida B:2. BMC Vet. Res. 2020, 16, 186. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.; Biewenga, J.; De Graaf, F.K. Vacuolating Cytotoxic Activity of Pasteurella Multocida Causing Haemorrhagic Septicaemia in Buffalo and Cattle. FEMS Microbiol. Lett. 1996, 143, 97–101. [Google Scholar] [CrossRef][Green Version]
- Sawada, T.; Rimler, R.B.; Rhoades, K.R. Hemorrhagic Septicemia: Naturally Acquired Antibodies against Pasteurella Multocida Types B and E in Calves in the United States. Am. J. Vet. Res. 1985, 46, 1247–1250. [Google Scholar]
- Yasin, I.-S.M.; Yusoff, S.M.; Mohd, Z.-S.; Mohd, E.A.W. Efficacy of an Inactivated Recombinant Vaccine Encoding a Fimbrial Protein of Pasteurella Multocida B:2 against Hemorrhagic Septicemia in Goats. Trop. Anim. Health Prod. 2010, 43, 179–187. [Google Scholar] [CrossRef]
- Zamri-Saad, M.; Ernie, Z.A.; Sabri, M.Y. Protective Effect Following Intranasal Exposure of Goats to Live Pasteurella Multocida B:2. Trop. Anim. Health Prod. 2006, 38, 541–546. [Google Scholar] [CrossRef]
- Kharb, S.; Charan, S. Mouse Model of Haemorrhagic Septicaemia: Dissemination and Multiplication of Pasteurella Multocida B:2 in Vital Organs after Intranasal and Subcutaneous Challenge in Mice. Vet. Res. Commun. 2012, 37, 59–63. [Google Scholar] [CrossRef]
- Kumar, B.; Chaturvedi, V.K.; Somrajan, S.R.; Kumar, P.; Sreedevi, R.; Kumar, S.; Kaushik, P. Comparative Immune Response of Purified Native Omph Protein Derived from Pasteurella Multocida P52 and Oil Adjuvant Vaccine against Hemorrhagic Septicemia in Mice. Indian J. Anim. Sci. 2011, 81, 1193–1196. [Google Scholar]
- Qu, Y.; Qiu, Z.; Cao, C.; Lu, Y.; Sun, M.; Liang, C.; Zeng, Z. Pharmacokinetics/Pharmacodynamics of Marbofloxacin in a Pasteurella Multocida Serious Murine Lung Infection Model. BMC Vet. Res. 2015, 11, 194. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, S.; Saxena, H.M. Efficacy of bacteriophage Lysed Pasteurella Marker Vaccine in Laboratory Animal Models with a Novel DIVA for Haemorrhagic Septicaemia. Saudi J. Biol. Sci. 2019, 26, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Nimtrakul, P.; Atthi, R.; Limpeanchob, N.; Tiyaboonchai, W. Development of Pasteurella Multocida-Loaded Microparticles for Hemorrhagic Septicemia Vaccine. Drug Dev. Ind. Pharm. 2013, 41, 423–429. [Google Scholar] [CrossRef]
- Ramdani, H.J.S.D.; Johnson, R.B.; Spencer, T.L.; Adler, B.; Dawkins, H. Pasteurella Multocida Infections in Mice with Reference to Haemorrhagic Septicaemia in Cattle snd Buffalo. Immunol. Cell Biol. 1990, 68, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Shivachandra, S.B.; Yogisharadhya, R.; Kumar, A.; Mohanty, N.N.; Nagaleekar, V.K. Recombinant Transferrin Binding Protein A (Rtbpa) Fragments of Pasteurella Multocida Serogroup B:2 Provide Variable Protection Following Homologous Challenge in Mouse Model. Res. Vet. Sci. 2015, 98, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Verma, R.; Pandit, R.S.; Tarfain, N.U. Host Responses and Bacterial Colonization Following Inoculation of Pasteurella Multocida P52 Strain into Unvaccinated and Aluminum Hydroxide Gel Hemorrhagic Septicemia Vaccinated Mice. Microb. Pathog. 2017, 111, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Tabatabaei, M.; Liu, Z.; Finucane, A.; Parton, R.; Coote, J. Protective Immunity Conferred by Attenuated aroA Derivatives of Pasteurella multocida B:2 Strains in a Mouse Model of Hemorrhagic Septicemia. Infect. Immun. 2002, 70, 3355–3362. [Google Scholar] [CrossRef] [PubMed]
- Chelliah, S.; Velappan, R.D.; Lim, K.T.; Swee, C.W.K.; Rashid, N.N.; Rothan, H.A.; Kabir, N.; Ismail, S. Potential DNA Vaccine for Haemorrhagic Septiceamia Disease. Mol. Biotechnol. 2020, 62, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Hussaini, J.; Nazmul, M.H.M.; Masyitah, N.; Abdullah, M.A.; Ismail, S. Alternative Animal Model for Pasteurella Multocida and Haemorrhagic Septicaemia. Biomed. Res. India 2013, 24, 263–266. [Google Scholar]
- Kang, T.L.; Velappan, R.D.; Kabir, N.; Mohamad, J.; Rashid, N.N.; Ismail, S. The ABA392/pET30a Protein of Pasteurella Mul-Tocida Provoked Mucosal Immunity against HS Disease in a Rat Model. Microb. Pathog. 2019, 128, 90–96. [Google Scholar] [CrossRef]
- Annas, S.; Abubakar, M.S.; Zamri-Saad, M.; Jesse, F.F.A.; Zunita, Z. Pathological Changes in the Respiratory, Gastrointestinal and Urinary Tracts of Buffalo Calves Following Experimental Hemorrhagic Septicemia. Pak. Vet. J. 2015, 35, 430–435. [Google Scholar]
- Hodgson, J.C.; Finucane, A.; Dagleish, M.P.; Ataei, S.; Parton, R.; Coote, J.G. Efficacy of Vaccination of Calves against Hemorrhagic Septicemia with a Live aroA Derivative of Pasteurella multocida B:2 by Two Different Routes of Administration. Infect. Immun. 2005, 73, 1475–1481. [Google Scholar] [CrossRef]
- Muangthai, K.; Tankaew, P.; Varinrak, T.; Uthi, R.; Rojanasthien, S.; Sawada, T.; Sthitmatee, N. Intranasal Immunization with a Recombinant Outer Membrane Protein H Based Haemorrhagic Septicemia Vaccine in Dairy Calves. J. Vet. Med Sci. 2018, 80, 68–76. [Google Scholar] [CrossRef]
- Muenthaisong, A.; Nambooppha, B.; Rittipornlertrak, A.; Tankaew, P.; Varinrak, T.; Muangthai, K.; Atthikanyaphak, K.; Sawada, T.; Sthitmatee, N. An Intranasal Vaccination with a Recombinant Outer Membrane Protein H against Haemorrhagic Sep-ticemia in Swamp Buffaloes. Vet. Med. Int. 2020, 2020, 3548973. [Google Scholar] [CrossRef]
- Tabatabaei, M.; Jula, G.R.M.; Jabbari, A.R.; Esmailzadeh, M. Vaccine Efficacy in Cattle Against Hemorrhagic Septicemia with Live Attenuated Aroa Mutant of Pasteurella Multocida B:2 Strain. J. Cell Anim. Biol. 2007, 1, 062–065. [Google Scholar]
- Namazi, F.; Tafti, A.K. Lumpy Skin Disease, an Emerging Transboundary Viral Disease: A Review. Vet. Med. Sci. 2021, 7, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Hunter, P.; Wallace, D. Lumpy Skin Disease in Southern Africa: A Review of the Disease and Aspects of Control. J. S. Afr. Vet. Assoc. 2001, 72, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Xie, J.; Luo, J.; Shao, R.; Jia, K.; Li, S. Lumpy Skin Disease Outbreaks in China, since 3 August 2019. Transbound. Emerg. Dis. 2021, 68, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Beard, P.M. Lumpy Skin Disease: A Direct Threat to Europe. Vet. Rec. 2016, 178, 557–558. [Google Scholar] [CrossRef]
- Tuppurainen, E.; Alexandrov, T.; Beltran-Alcrudo, D. Lumpy Skin Disease Field Manual—A Manual for Veterinarians. FAO Animal Production and Health Manual No. 20. Rome; Food an Agriculture Organization of the United Nations (FAO): Roma, Italy, 2017; p. 60. [Google Scholar]
- Mulatu, E.; Feyisa, A. Review: Lumpy Skin Disease. J. Vet. Sci. Technol. 2018, 9, 1–8. [Google Scholar] [CrossRef]
- Davies, F.G. Lumpy Skin Disease, an African Capripox Virus Disease of Cattle. Br. Vet. J. 1991, 147, 489–503. [Google Scholar] [CrossRef]
- Kumar, S.M. An Outbreak of Lumpy Skin Disease in a Holstein Dairy Herd in Oman: A Clinical Report. Asian J. Anim. Vet. Adv. 2011, 6, 851–859. [Google Scholar]
- Tageldin, M.H.; Wallace, D.B.; Gerdes, G.H.; Putterill, J.F.; Greyling, R.R.; Phosiwa, M.N.; Al Busaidy, R.M.; Al Ismaaily, S.I. Lumpy Skin Disease of Cattle: An Emerging Problem in the Sultanate of Oman. Trop. Anim. Health Prod. 2014, 46, 241–246. [Google Scholar] [CrossRef]
- Sprygin, A.; Pestova, Y.; Wallace, D.; Tuppurainen, E.; Kononov, A. Transmission of Lumpy Skin Disease Virus: A Short Review. Virus Res. 2019, 269, 197637. [Google Scholar] [CrossRef]
- Calistri, P.; De Clercq, K.; Gubbins, S.; Klement, E.; Stegeman, A.; Abrahantes, J.C.; Marojevic, D.; Antoniou, S.; Broglia, A. Lumpy Skin Disease Epidemiological Report IV: Data Collection and Analysis. EFSA J. 2020, 18, e06010. [Google Scholar] [CrossRef] [PubMed]
- Tuppurainen, E.; Venter, E.; Coetzer, J.; Bell-Sakyi, L. Lumpy Skin Disease: Attempted Propagation in Tick Cell Lines and Presence of Viral DNA in Field Ticks Collected from Naturally-Infected Cattle. Ticks Tick Borne Dis. 2015, 6, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Sohier, C.; Haegeman, A.; Mostin, L.; De Leeuw, I.; Van Campe, W.; De Vleeschauwer, A.; Tuppurainen, E.S.M.; Berg, T.V.D.; De Regge, N.; De Clercq, K. Experimental Evidence of Mechanical Lumpy Skin Disease Virus Transmission by Stomoxys Calcitrans Biting Flies and Haematopota spp. Horseflies. Sci. Rep. 2019, 9, 20076. [Google Scholar] [CrossRef] [PubMed]
- Lubinga, J.C.; Tuppurainen, E.S.M.; Mahlare, R.; Coetzer, J.A.W.; Stoltsz, W.H.; Venter, E.H. Evidence of Transstadial and Mechanical Transmission of Lumpy Skin Disease Virus byAmblyomma hebraeumTicks. Transbound. Emerg. Dis. 2013, 62, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Lubinga, J.C.; Tuppurainen, E.S.M.; Coetzer, J.A.W.; Stoltsz, W.H.; Venter, E.H. Transovarial Passage and Transmission of LSDV by Amblyomma Hebraeum, Rhipicephalus Appendiculatus and Rhipicephalus Decoloratus. Exp. Appl. Acarol. 2014, 62, 67–75. [Google Scholar] [CrossRef]
- Lubinga, J.C.; Tuppurainen, E.S.M.; Coetzer, J.A.W.; Stoltsz, W.H.; Venter, E.H. Evidence of Lumpy Skin Disease Virus Over-Wintering by Transstadial Persistence in Amblyomma Hebraeum and Transovarial Persistence in Rhipicephalus Decoloratus Ticks. Exp. Appl. Acarol. 2013, 62, 77–90. [Google Scholar] [CrossRef]
- Aleksandr, K.; Olga, B.; David, W.B.; Pavel, P.; Yana, P.; Svetlana, K.; Alexander, N.; Vladimir, R.; Dmitriy, L.; Alexander, S. Non-Vector-Borne Transmission of Lumpy Skin Disease Virus. Sci. Rep. 2020, 10, 7436. [Google Scholar] [CrossRef]
- Irons, P.; Tuppurainen, E.; Venter, E. Excretion of Lumpy Skin Disease Virus in Bull Semen. Theriogenology 2005, 63, 1290–1297. [Google Scholar] [CrossRef] [PubMed]
- Kononov, A.; Prutnikov, P.; Shumilova, I.; Kononova, S.; Nesterov, A.; Byadovskaya, O.; Pestova, Y.; Diev, V.; Sprygin, A. Determination of Lumpy Skin Disease Virus in Bovine Meat and Offal Products Following Experimental Infection. Transbound. Emerg. Dis. 2019, 66, 1332–1340. [Google Scholar] [CrossRef]
- Annandale, C.; Cornelius, H.; Holm, D.E.; Ebersohn, K.; Venter, E.H. Seminal Transmission of Lumpy Skin Disease Virus in Heifers. Transbound. Emerg. Dis. 2013, 61, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Gari, G.; Waret-Szkuta, A.; Grosbois, V.; Jacquiet, P.; Roger, F. Risk Factors Associated with Observed Clinical Lumpy Skin Disease in Ethiopia. Epidemiol. Infect. 2010, 138, 1657–1666. [Google Scholar] [CrossRef]
- Abdulqa, H.Y.; Rahman, H.S.; Dyary, H.O.; Othman, H.H. Lumpy Skin Disease. Reprod. Immunol. Open Access 2016, 1, 6. [Google Scholar] [CrossRef]
- Haegeman, A.; De Vleeschauwer, A.; De Leeuw, I.; Vidanović, D.; Šekler, M.; Petrović, T.; Demarez, C.; Lefebvre, D.; De Clercq, K. Overview of Diagnostic Tools for Capripox Virus Infections. Prev. Vet. Med. 2020, 181, 104704. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, J.; Boumart, Z.; Daouam, S.; El Arkam, A.; Bamouh, Z.; Jazouli, M.; Tadlaoui, K.O.; Fihri, O.F.; Gavrilov, B.; El Harrak, M. Development and Evaluation of an Inactivated Lumpy Skin Disease Vaccine for Cattle. Vet. Microbiol. 2020, 245, 108689. [Google Scholar] [CrossRef] [PubMed]
- Haegeman, A.; De Leeuw, I.; Mostin, L.; Campe, W.; Aerts, L.; Venter, E.; Tuppurainen, E.; Saegerman, C.; De Clercq, K. Comparative Evaluation of Lumpy Skin Disease Virus-Based Live Attenuated Vaccines. Vaccines 2021, 9, 473. [Google Scholar] [CrossRef]
- Alkhamis, M.A.; VanderWaal, K. Spatial and Temporal Epidemiology of Lumpy Skin Disease in the Middle East, 2012–2015. Front. Vet. Sci. 2016, 3, 19. [Google Scholar] [CrossRef]
- European Food Safety. Lumpy Skin Disease II. Data Collection and Analysis. EFSA J. 2018, 16, e05176. [Google Scholar]
- Gubbins, S.; Stegeman, A.; Klement, E.; Pite, L.; Broglia, A.; Abrahantes, J.C. Inferences about the Transmission of Lumpy Skin Disease Virus between Herds from Outbreaks in Albania in 2016. Prev. Vet. Med. 2020, 181, 104602. [Google Scholar] [CrossRef]
- Kahana-Sutin, E.; Klement, E.; Lensky, I.; Gottlieb, Y. High Relative Abundance of the Stable Fly Stomoxys Calcitrans Is Associated with Lumpy Skin Disease Outbreaks in Israeli Dairy Farms. Med. Vet. Entomol. 2016, 31, 150–160. [Google Scholar] [CrossRef]
- Saegerman, C.; Bertagnoli, S.; Meyer, G.; Ganière, J.-P.; Caufour, P.; De Clercq, K.; Jacquiet, P.; Fournié, G.; Hautefeuille, C.; Etore, F.; et al. Risk of Introduction of Lumpy Skin Disease in France by the Import of Vectors in Animal Trucks. PLoS ONE 2018, 13, e0198506. [Google Scholar] [CrossRef]
- Saegerman, C.; Bertagnoli, S.; Meyer, G.; Ganière, J.-P.; Caufour, P.; De Clercq, K.; Jacquiet, P.; Hautefeuille, C.; Etore, F.; Casal, J.; et al. Risk of introduction of Lumpy Skin Disease into France through Imports of Cattle. Transbound. Emerg. Dis. 2018, 66, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Machado, G.; Korennoy, F.; Alvarez, J.; Picasso-Risso, C.; Perez, A.; VanderWaal, K. Mapping Changes in the Spatiotemporal Distribution of Lumpy Skin Disease Virus. Transbound. Emerg. Dis. 2019, 66, 2045–2057. [Google Scholar] [CrossRef] [PubMed]
- Allepuz, A.; Casal, J.; Beltrán-Alcrudo, D. Spatial Analysis of Lumpy Skin Disease in Eurasia—Predicting Areas at Risk for Further Spread within the Region. Transbound. Emerg. Dis. 2018, 66, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Magori-Cohen, R.; Louzoun, Y.; Herziger, Y.; Oron, E.; Arazi, A.; Tuppurainen, E.; Shpigel, N.Y.; Klement, E. Mathematical Modelling and Evaluation of the Different Routes of Transmission of Lumpy Skin Disease Virus. Vet. Res. 2012, 43, 1. [Google Scholar] [CrossRef]
- Ardestani, E.G.; Mokhtari, A. Modeling the Lumpy Skin Disease Risk Probability in Central Zagros Mountains of Iran. Prev. Vet. Med. 2020, 176, 104887. [Google Scholar] [CrossRef] [PubMed]
- Kononova, S.; Kononov, A.; Shumilova, I.; Byadovskaya, O.; Nesterov, A.; Prutnikov, P.; Babiuk, S.; Sprygin, A. A Lumpy Skin Disease Virus which Underwent a Recombination Event Demonstrates More Aggressive Growth in Primary Cells and Cattle than the Classical Field Isolate. Transbound. Emerg. Dis. 2021, 68, 1377–1383. [Google Scholar] [CrossRef]
- Wolff, J.; Krstevski, K.; Beer, M.; Hoffmann, B. Minimum Infective Dose of a Lumpy Skin Disease Virus Field Strain from North Macedonia. Viruses 2020, 12, 768. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Bernardo, B.; Haga, I.R.; Wijesiriwardana, N.; Hawes, P.C.; Simpson, J.; Morrison, L.R.; MacIntyre, N.; Brocchi, E.; Atkinson, J.; Haegeman, A.; et al. Lumpy Skin Disease Is Characterized by Severe Multifocal Dermatitis with Necrotizing Fibrinoid Vasculitis Following Experimental Infection. Vet. Pathol. 2020, 57, 388–396. [Google Scholar] [CrossRef]
- Munyanduki, H.; Douglass, N.; Offerman, K.; Carulei, O.; Williamson, A.-L. Influence of the Lumpy Skin Disease Virus (LSDV) Superoxide Dismutase Homologue on Host Transcriptional Activity, Apoptosis and Histopathology. J. Gen. Virol. 2020, 101, 645–650. [Google Scholar] [CrossRef]
- Van Boheemen, S.; de Graaf, M.; Lauber, C.; Bestebroer, T.M.; Raj, V.S.; Zaki, A.M.; Osterhaus, A.D.M.E.; Haagmans, B.L.; Gorbalenya, A.E.; Snijder, E.J.; et al. Genomic Characterization of a Newly Discovered Coronavirus Associated with Acute Respiratory Distress Syndrome in Humans. mBio 2012, 3, 00473–00512. [Google Scholar] [CrossRef]
- Zaki, A.; Van Boheemen, S.; Bestebroer, T.; Osterhaus, A.; Fouchier, R. Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, N.; Shaib, H. Middle East. Respiratory Syndrome Coronavirus (MERS-CoV): A Review. Germs 2019, 9, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Widagdo, W.; Okba, N.; Raj, V.S.; Haagmans, B.L. MERS-Coronavirus: From Discovery to Intervention. One Health 2017, 3, 11–16. [Google Scholar] [CrossRef]
- Azhar, E.I.; El-Kafrawy, S.A.; Farraj, S.A.; Hassan, A.M.; Al-Saeed, M.S.; Hashem, A.M.; Madani, T.A. Evidence for Cam-el-to-Human Transmission of MERS Coronavirus. N. Engl. J. Med. 2014, 370, 2499–2505. [Google Scholar] [CrossRef] [PubMed]
- Dighe, A.; Jombart, T.; van Kerkhove, M.D.; Ferguson, N. A Systematic Review of MERS-Cov Seroprevalence and RNA Preva-Lence in Dromedary Camels: Implications for Animal Vaccination. Epidemics 2019, 29, 100350. [Google Scholar] [CrossRef] [PubMed]
- Haagmans, B.L.; van den Brand, J.M.A.; Raj, V.S.; Volz, A.; Wohlsein, P.; Smits, S.L.; Schipper, D.; Bestebroer, T.M.; Okba, N.; Fux, R.; et al. An Orthopoxvirus-Based Vaccine Reduces Virus Excretion After MERS-Cov Infection in Dromedary Camels. Science 2016, 351, 77. [Google Scholar] [CrossRef]
- Munster, V.; De Wit, E.; Feldmann, H. Pneumonia from Human Coronavirus in a Macaque Model. N. Engl. J. Med. 2013, 368, 1560–1562. [Google Scholar] [CrossRef]
- Yao, Y.; Bao, L.; Deng, W.; Xu, L.; Li, F.; Lv, Q.; Yu, P.; Chen, T.; Xu, Y.; Zhu, H.; et al. An Animal Model of MERS Produced by Infection of Rhesus Macaques with MERS Coronavirus. J. Infect. Dis. 2014, 209, 236–242. [Google Scholar] [CrossRef]
- De Wit, E.; Prescott, J.; Baseler, L.; Bushmaker, T.; Thomas, T.; Lackemeyer, M.G.; Martellaro, C.; Milne-Price, S.; Haddock, E.; Haagmans, B.L.; et al. The Middle East. Respiratory Syndrome Coronavirus (MERS-CoV) Does Not. Replicate in Syrian Hamsters. PLoS ONE 2013, 8, e69127. [Google Scholar] [CrossRef]
- Jaiswal, N.K.; Saxena, S.K. Classical Coronaviruses. In Coronavirus Disease 2019 (COVID-19): Epidemiology, Pathogenesis, Diagnosis, and Therapeutics; Saxena, S.K., Ed.; Springer: Singapore, 2020; pp. 141–150. [Google Scholar]
- Liya, G.; Yuguang, W.; Jian, L.; Huaiping, Y.; Xue, H.; Jianwei, H.; Jiaju, M.; Youran, L.; Chen, M.; Yiqing, J. Studies on Viral Pneumonia Related to Novel Coronavirus SARS-CoV-2, SARS-CoV, and MERS-CoV: A Literature Review. APMIS 2020, 128, 423–432. [Google Scholar] [CrossRef]
- Xie, M.; Chen, Q. Insight into 2019 Novel Coronavirus—An Updated Interim Review and Lessons from SARS-CoV and MERS-CoV. Int. J. Infect. Dis. 2020, 94, 119–124. [Google Scholar] [CrossRef]
- Yan, Y.; Chang, L.; Wang, L. Laboratory Testing of SARS-CoV, MERS-CoV, and SARS-CoV-2 (2019-nCoV): Current Status, Challenges, and Countermeasures. Rev. Med. Virol. 2020, 30, e2106. [Google Scholar] [CrossRef]
- Alexander, D.J.; Bell, J.G.; Alders, R.G. A Technology Review: Newcastle Disease—With Special Emphasis on Its Effects on Village Chickens; Food and Agriculture Organization of the United Nations: Rome, Italy, 2004; pp. 1–63. [Google Scholar]
- Alexander, D.J. Newcastle Disease and Other Avian Paramyxovirus. Rev. Sci. Tech. 2000, 19, 443–462. [Google Scholar] [CrossRef]
- Alexander, D.J. Gordon Memorial Lecture. Newcastle Disease. Br. Poult. Sci. 2001, 42, 5–22. [Google Scholar] [CrossRef]
- Bello, M.B.; Yusoff, K.M.; Ideris, A.; Hair-Bejo, M.; Peeters, B.P.H.; Jibril, A.H.; Tambuwal, F.M.; Omar, A.R. Genotype Diversity of Newcastle Disease Virus in Nigeria: Disease Control Challenges and Future Outlook. Adv. Virol. 2018, 2018, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.J. Ecology and Epidemiology of Newcastle Disease; Springer: Milan, Italy, 2009; pp. 19–26. [Google Scholar]
- Jadhav, A.; Zhao, L.; Ledda, A.; Liu, W.; Ding, C.; Nair, V.; Ferretti, L. Patterns of RNA Editing in Newcastle Disease Virus Infections. Viruses 2020, 12, 1249. [Google Scholar] [CrossRef]
- Mayers, J.; Mansfield, K.L.; Brown, I.H. The Role of Vaccination in Risk Mitigation and Control of Newcastle Disease in Poultry. Vaccine 2017, 35, 5974–5980. [Google Scholar] [CrossRef]
- Dimitrov, K.M.; Ramey, A.; Qiu, X.; Bahl, J.; Afonso, C.L. Temporal, Geographic, and Host Distribution of Avian Paramyxovirus 1 (Newcastle Disease Virus). Infect. Genet. Evol. 2016, 39, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.M.; Plemper, R.K. Structure and Organization of Paramyxovirus Particles. Curr. Opin. Virol. 2017, 24, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Nooruzzaman, M.; Mumu, T.T.; Kabiraj, C.K.; Hasnat, A.; Rahman, M.M.; Chowdhury, E.H.; Dimitrov, K.M.; Islam, M.R. Genetic and Biological Characterization of Newcastle Disease Viruses Circulating in Bangladesh During 2010–2017: Further Genetic Di-Versification of Class II Genotype XIII in Southcentral Asia. J. Gen. Virol. 2021, 102, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Alamian, A.; A Pourbakhsh, S.; Shoushtari, A.; Keivanfar, H. Seroprevalence Investigation of Newcastle Disease in Rural Poultries of the Northern Provinces (Golestan, Gilan, and Mazandaran) of Iran. Arch. Razi. Inst. 2019, 74, 365–373. [Google Scholar] [PubMed]
- Alsahami, A.; Ideris, A.; Omar, A.; Ramanoon, S.Z.; Sadiq, M.B. Seroprevalence of Newcastle Disease Virus in Backyard Chickens and Herd-Level Risk Factors of Newcastle Disease in Poultry Farms in Oman. Int. J. Vet. Sci. Med. 2018, 6, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Nwanta, J.; Abdu, P.; Ezema, W. Epidemiology, Challenges and Prospects for Control of Newcastle Disease in Village Poultry in Nigeria. World Poult. Sci. J. 2008, 64, 119–127. [Google Scholar] [CrossRef]
- Terfa, Z.; Garikipati, S.; Kassie, G.; Bettridge, J.; Christley, R. Eliciting Preferences for Attributes of Newcastle Disease Vaccination Programmes for Village Poultry in Ethiopia. Prev. Vet. Med. 2018, 158, 146–151. [Google Scholar] [CrossRef]
- De Bruyn, J.; Thomson, P.C.; Bagnol, B.; Maulaga, W.; Rukambile, E.; Alders, R.G. The Chicken or the Egg? Exploring Bi-Directional Associations between Newcastle Disease Vaccination and Village Chicken Flock Size in Rural Tanzania. PLoS ONE 2017, 12, e0188230. [Google Scholar] [CrossRef]
- Ipara, B.O.; Otieno, D.J.; Nyikal, R.; Makokha, N.S. The Contribution of Extensive Chicken Production Systems and Practices to Newcastle Disease Outbreaks in Kenya. Trop. Anim. Health Prod. 2021, 53, 1–13. [Google Scholar] [CrossRef]
- Hugo, A.; Makinde, O.D.; Kumar, S.; Chibwana, F.F. Optimal Control and Cost Effectiveness Analysis for Newcastle Disease Eco-Epidemiological Model in Tanzania. J. Biol. Dyn. 2016, 11, 190–209. [Google Scholar] [CrossRef]
- Habibi, H.; Firuzi, S.; Nili, H.; Asasi, K.; Mosleh, N. Efficacy of Thermostable Newcastle Disease Virus Strain I-2 in Broiler Chickens Challenged with Highly Virulent Newcastle Virus. Arch. Razi. Inst. 2020, 75, 31–37. [Google Scholar]
- Taylor, T.L.; Miller, P.J.; Olivier, T.L.; Montiel, E.; Garcia, S.C.; Dimitrov, K.M.; Williams-Coplin, D.; Afonso, C.L. Repeated Challenge with Virulent Newcastle Disease Virus Does Not Decrease the Efficacy of Vaccines. Avian Dis. 2017, 61, 245–249. [Google Scholar] [CrossRef]
- Osman, N.; Goovaerts, D.; Sultan, S.; Salt, J.; Grund, C. Vaccine Quality Is a Key Factor to Determine Thermal Stability of Commercial Newcastle Disease (ND)Vaccines. Vaccines 2021, 9, 363. [Google Scholar] [CrossRef]
- Mahmood, M.S.; Sabir, R. Preparation and Evaluation of Avian Influenza (H9) and Newcastle Disease (Thermostable I-2 Strain) Bivalent Vaccine for Commercial Poultry. Agrobiol. Rec. 2021, 3, 17–23. [Google Scholar]
- Zhao, Y.; Liu, H.; Cong, F.; Wu, W.; Zhao, R.; Kong, X. Phosphoprotein Contributes to the Thermostability of Newcastle Disease Virus. BioMed Res. Int. 2018, 2018, 1–6. [Google Scholar] [CrossRef]
- Peebles, E. In ovo Applications on Poultry: A review. Poult. Sci. 2018, 97, 2322–2338. [Google Scholar] [CrossRef] [PubMed]
- Ike, A.C.; Ononugbo, C.M.; Obi, O.J.; Onu, C.J.; Olovo, C.V.; Muo, S.O.; Chukwu, O.S.; Reward, E.E.; Omeke, O.P. Towards Improved Use of Vaccination in the Control of Infectious Bronchitis and Newcastle Disease in Poultry: Understanding the Immunological Mechanisms. Vaccines 2021, 9, 20. [Google Scholar] [CrossRef] [PubMed]
- Cvetić, Ž.; Nedeljković, G.; Jergović, M.; Bendelja, K.; Mazija, H.; Gottstein, Ž. Immunogenicity of Newcastle Disease Virus Strain ZG1999HDS Applied Oculonasally or by Means of Nebulization to Day-Old Chicks. Poult. Sci. 2021, 100, 101001. [Google Scholar] [CrossRef] [PubMed]
- Shittu, I.; Zhu, Z.; Lu, Y.; Hutcheson, J.M.; Stice, S.L.; West, F.D.; Donadeu, M.; Dungu, B.; Fadly, A.M.; Zavala, G.; et al. Development, Characterization and Optimization of a New Suspension Chicken-Induced Pluripotent Cell Line for the Production of Newcastle Disease Vaccine. Biologicals 2016, 44, 24–32. [Google Scholar] [CrossRef]
- Shahar, E.; Haddas, R.; Goldenberg, D.; Lublin, A.; Bloch, I.; Hinenzon, N.B.; Pitcovski, J. Newcastle Disease Virus: Is an Updated Attenuated Vaccine Needed? Avian Pathol. 2018, 47, 467–478. [Google Scholar] [CrossRef]
- Ferreira, H.; Miller, P.; Suarez, D. Protection against Different Genotypes of Newcastle Disease Viruses (NDV) Afforded by an Adenovirus-Vectored Fusion Protein and Live NDV Vaccines in Chickens. Vaccines 2021, 9, 182. [Google Scholar] [CrossRef]
- Aldous, E.; Alexander, D. Newcastle Disease in Pheasants (Phasianus Colchicus): A Review. Vet. J. 2008, 175, 181–185. [Google Scholar] [CrossRef]
- Alexander, D.J.; Alexander, D.J. Newcastle Disease in Ostriches (Struthio Camelus)—A Review. Avian Pathol. 2000, 29, 95–100. [Google Scholar] [CrossRef]
- Kuiken, T.; Frandsen, D.; Clavijo, A. Newcastle Disease in Cormorants. Can. Vet. J. 1998, 39, 299. [Google Scholar]
- Samour, J. Newcastle Disease in Captive Falcons in the Middle East: A Review of Clinical and Pathologic Findings. J. Avian Med. Surg. 2014, 28, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Brown, V.R.; Bevins, S.N. A Review of Virulent Newcastle Disease Viruses in the United States and the Role of Wild Birds in Viral Persistence and Spread. Vet. Res. 2017, 48, 68. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Sun, L.; Sun, X.; Li, S.; Zhang, W.; Pulscher, L.A.; Chai, H.; Xing, M. Newcastle Disease Virus from Domestic Mink, China, 2014. Vet. Microbiol. 2017, 198, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Elmberg, J.; Berg, C.; Lerner, H.; Waldenström, J.; Hessel, R. Potential Disease Transmission from Wild Geese and Swans to Livestock, Poultry and Humans: A Review of the Scientific Literature from a One Health Perspective. Infect. Ecol. Epidemiol. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Rehan, M.; Aslam, A.; Khan, M.-U.-R.; Abid, M.; Hussain, S.; Umber, J.; Anjum, A.; Hussain, A. Potential Economic Impact of Newcastle Disease Virus Isolated from Wild Birds on Commercial Poultry Industry of Pakistan: A Review. Hosts Viruses 2019, 6, 1–15. [Google Scholar] [CrossRef]
- Bello, M.B.; Yusoff, K.; Ideris, A.; Hair-Bejo, M.; Jibril, A.H.; Peeters, B.P.H.; Omar, A.R. Exploring the Prospects of Engineered Newcastle Disease Virus in Modern Vaccinology. Viruses 2020, 12, 451. [Google Scholar] [CrossRef]
- Lundstrom, K. RNA Viruses as Tools in Gene Therapy and Vaccine Development. Genes 2019, 10, 189. [Google Scholar] [CrossRef]
- Vijayakumar, G.; McCroskery, S.; Palese, P. Engineering Newcastle Disease Virus as an Oncolytic Vector for Intratumoral De-livery of Immune Checkpoint Inhibitors and Immunocytokines. J. Virol. 2020, 94, 1–14. [Google Scholar] [CrossRef]
- Choi, K.-S. Newcastle Disease Virus Vectored Vaccines as Bivalent or Antigen Delivery Vaccines. Clin. Exp. Vaccine Res. 2017, 6, 72–82. [Google Scholar] [CrossRef]
- Chen, X.; Yang, J.; Ji, Y.; Okoth, E.; Liu, B.; Li, X.; Yin, H.; Zhu, Q. Recombinant Newcastle Disease Virus Expressing African Swine Fever Virus Protein 72 Is Safe and Immunogenic in Mice. Virol. Sin. 2016, 31, 150–159. [Google Scholar] [CrossRef]
- Ch’Ng, W.-C.; Stanbridge, E.J.; Ong, K.-C.; Wong, K.-T.; Yusoff, K.; Shafee, N. Partial Protection against Enterovirus 71 (EV71) Infection in a Mouse Model Immunized with Recombinant Newcastle Disease Virus Capsids Displaying the EV71 VP1 Fragment. J. Med. Virol. 2011, 83, 1783–1791. [Google Scholar] [CrossRef] [PubMed]
- Debnath, A.; Pathak, D.C.; D’Silva, A.L.; Batheja, R.; Ramamurthy, N.; Vakharia, V.N.; Chellappa, M.M.; Dey, S. Newcastle Disease Virus Vectored Rabies Vaccine Induces Strong Humoral and Cell Mediated Immune Responses in Mice. Vet. Microbiol. 2020, 251, 108890. [Google Scholar] [CrossRef] [PubMed]
- Manoharan, V.K.; Khattar, S.K.; LaBranche, C.C.; Montefiori, D.C.; Samal, S.K. Modified Newcastle Disease Virus as an Im-Proved Vaccine Vector against Simian Immunodeficiency Virus. Sci. Rep. 2018, 8, 8952. [Google Scholar] [CrossRef] [PubMed]
- Shirvani, E.; Samal, S.K. Newcastle Disease Virus as a Vaccine Vector for SARS-CoV-2. Pathogens 2020, 9, 619. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; McCroskery, S.; Liu, W.C.; Leist, S.R.; Liu, Y.; Albrecht, R.A.; Slamanig, S.; Oliva, J.; Amanat, F.; Schäfer, A.; et al. A Newcastle Disease Virus (NDV) Expressing a Mem-brane-Anchored Spike as a Cost-Effective Inactivated SARS-CoV-2 Vaccine. Vaccines 2020, 8, 771. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Shi, W.; Abiona, O.M.; Nazzari, A.; Olia, A.S.; Ou, L.; Phung, E.; Stephens, T.; Tsybovsky, Y.; Verardi, R.; et al. Newcastle Disease Virus-Like Particles Displaying Prefusion-Stabilized SARS-CoV-2 Spikes Elicit Potent Neutralizing Responses. Vaccines 2021, 9, 73. [Google Scholar] [CrossRef]
- Schirrmacher, V. Immunobiology of Newcastle Disease Virus and Its Use for Prophylactic Vaccination in Poultry and as Adjuvant for Therapeutic Vaccination in Cancer Patients. Int. J. Mol. Sci. 2017, 18, 1103. [Google Scholar] [CrossRef]
- Vijayakumar, G.; Zamarin, D. Design and Production of Newcastle Disease Virus for Intratumoral Immunomodulation. Methods Mol. Biol. 2019, 2058, 133–154. [Google Scholar] [CrossRef]
- Burman, B.; Pesci, G.; Zamarin, D. Newcastle Disease Virus at the Forefront of Cancer Immunotherapy. Cancers 2020, 12, 3552. [Google Scholar] [CrossRef]
- Meng, Q.; He, J.; Zhong, L.; Zhao, Y. Advances in the Study of Antitumour Immunotherapy for Newcastle Disease Virus. Int. J. Med. Sci. 2021, 18, 2294–2302. [Google Scholar] [CrossRef]
- Yurchenko, K.S.; Zhou, P.; Kovner, A.V.; Zavjalov, E.L.; Shestopalova, L.V.; Shestopalov, A.M. Oncolytic Effect of Wild-Type Newcastle Disease Virus Isolates in Cancer Cell Lines In Vitro and In Vivo on Xenograft Model. PLoS ONE 2018, 13, e0195425. [Google Scholar] [CrossRef]
- Amin, Z.M.; Ani, M.A.C.; Tan, S.W.; Yeap, S.K.; Alitheen, N.B.; Najmuddin, S.U.F.S.; Kalyanasundram, J.; Chan, S.C.; Veerakumarasivam, A.; Chia, S.L.; et al. Evaluation of a Recombinant Newcastle Disease Virus Expressing Human IL12 against Human Breast Cancer. Sci. Rep. 2019, 9, 13999. [Google Scholar] [CrossRef]
- Li, X.; Chai, T.; Wang, Z.; Song, C.; Cao, H.; Liu, J.; Zhang, X.; Wang, W.; Yao, M.; Miao, Z. Occurrence and Transmission of Newcastle Disease Virus Aerosol Originating from Infected Chickens under Experimental Conditions. Vet. Microbiol. 2009, 136, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Soliman, M.A.; Nour, A.A.; Erfan, A.M. Quantitative Evaluation of Viral Interference among Egyptian Isolates of Highly Pathogenic Avian Influenza Viruses (H5N1 And H5N8) with the Lentogenic and Velogenic Newcastle Disease Virus Genotype VII in Specific Patho-Gen-Free Embryonated Chicken Eggs Model. Vet. World 2019, 12, 1833–1839. [Google Scholar] [PubMed]
- Carrasco, A.D.O.T.; Seki, M.C.; Benevenute, J.L.; Ikeda, P.; Pinto, A.A. Experimental Infection with Brazilian Newcastle Disease Virus Strain in Pigeons and Chickens. Braz. J. Microbiol. 2016, 47, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Gallili, G.E.; Ben-Nathan, D. Newcastle Disease Vaccines. Biotechnol. Adv. 1998, 16, 343–366. [Google Scholar] [CrossRef]
- Palya, V.; Tatár-Kis, T.; Arafa, A.S.A.; Felföldi, B.; Mató, T.; Setta, A. Efficacy of a Turkey Herpesvirus Vectored Newcastle Disease Vaccine against Genotype VII.1.1 Virus: Challenge Route Affects Shedding Pattern. Vaccines 2021, 9, 37. [Google Scholar] [CrossRef]
- Rehman, Z.; Ren, S.; Butt, S.; Manzoor, Z.; Iqbal, J.; Anwar, M.; Sun, Y.; Qiu, X.; Tan, L.; Liao, Y.; et al. Newcastle Disease Virus Induced Pathologies Severely Affect the Exocrine and Endocrine Functions of the Pancreas in Chickens. Genes 2021, 12, 495. [Google Scholar] [CrossRef]
- Lardinois, A.; Berg, T.V.D.; Lambrecht, B.; Steensels, M. A Model for the Transfer of Passive Immunity against Newcastle Disease and Avian Influenza in Specific Pathogen Free Chickens. Avian Pathol. 2014, 43, 118–124. [Google Scholar] [CrossRef]
- Susta, L.; Segovia, D.; Olivier, T.L.; Dimitrov, K.M.; Shittu, I.; Marcano, V.; Miller, P.J. Newcastle Disease Virus Infection in Quail. Vet. Pathol. 2018, 55, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, K.M.; Ferreira, H.L.; Pantin-Jackwood, M.J.; Taylor, T.L.; Goraichuk, I.V.; Crossley, B.M.; Killian, M.L.; Bergeson, N.H.; Torchetti, M.K.; Afonso, C.L.; et al. Pathogenicity and Transmission of Virulent Newcastle Disease Virus from the 2018–2019 California Outbreak and Related Viruses in Young and Adult Chickens. Virology 2019, 531, 203–218. [Google Scholar] [CrossRef]
- Schilling, M.A.; Katani, R.; Memari, S.; Cavanaugh, M.; Buza, J.; Radzio-Basu, J.; Mpenda, F.N.; Deist, M.S.; Lamont, S.J.; Kapur, V. Transcriptional Innate Immune Response of the Developing Chicken Embryo to Newcastle Disease Virus Infection. Front. Genet. 2018, 9, 61. [Google Scholar] [CrossRef]
- Miller, P.J.; Kim, L.M.; Ip, H.; Afonso, C.L. Evolutionary Dynamics of Newcastle Disease Virus. Virology 2009, 391, 64–72. [Google Scholar] [CrossRef]
- Abd El-Baky, N.; Amara, A.A. Newcastle Disease Virus (Lasota Strain) as a Model for Virus Ghosts Preparation Using H2O2 Bio-Critical Concentration. Int. Sci. Investig. J. 2014, 3, 38–50. [Google Scholar]
- Njeumi, F.; Bailey, D.; Soula, J.J.; Diop, B.; Tekola, B.G. Eradicating the Scourge of Peste des Petits Ruminants from the World. Viruses 2020, 12, 313. [Google Scholar] [CrossRef]
- Albina, E.; Kwiatek, O.; Minet, C.; Lancelot, R.; de Almeida, R.S.; Libeau, G. Peste des Petits Ruminants, the Next Eradicated Animal Disease? Vet. Microbiol. 2013, 165, 38–44. [Google Scholar] [CrossRef]
- Idoga, E.S.; Armson, B.; Alafiatayo, R.; Ogwuche, A.; Mijten, E.; Ekiri, A.B.; Varga, G.; Cook, A.J.C. A Review of the Current Status of Peste des Petits Ruminants Epidemiology in Small Ruminants in Tanzania. Front. Vet. Sci. 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Baron, M.D.; Parida, S.; Oura, C.A.L. Peste des Petits Ruminants: A Suitable Candidate for Eradication? Vet. Rec. 2011, 169, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Kabir, A.; Kalhoro, D.H.; Abro, S.H.; Kalhoro, M.S.; Yousafzai, H.A.; Shams, S.; Khan, I.U.; Lochi, G.M.; Mazari, M.Q.; Baloch, M.W.; et al. Peste des Petits Ruminants: A Review. Pure Appl. Biol. 2019, 8, 1214–1222. [Google Scholar] [CrossRef]
- Torsson, E.; Kgotlele, T.; Berg, M.; Mtui-Malamsha, N.; Swai, E.S.; Wensman, J.J.; Misinzo, G. History and Current Status of Peste des Petits Ruminants Virus in Tanzania. Infect. Ecol. Epidemiol. 2016, 6, 32701. [Google Scholar] [CrossRef] [PubMed]
- Ahaduzzaman, M. Peste des Petits Ruminants (PPR) in Africa and Asia: A Systematic Review and Meta-Analysis of the Prevalence in Sheep and Goats between 1969 and 2018. Vet. Med. Sci. 2020, 6, 813–833. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.U.; Dhama, K.; Ali, Q.; Hussain, I.; Oneeb, M.; Chaudhary, U.; Wensman, J.J.; Shabbir, M.Z. Peste des Petits Ru-Minants in Large Ruminants, Camels and Unusual Hosts. Vet. Q. 2020, 40, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Agga, G.E.; Raboisson, D.; Walch, L.; Alemayehu, F.; Semu, D.T.; Bahiru, G.; Woube, Y.A.; Belihu, K.; Tekola, B.G.; Bekana, M.; et al. Epidemiological Survey of Peste des Petits Ruminants in Ethiopia: Cattle as Potential Sentinel for Sur-veillance. Front. Vet. Sci. 2019, 6, 302. [Google Scholar] [CrossRef]
- Rahman, A.U.; Wensman, J.J.; Abubakar, M.; Shabbir, M.Z.; Rossiter, P. Peste des Petits Ruminants in Wild Ungulates. Trop. Anim. Health Prod. 2018, 50, 1815–1819. [Google Scholar] [CrossRef]
- Schulz, C.; Fast, C.; Wernery, U.; Kinne, J.; Joseph, S.; Schlottau, K.; Jenckel, M.; Höper, D.; Patteril, N.A.G.; Syriac, G.; et al. Camelids and Cattle Are Dead-End Hosts for Peste-des-Petits-Ruminants Virus. Viruses 2019, 11, 1133. [Google Scholar] [CrossRef]
- Nour, H.S.H. Challenges and Opportunities for Global Eradication of Paste des Petits Ruminants (PPR). J. Trop. Dis. Public Health 2020, 8, 1–10. [Google Scholar]
- Zakian, A.; Nouri, M.; Faramarzian, F.; Tehrani-Sharif, M.; Rezaie, A.; Mokhber-Dezfouli, M.R. Comprehensive Review on Peste Des. Petits Ruminants [PPR] Disease in Ruminants and Camels: With Emphasis on Clinical Signs and Histopathological Finding. J. Vet. Sci. Med. Diagn. 2016, 5, 1–8. [Google Scholar] [CrossRef]
- Balamurugan, V.; Hemadri, D.; Gajendragad, M.R.; Singh, R.K.; Rahman, H. Diagnosis and Control of Peste des Petits Ruminants: A Comprehensive Review. Virus Dis. 2013, 25, 39–56. [Google Scholar] [CrossRef]
- Giato, C.G.; SKihu, M.H.; Bebora, L.C.; Njenga, J.M.; Wairire, G.G.; Maingi, N.; Muse, E.A.; Karimuribo, E.D.; Misinzo, G.; Mellau, L.S.B.; et al. Comparison of Peste des Petits Ruminants (PPR) Disease between Tanzania and Kenya. In Proceedings of the 3rd RUFORUM Biennial Conference, Entebbe, Uganda, 24–28 September 2012. [Google Scholar]
- Akanbi, O.B.; Franzke, K.; Adedeji, A.J.; Ulrich, R.; Teifke, J.P. Peste des Petits Ruminants Virus and Goatpox Virus Co-Infection in Goats. Vet. Pathol. 2020, 57, 550–553. [Google Scholar] [CrossRef] [PubMed]
- Adedeji, A.J.; Dashe, Y.; Akanbi, O.B.; Woma, T.Y.; Jambol, A.R.; Adole, J.A.; Bolajoko, M.B.; Chima, N.; Asala, O.; Tekki, I.S.; et al. Co-Infection of Peste des Petits Ruminants and Goatpox in a Mixed Flock of Sheep and Goats in Kanam, North Central Nigeria. Vet. Med. Sci. 2019, 5, 412–418. [Google Scholar] [CrossRef]
- Kinimi, E.; Odongo, S.; Muyldermans, S.; Kock, R.; Misinzo, G. Paradigm Shift in the Diagnosis of Peste des Petits Ruminants: Scoping Review. Acta Vet. Scand. 2020, 62, 7–14. [Google Scholar] [CrossRef]
- Manzoor, S.; Abubakar, M.; Bin Zahur, A.; Yunus, A.W.; Ullah, A.; Afzal, M. Genetic Characterization of Peste des Petits Ruminants Virus (Pakistani Isolates) and Comparative Appraisal of Diagnostic Assays. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Kardjadj, M.; Luka, P.D. Molecular Epidemiology of Foot and Mouth Disease, Bluetongue and Pest des Petits Ruminants in Algeria: Historical Perspective, Diagnosis and Control. Afr. J. Biotechnol. 2016, 15, 2474–2479. [Google Scholar]
- Mahapatra, M.; Howson, E.; Fowler, V.; Batten, C.; Flannery, J.; Selvaraj, M.; Parida, S. Rapid Detection of Peste des Petits Ruminants Virus (PPRV) Nucleic Acid Using a Novel Low-Cost Reverse Transcription Loop-Mediated Isothermal Amplification (RT-LAMP) Assay for Future Use in Nascent PPR Eradication Programme. Viruses 2019, 11, 699. [Google Scholar] [CrossRef] [PubMed]
- Baron, M.; Diallo, A.; Lancelot, R.; Libeau, G. Peste des Petits Ruminants Virus. Adv. Clin. Chem. 2016, 95, 1–42. [Google Scholar] [CrossRef]
- Edo, T.; Deneke, Y.; Abdela, N. Peste Des Petits Ruminants and its Economic Importance. Glob. Vet. 2017, 18, 256–268. [Google Scholar]
- Abubakar, M.; Irfan, M. An Overview of Treatment Options to Combat Peste des Petits Ruminants in Endemic Situations. Res. J. Vet. Pract. 2014, 2, 4–7. [Google Scholar] [CrossRef]
- Baron, M.D.; Diop, B.; Njeumi, F.; Willett, B.J.; Bailey, D. Future research to underpin successful peste des petits ruminants virus (PPRV) eradication. J. Gen. Virol. 2017, 98, 2635–2644. [Google Scholar] [CrossRef] [PubMed]
- Parida, S.; Muniraju, M.; Mahapatra, M.; Muthuchelvan, D.; Buczkowski, H.; Banyard, A. Peste des Petits Ruminants. Vet. Microbiol. 2015, 181, 90–106. [Google Scholar] [CrossRef]
- Kumar, N.; Barua, S.; Riyesh, T.; Tripathi, B.N. Advances in Peste des Petits Ruminants Vaccines. Vet. Microbiol. 2017, 206, 91–101. [Google Scholar] [CrossRef]
- Jia, X.-X.; Wang, H.; Liu, Y.; Meng, D.-M.; Fan, Z.-C.; Hui, W. Development of Vaccines for Prevention of Peste-des-Petits-Ruminants Virus Infection. Microb. Pathog. 2020, 142, 104045. [Google Scholar] [CrossRef]
- Dundon, W.G.; Diallo, A.; Cattoli, G. Peste des Petits Ruminants in Africa: A Review of Currently Available Molecular Epide-Miological Data, 2020. Arch. Virol. 2020, 165, 2147–2163. [Google Scholar] [CrossRef]
- Cameron, A.R. Strategies for the Global Eradication of Peste des Petits Ruminants: An Argument for the Use of Guerrilla Rather Than Trench Warfare. Front. Vet. Sci. 2019, 6, 331. [Google Scholar] [CrossRef] [PubMed]
- Jafari-Gh, A.; RLaven, A.; Eila, N.; Yadi, J.; Hatami, Z.; Soleimani, P.; Jafari-Gh, S.; Lesko, M.M.; Sinafar, M.; Heidari, E. Transboundary and Infectious Diseases of Small Ruminants: Knowledge, Attitude, and Practice of Nomadic and Semi-Nomadic Pastor-Alists in Northern Iran. Small Rumin. Res. 2020, 183, 1–6. [Google Scholar] [CrossRef]
- Acosta, D.; Hendrickx, S.; McKune, S. The Livestock Vaccine Supply Chain: Why It Matters and How it Can Help Eradicate Peste des Petits Ruminants, Based on Findings in Karamoja, Uganda. Vaccine 2019, 37, 6285–6290. [Google Scholar] [CrossRef]
- Bardhan, D.; Kumar, S.; Anandsekaran, G.; Chaudhury, J.K.; Meraj, M.; Singh, R.K.; Verma, M.R.; Kumar, D.; Kumar, P.T.N.; Lone, S.A.; et al. The Economic Impact of Peste des Petits Ruminants in India. Rev. Sci. Tech. 2017, 36, 245–263. [Google Scholar]
- Ma, J.; Gao, X.; Liu, B.; Chen, H.; Xiao, J.; Wang, H. Peste des Petits Ruminants in China: Spatial Risk Analysis. Transbound. Emerg. Dis. 2019, 66, 1784–1788. [Google Scholar] [CrossRef] [PubMed]
- Fournié, G.; Waret-Szkuta, A.; Camacho, A.; Yigezu, L.M.; Pfeiffer, D.; Roger, F. A Dynamic Model of Transmission and Elimination of Peste des Petits Ruminants in Ethiopia. Proc. Natl. Acad. Sci. USA 2018, 115, 8454–8459. [Google Scholar] [CrossRef] [PubMed]
- Assefa, A.; Tibebu, A.; Bihon, A.; Yimana, M. Global Ecological Niche Modelling of Current and Future Distribution of Peste des Petits Ruminants Virus (Pprv) with an Ensemble Modelling Algorithm. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Ruget, A.-S.; Tran, A.; Waret-Szkuta, A.; Moutroifi, Y.O.; Charafouddine, O.; Cardinale, E.; Cêtre-Sossah, C.; Chevalier, V. Spatial Multicriteria Evaluation for Mapping the Risk of Occurrence of Peste des Petits Ruminants in Eastern Africa and the Union of the Comoros. Front. Vet. Sci. 2019, 6, 455. [Google Scholar] [CrossRef]
- Munduganore, D.S.; YMundaganore, D.; Ashokan, K.V. Sequence Analysis of Protein in Peste des. Petits Ruminants: An. In Silico Approach. Int. J. Food Agric. Vet. Sci. 2012, 2, 26–36. [Google Scholar]
- Parida, S.; Selvaraj, M.; Gubbins, S.; Pope, R.; Banyard, A.; Mahapatra, M. Quantifying Levels of Peste des Petits Ruminants (PPR) Virus in Excretions from Experimentally Infected Goats and Its Importance for Nascent PPR Eradication Programme. Viruses 2019, 11, 249. [Google Scholar] [CrossRef] [PubMed]
- Truong, T.; Boshra, H.; Embury-Hyatt, C.; Nfon, C.; Gerdts, V.; Tikoo, S.; Babiuk, L.A.; Kara, P.D.; Chetty, T.; Mather, A.; et al. Peste des Petits Ruminants Virus Tissue Tropism and Pathogenesis in Sheep and Goats following Experimental Infection. PLoS ONE 2014, 9, e87145. [Google Scholar] [CrossRef] [PubMed]
- Bamouh, Z.; Fakri, F.; Jazouli, M.; Safini, N.; Tadlaoui, K.O.; Elharrak, M. Peste des Petits Ruminants Pathogenesis on Experimental Infected Goats by the Moroccan 2015 Isolate. BMC Vet. Res. 2019, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chandran, N.D.J.; Kumanan, K.; Venkatesan, R.A. Differentiation of Peste des Petits Ruminants and Rinderpest Viruses by Neutralisation Indices Using Hyperimmune Rinderpest Antiserum. Trop. Anim. Health Prod. 1995, 27, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Banadyga, L.; Schiffman, Z.; Huang, P.; Li, E.; Gao, Y.; Feng, N.; Xia, X.; Feng, N. Peste des Petits Ruminants Virus-Like Particles Induce a Potent Humoral and Cellular Immune Response in Goats. Viruses 2019, 11, 918. [Google Scholar] [CrossRef] [PubMed]
- Youssef, M.; Ibrahim, H. Preparation of Hyperimmune Serum against Peste des Petits Ruminants to Be Used in Emergency Cases. Suez Canal Vet. Med. J. SCVMJ 2016, 21, 143–152. [Google Scholar] [CrossRef][Green Version]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Changes to Taxonomy and the International Code of Virus Classification and Nomenclature Ratified by the International Committee on Taxonomy of Viruses (2017). Arch. Virol. 2017, 162, 2505–2538. [Google Scholar] [CrossRef]
- Ikegami, T. Molecular Biology and Genetic Diversity of Rift Valley Fever Virus. Antivir. Res. 2012, 95, 293–310. [Google Scholar] [CrossRef]
- Grobbelaar, A.A.; Weyer, J.; Leman, P.A.; Kemp, A.; Paweska, J.T.; Swanepoel, R. Molecular Epidemiology of Rift Valley Fever Virus. Emerg. Infect. Dis. 2011, 17, 2270–2276. [Google Scholar] [CrossRef]
- Nanyingi, M.O.; Munyua, P.; Kiama, S.G.; Muchemi, G.M.; Thumbi, S.M.; Bitek, A.O.; Bett, B.; Muriithi, R.M.; Njenga, M.K. A Systematic Review of Rift Valley Fever Epidemiology 1931–2014. Infect. Ecol. Epidemiol. 2015, 5, 28024. [Google Scholar] [CrossRef]
- Anyamba, A.; Linthicum, K.J.; Small, J.; Britch, S.C.; Pak, E.; de la Rocque, S.; Formenty, P.; Hightower, A.W.; Breiman, R.F.; Chretien, J.-P.; et al. Prediction, Assessment of the Rift Valley Fever Activity in East. and Southern Africa 2006?2008 and Possible Vector Control. Strategies. J. Am. Soc. Trop. Med. Hyg. 2010, 83 (Suppl. S2), 43–51. [Google Scholar] [CrossRef]
- Linthicum, K.J. A Rift Valley Fever Risk Surveillance System for Africa Using Remotely Sensed Data: Potential for Use on Other Continents. Vet. Ital. 2007, 43, 663. [Google Scholar]
- Tantely, L.M.; Boyer, S.; Fontenille, D. A Review of Mosquitoes Associated with Rift Valley Fever Virus in Madagascar. Am. J. Trop. Med. Hyg. 2015, 92, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Pepin, M.; Bouloy, M.; Bird, B.H.; Kemp, A.; Paweska, J. Rift Valley Fever Virus (Bunyaviridae: Phlebovirus): An Update on Pathogenesis, Molecular Epidemiology, Vectors, Diagnostics and Prevention. Vet. Res. 2010, 41, 61. [Google Scholar] [CrossRef]
- Clark, M.H.A.; Warimwe, G.M.; di Nardo, A.; Lyons, N.A.; Gubbins, S. Systematic Literature Review of Rift Valley Fever Virus Seroprevalence in Livestock, Wildlife and Humans in Africa from 1968 to 2016. PLoS Negl. Trop. Dis. 2018, 12, e0006627. [Google Scholar] [CrossRef] [PubMed]
- Bird, B.H.; Ksiazek, T.G.; Nichol, S.T.; Maclachlan, N.J. Rift Valley Fever Virus. J. Am. Vet. Med. Assoc. 2009, 234, 883–893. [Google Scholar] [CrossRef] [PubMed]
- Odendaal, L.; Davis, A.S.; Venter, E.H. Insights into the Pathogenesis of Viral Haemorrhagic Fever Based on Virus Tropism and Tissue Lesions of Natural Rift Valley Fever. Viruses 2021, 13, 709. [Google Scholar] [CrossRef]
- Anyangu, A.S.; Gould, L.H.; Sharif, S.K.; Nguku, P.M.; Omolo, J.O.; Mutonga, D.; Rao, C.Y.; Lederman, E.R.; Schnabel, D.; Paweska, J.T.; et al. Risk Factors for Severe Rift Valley Fever Infection in Kenya, 2007. Am. Soc. Trop. Med. Hyg. 2010, 83 (Suppl. S2), 14–21. [Google Scholar] [CrossRef]
- Lang, Y.; Li, Y.; Jasperson, D.; Henningson, J.; Lee, J.; Ma, J.; Li, Y.; Duff, M.; Liu, H.; Bai, D.; et al. Identification and Evaluation of Antivirals for Rift Valley Fever Virus. Vet. Microbiol. 2019, 230, 110–116. [Google Scholar] [CrossRef]
- Atkins, C.; Freiberg, A.N. Recent Advances in the Development of Antiviral Therapeutics for Rift Valley Fever Virus Infection. Future Virol. 2017, 12, 651–665. [Google Scholar] [CrossRef]
- Faburay, B.; LaBeaud, A.D.; McVey, D.S.; Wilson, W.C.; Richt, J.A. Current Status of Rift Valley Fever Vaccine Development. Vaccines 2017, 5, 29. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, T.; Makino, S. Rift Valley Fever Vaccines. Vaccine 2009, 27 (Suppl. S4), D69–D72. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Pinilla, E.; Marín-López, A.; Moreno, S.; Lorenzo, G.; Trigo, S.U.; Jiménez-Cabello, L.; Benavides, J.; Nogales, A.; Blasco, R.; Brun, A.; et al. A protective bivalent vaccine against Rift Valley fever and bluetongue. npj Vaccines 2020, 5, 70. [Google Scholar] [CrossRef]
- Roeder, P.; Mariner, J.; Kock, R. Rinderpest: The veterinary perspective on eradication. Philos. Trans. R. Soc. B: Biol. Sci. 2013, 368, 20120139. [Google Scholar] [CrossRef] [PubMed]
- Düx, A.; Lequime, S.; Patrono, L.V.; Vrancken, B.; Boral, S.; Gogarten, J.F.; Hilbig, A.; Horst, D.; Merkel, K.; Prepoint, B.; et al. Measles virus and rinderpest virus divergence dated to the sixth century BCE. Science 2020, 368, 1367–1370. [Google Scholar] [CrossRef] [PubMed]
- Roeder, P.L.; Taylor, W.P. Rinderpest. Vet. Clin. N. Am. Food Anim. Pract. 2002, 18, 515–547. [Google Scholar] [CrossRef]
- Tounkara, K.; Nwankpa, N. Rinderpest Experience. Rev. Sci. Tech. 2017, 36, 569–578. [Google Scholar] [CrossRef]
- Njeumi, F.; Taylor, W.; Diallo, A.; Miyagishima, K.; Pastoret, P.-P.; Vallat, B.; Traore, M. The Long Journey: A Brief Review of the Eradication of Rinderpest. Rev. Sci. Tech. 2012, 31, 729–746. [Google Scholar] [CrossRef]
- Barrett, T.; Rossiter, P. Rinderpest: The Disease and Its Impact on Humans and Animals. Adv. Appl. Microbiol. 1999, 53, 89–110. [Google Scholar] [CrossRef]
- Vallat, F. An Outbreak in France in the XVIIIth Century: Rinderpest. C. R. Biol. 2012, 335, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Roeder, P.L. Rinderpest: The End of Cattle Plague. Prev. Vet. Med. 2011, 102, 98–106. [Google Scholar] [CrossRef]
- Carrillo, C.; Prarat, M.; Vagnozzi, A.; Calahan, J.D.; Smoliga, G.; Nelson, W.M.; Rodriguez, L.L. Specific Detection of Rinderpest Virus by Real-Time Reverse Transcription-PCR in Preclinical and Clinical Samples from Experimentally Infected Cattle. J. Clin. Microbiol. 2010, 48, 4094–4101. [Google Scholar] [CrossRef]
- Letter to Mr Urban. In The Gentleman’s Magazine; Henry and Cave: St. John’s Gate, London, UK, 1754; pp. 493 and 549.
- Rossiter, P.B.; James, A.D. An epidemiological Model of Rinderpest. II. Simulations of the Behaviour of Rinderpest Virus in Popula-Tions. Trop. Anim. Health Prod. 1989, 21, 69–84. [Google Scholar] [CrossRef]
- Mariner, J.C.; House, J.A.; Mebus, C.A.; Sollod, A.E.; Chibeu, D.; Jones, B.A.; Roeder, P.L.; Admassu, B.; Klooster, G.G.V. Rinderpest Eradication: Appropriate Technology and Social Innovations. Science 2012, 337, 1309–1312. [Google Scholar] [CrossRef] [PubMed]
- Butler, D. Rinderpest Research Restarts. Nat. Cell Biol. 2013, 499, 264–265. [Google Scholar] [CrossRef]
- Butler, D. Sequence and Destroy: The Quest to Eliminate the Last Stocks of Deadly Rinderpest Virus. Nat. Cell Biol. 2019, 572, 18. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, K.; Visser, D.; Evans, B.; Vallat, B. Identifying and Reducing Remaining Stocks of Rinderpest Virus. Emerg. Infect. Dis. 2015, 21, 2117–2121. [Google Scholar] [CrossRef]
- Fournié, G.; Jones, B.A.; Beauvais, W.; Lubroth, J.; Njeumi, F.; Cameron, A.; Pfeiffer, D.U. The Risk of Rinderpest Re-Introduction in Post-Eradication Era. Prev. Vet. Med. 2014, 113, 175–184. [Google Scholar] [CrossRef]
- Hamilton, K.; Baron, M.; Matsuo, K.; Visser, D. Rinderpest Eradication: Challenges for Remaining Disease Free and Implications for Future Eradication Efforts. Rev. Sci. Tech. 2017, 36, 579–588. [Google Scholar] [CrossRef]
- King, S.; Rajko-Nenow, P.; Ropiak, H.M.; Ribeca, P.; Batten, C.; Baron, M.D. Full Genome Sequencing of Archived Wild Type and Vaccine Rinderpest Virus Isolates Prior to Their Destruction. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- The Lancet Infectious Diseases. Rinderpest, Smallpox, and the Imperative of Destruction. Lancet Infect. Dis. 2019, 19, 789. [Google Scholar] [CrossRef]
- Kurosawa, S.; Kobune, F.; Okuyama, K.; Sugiura, A. Effects of Antipyretics in Rinderpest Virus Infection in Rabbits. J. Infect. Dis. 1987, 155, 991–997. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, M.; Bandyopadhyay, S.K.; Shiotani, M.; Fujita, K.; Nuntaprasert, A.; Miura, R.; Baron, M.D.; Barrett, T.; Kai, C. Rinderpest Virus H Protein: Role in Determining Host Range in Rabbits. J. Gen. Virol. 2002, 83, 1457–1463. [Google Scholar] [CrossRef]
- Rahman, M.; Shaila, M.S.; Gopinathan, K.P. Baculovirus Display of Fusion Protein of Peste des Petits Ruminants Virus and Hemagglutination Protein of Rinderpest Virus and Immunogenicity of the Displayed Proteins in Mouse Model. Virology 2003, 317, 36–49. [Google Scholar] [CrossRef]
- Galbraith, S.E.; McQuaid, S.; Hamill, L.; Püllen, L.; Barrett, T.; Cosby, S.L. Rinderpest and Peste des Petits Ruminants Viruses Exhibit Neurovirulence in Mice. J. Neurovirol. 2002, 8, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, A.; Renukaradhya, G.J.; Rajasekhar, M.; Sita, G.L.; Shaila, M.S. Systemic and Oral Immunogenicity of Hemagglu-Tinin Protein of Rinderpest Virus Expressed by Transgenic Peanut Plants in a Mouse Model. Virology 2004, 323, 284–291. [Google Scholar] [CrossRef] [PubMed][Green Version]
- James, A.D.; Rossiter, P.B. An Epidemiological Model of Rinderpest. I. Description of the Model. Trop. Anim. Health Prod. 1989, 21, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Manore, C.; McMahon, B.; Fair, J.; Hyman, J.M.; Brown, M.; LaBute, M. Disease Properties, Geography, and Mitigation Strategies in a Simulation Spread of Rinderpest across the United States. Vet. Res. 2011, 42, 55. [Google Scholar] [CrossRef]
- Ortiz-Pelaez, A.; Pfeiffer, D.U.; Tempia, S.; Otieno, F.T.; Aden, H.H.; Costagli, R. Risk Mapping of Rinderpest Sero-Prevalence in Central and Southern Somalia Based on Spatial and Network Risk Factors. BMC Vet. Res. 2010, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Tillé, A.; Lefèvre, C.; Pastoret, P.-P.; Thiry, E. A Mathematical Model of Rinderpest Infection in Cattle Populations. Epidemiol. Infect. 1991, 107, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Mariner, J.; McDermott, J.; Heesterbeek, J.; Catley, A.; Roeder, P. A Model of Lineage-1 and Lineage-2 Rinderpest Virus Transmission in Pastoral Areas of East Africa. Prev. Vet. Med. 2005, 69, 245–263. [Google Scholar] [CrossRef][Green Version]
- Mourant, J.R.; Fenimore, P.W.; Manore, C.A.; McMahon, B.H. Decision Support for Mitigation of Livestock Disease: Rinderpest as a Case Study. Front. Vet. Sci. 2018, 5, 182. [Google Scholar] [CrossRef]
- Holzer, B.; Hodgson, S.; Logan, N.; Willett, B.; Baron, M.D. Protection of Cattle against Rinderpest by Vaccination with Wild-Type but Not Attenuated Strains of Peste des Petits Ruminants Virus. J. Virol. 2016, 90, 5152–5162. [Google Scholar] [CrossRef] [PubMed]
- De Swart, R.L.; Duprex, W.P.; Osterhaus, A. Rinderpest Eradication: Lessons for Measles Eradication? Curr. Opin. Virol. 2012, 2, 330–334. [Google Scholar] [CrossRef]
- Thomson, G.R.; Penrith, M.L. Eradication of Transboundary Animal Diseases: Can the Rinderpest Success Story be Repeated? Transbound. Emerg. Dis. 2017, 64, 459–475. [Google Scholar] [CrossRef][Green Version]
- Mukhopadhyay, A.K.; Taylor, W.P.; Roeder, P.L. Rinderpest: A Case Study of Animal Health Emergency Management. Rev. Sci. Tech. 1999, 18, 164–178. [Google Scholar] [CrossRef]
- Rao, T.V.; Bandyopadhyay, S.K. A Comprehensive Review of Goat Pox and Sheep Pox and Their Diagnosis. Anim. Health Res. Rev. 2000, 1, 127–136. [Google Scholar] [CrossRef]
- Ben Chehida, F.; Ayari-Fakhfakh, E.; Caufour, P.; Amdouni, J.; Nasr, J.; Messaoudi, L.; Ammar, H.H.; Sghaier, S.; Bernard, C.; Ghram, A.; et al. Sheep Pox in Tunisia: Current Status and Perspectives. Transbound. Emerg. Dis. 2018, 65, 50–63. [Google Scholar] [CrossRef]
- European Food Safety. A Scientific Opinion on Sheep and Goat Pox. EFSA J. 2014, 12, 1–122. [Google Scholar]
- Bhanuprakash, V.; Venkatesan, G.; Balamurugan, V.; Hosamani, M.; Yogisharadhya, R.; Chauhan, R.S.; Pande, A.; Mondal, B.; Singh, R.K. Pox Outbreaks in Sheep and Goats at Makhdoom (Uttar Pradesh), India: Evidence of Sheeppox Virus Infection in Goats. Transbound. Emerg. Dis. 2010, 57, 375–382. [Google Scholar] [CrossRef]
- Yune, N.; Abdela, N. Epidemiology and Economic Importance of Sheep and Goat Pox: A Review on Past and Current Aspects. J. Vet. Sci. Technol. 2017, 8, 1–5. [Google Scholar] [CrossRef]
- Mirzaie, K.; Barani, S.M.; Bokaie, S. A Review of Sheep Pox and Goat Pox: Perspective of Their Control and Eradication in Iran. J. Adv. Vet. Anim. Res. 2015, 2, 373–381. [Google Scholar] [CrossRef]
- Malesios, C.; Kostoulas, P.; Dadousis, K.; Demiris, N. An Early Warning Indicator for Monitoring Infectious Animal Diseases and Its Application in the Case of a Sheep Pox Epidemic. Stoch. Environ. Res. Risk Assess. 2017, 31, 329–337. [Google Scholar] [CrossRef]
- Aregahagn, S.; Tadesse, B.; Tegegne, B.; Worku, Y.; Mohammed, S. Spatiotemporal Distributions of Sheep and Goat Pox Disease Outbreaks in the Period. 2013–2019 in Eastern Amhara Region, Ethiopia. Vet. Med. Int. 2021, 2021, 6629510. [Google Scholar] [CrossRef] [PubMed]
- Babiuk, S.; Bowden, T.R.; Boyle, D.B.; Wallace, D.B.; Kitching, R.P. Capripoxviruses: An Emerging Worldwide Threat to Sheep, Goats and Cattle. Transbound. Emerg. Dis. 2008, 55, 263–272. [Google Scholar] [CrossRef]
- Gitao, C.G.; Mbindyo, C.; Omani, R.; Chemweno, V. Review of Sheep Pox Disease in Sheep. J. Vet. Med. Res. 2017, 4, 1–5. [Google Scholar]
- Balinsky, C.A.; Delhon, G.; Smoliga, G.; Prarat, M.; French, R.A.; Geary, S.J.; Rock, D.L.; Rodriguez, L.L. Rapid Preclinical Detection of Sheeppox Virus by a Real-Time PCR Assay. J. Clin. Microbiol. 2008, 46, 438–442. [Google Scholar] [CrossRef]
- Bhanuprakash, V.; Indrani, B.K.; Hosamani, M.; Singh, R.K. The Current Status of Sheep Pox Disease. Comp. Immunol. Microbiol. Infect. Dis. 2006, 29, 27–60. [Google Scholar]
- Bowden, T.R.; Coupar, B.E.; Babiuk, S.L.; White, J.R.; Boyd, V.; Duch, C.J.; Shiell, B.J.; Ueda, N.; Parkyn, G.R.; Copps, J.S.; et al. Detection of Antibodies Specific for Sheeppox and Goatpox Viruses Using Recombinant Capripoxvirus Antigens in an Indirect Enzyme-Linked Immunosorbent Assay. J. Virol. Methods 2009, 161, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Haegeman, A.; Zro, K.; Sammin, D.; Vandenbussche, F.; Ennaji, M.M.; de Clercq, K. Investigation of a Possible Link Between Vaccination and the 2010 Sheep Pox Epizootic in Morocco. Transbound. Emerg. Dis. 2016, 63, e278–e287. [Google Scholar] [CrossRef] [PubMed]
- Hosamani, M.; Mondal, B.; Tembhurne, P.A.; Bandyopadhyay, S.K.; Singh, R.K.; Rasool, T.J. Differentiation of Sheep Pox and Goat Poxviruses by Sequence Analysis and PCR-RFLP of P32 Gene. Virus Genes 2004, 29, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Pestova, Y.; Byadovskaya, O.; Kononov, A.; Sprygin, A. A Real Time High-Resolution Melting PCR Assay for Detection and Differentiation among Sheep Pox Virus, Goat Pox Virus, Field and Vaccine Strains of Lumpy Skin Disease Virus. Mol. Cell. Probes 2018, 41, 57–60. [Google Scholar] [CrossRef]
- Sumana, K.; Revanaiah, Y.; Shivachandra, S.B.; Mothay, D.; Apsana, R.; Saminathan, M.; Basavaraj, S.; Reddy, G.B.M. Molecular Phylogeny of Capripoxviruses Based on Major Immunodominant Protein (P32) Reveals Circulation of Host Specific Sheeppox and Goatpox Viruses in Small Ruminants of India. Infect. Genet. Evol. 2020, 85, 104472. [Google Scholar] [CrossRef]
- Tian, H.; Chen, Y.; Wu, J.; Shang, Y.; Liu, X. Serodiagnosis of Sheeppox and Goatpox Using an Indirect ELISA Based on Synthetic Peptide Targeting for the Major Antigen P32. Virol. J. 2010, 7, 245. [Google Scholar] [CrossRef]
- Kardjadj, M. Prevalence, Distribution, and Risk Factor for Sheep Pox and Goat Pox (SPGP) in Algeria. Trop. Anim. Health Prod. 2017, 49, 649–652. [Google Scholar] [CrossRef]
- OIE. Sheep Pox and Goat Pox. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; OIE: Paris, France, 2019; pp. 1513–1524. [Google Scholar]
- Boumart, Z.; Daouam, S.; Belkourati, I.; Rafi, L.; Tuppurainen, E.; Tadlaoui, K.O.; el Harrak, M. Comparative Innocuity and Efficacy of Live and Inactivated Sheeppox Vaccines. BMC Vet. Res. 2016, 12, 133. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.; Lamien, C.E.; Diallo, A. Capripox (Lumpy Skin Disease, Sheep Pox, and Goat Pox). In Veterinary Vaccines; Metwally, S., El Idrissi, A., Viljoen, G., Eds.; John Wiley & Sons, Ltd.: London, UK, 2021. [Google Scholar]
- Kali, K.; Kardjadj, M.; Touaghit, N.; Yahiaoui, F.; Ben-Mahdi, M.H. Understanding the Epidemiology of Sheep-Pox Outbreaks among Vaccinated Algerian Sheep and Post Vaccination Evaluation of the Antibodies Kinetics of the Commercially Used Vaccine. Comp. Immunol. Microbiol. Infect. Dis. 2019, 65, 128–131. [Google Scholar] [CrossRef]
- Boshra, H.; Truong, T.; Nfon, C.; Gerdts, V.; Tikoo, S.; Babiuk, L.A.; Kara, P.; Mather, A.; Wallace, D.; Babiuk, S. Capripoxvirus-Vectored Vaccines against Livestock Diseases in Africa. Antiviral Res. 2013, 98, 217–227. [Google Scholar] [CrossRef]
- Fakri, F.; Ghzal, F.; Daouam, S.; Elarkam, A.; Douieb, L.; Zouheir, Y.; Tadlaoui, K.O.; Fassi-Fihri, O. Development and Field Application of a New Combined Vaccine against Peste des Petits Ruminants and Sheep Pox. Trials Vaccinol. 2015, 4, 33–37. [Google Scholar] [CrossRef]
- Didarkhah, M.; Vatandoost, M.; Dirandeh, E.; Davachi, N.D. Characterization and Pattern of Culling in Goats. Arch. Razi. Inst. 2019, 74, 441–446. [Google Scholar] [PubMed]
- Garner, M.G.; Sawarkar, S.D.; Brett, E.K.; Edwards, J.R.; Kulkarni, V.B.; Boyle, D.B.; Singh, S.N. The Extent and Impact of Sheep Pox and Goat Pox in the State of Maharashtra, India. Trop. Anim. Health Prod. 2000, 32, 205–223. [Google Scholar] [CrossRef] [PubMed]
- Limon, G.; Gamawa, A.A.; Ahmed, A.I.; Lyons, N.A.; Beard, P.M. Epidemiological Characteristics and Economic Impact of Lumpy Skin Disease, Sheeppox and Goatpox Among Subsistence Farmers in Northeast Nigeria. Front. Vet. Sci. 2020, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.H.; Lila, M.A.M.; Rahaman, N.Y.A.; Lai, H.L.T.; Nguyen, L.T.; Do, K.V.; Noordin, M.M. Epidemiology and Clinico-Pathological Characteristics of Current Goat Pox Outbreak in North Vietnam. BMC Vet. Res. 2020, 16, 128. [Google Scholar] [CrossRef]
- Chamchod, F. Modeling the Spread of Capripoxvirus Among Livestock and Optimal Vaccination Strategies. J. Theor. Biol. 2018, 437, 179–186. [Google Scholar] [CrossRef]
- Singh, B.; Prasad, S. Modelling of Economic Losses due to Some Important Diseases in Goats in India. Agric. Econ. Res. Rev. 2008, 21, 297–302. [Google Scholar]
- Zhuravlyova, V.A.; Lunitsin, A.V.; Kneize, A.V.; Guzalova, A.G.; Balyshev, V.M. Epizootic Situation and Modeling of Potential Nosoareals of Peste des Petits Ruminants, Sheep and Goat Pox and Rift Valley Fever Up to 2030. Agric. Biol. 2020, 55, 343–354. [Google Scholar] [CrossRef]
- Chervyakova, O.V.; Zaitsev, V.L.; Iskakov, B.K.; Tailakova, E.T.; Strochkov, V.M.; Sultankulova, K.T.; Sandybayev, N.T.; Stanbekova, G.E.; Beisenov, D.K.; Abduraimov, Y.O.; et al. Recombinant Sheep Pox Virus Proteins Elicit Neutralizing Antibodies. Viruses 2016, 8, 159. [Google Scholar] [CrossRef]
- Hamdi, J.; Bamouh, Z.; Jazouli, M.; Boumart, Z.; Tadlaoui, K.O.; Fihri, O.F.; el Harrak, M. Experimental Evaluation of the Cross-Protection between Sheeppox and Bovine Lumpy Skin Vaccines. Sci. Rep. 2020, 10, 8888. [Google Scholar] [CrossRef]
- Kitching, R.P. Passive Protection of Sheep against Capripoxvirus. Res. Vet. Sci. 1986, 41, 247–250. [Google Scholar] [CrossRef]
- Kitching, R.P.; Taylor, W.P. Clinical and Antigenic Relationship between Isolates of Sheep and Goat Pox Viruses. Trop. Anim. Health Prod. 1985, 17, 64–74. [Google Scholar] [CrossRef]
- Zhugunissov, K.; Bulatov, Y.; Orynbayev, M.; Kutumbetov, L.; Abduraimov, Y.; Shayakhmetov, Y.; Taranov, D.; Amanova, Z.; Mambetaliyev, M.; Absatova, Z.; et al. Goatpox Virus (G20-LKV) Vaccine Strain Elicits a Protective Response in Cattle against Lumpy Skin Disease at Challenge with Lumpy Skin Disease Virulent Field Strain in a Comparative Study. Vet. Microbiol. 2020, 245, 108695. [Google Scholar] [CrossRef]
- Plowright, W.; MacLeod, W.G.; Ferris, R.D. The Pathogenesis of Sheep Pox in the Skin of Sheep. J. Comp. Pathol. 2012, 146, 97–105. [Google Scholar] [CrossRef]
- Wolff, J.; el Rahman, S.A.; King, J.; El-Beskawy, M.; Pohlmann, A.; Beer, M.; Hoffmann, B. Establishment of a Challenge Model for Sheeppox Virus Infection. Microorganisms 2020, 8, 2001. [Google Scholar] [CrossRef]
- Bowden, T.R.; Babiuk, S.L.; Parkyn, G.R.; Copps, J.S.; Boyle, D.B. Capripoxvirus Tissue Tropism and Shedding: A Quantitative Study in Experimentally Infected Sheep and Goats. Virology 2008, 371, 380–393. [Google Scholar] [CrossRef]
- Brocchi, E.; Zhang, G.; Knowles, N.J.; Wilsden, G.; McCauley, J.W.; Marquardt, O.; Ohlinger, V.F.; de Simone, F. Molecular Epidemiology of Recent Outbreaks of Swine Vesicular Disease: Two Genetically and Antigenically Distinct Variants in Europe, 1987–1994. Epidemiol. Infect. 1997, 118, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Kitching, R.P. Swine Vesicular Disease: An Overview. Vet. J. 2000, 160, 192–201. [Google Scholar] [PubMed]
- Tamba, M.; Plasmati, F.; Brocchi, E.; Ruocco, L. Eradication of Swine Vesicular Disease in Italy. Viruses 2020, 12, 1269. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel Animal Health and Welfare (AHAW) Scientific Opinion on Swine Vesicular Disease and Vesicular Stomatitis. EFSA J. 2012, 10, 2631. [CrossRef]
- Terpstra, C. Vesicular Swine Disease in The Netherlands. Tijdschr Diergeneeskd 1992, 117, 623–626. [Google Scholar] [PubMed]
- Dekker, A.; Moonen, P.; de Boer-Luijtze, E.A.; Terpstra, C. Pathogenesis of Swine Vesicular Disease after Exposure of Pigs to an Infected Environment. Vet. Microbiol. 1995, 45, 243–250. [Google Scholar] [CrossRef]
- Dibaba, A.B. The Risk of Introduction of Swine Vesicular Disease Virus into Kenya via Natural Sausage Casings Imported from Italy. Prev. Vet. Med. 2019, 169, 104703. [Google Scholar] [CrossRef]
- Yang, M.; Gagliardi, K.; McIntyre, L.; Xu, W.; Goolia, M.; Ambagala, T.; Brocchi, E.; Grazioli, S.; Hooper–McGrevy, K.; Nfon, C.; et al. Development and Evaluation of Swine Vesicular Disease Isotype-Specific Antibody Elisas Based on Recombinant Virus-Like Particles. Transbound. Emerg. Dis. 2020, 67, 406–416. [Google Scholar] [CrossRef]
- Brocchi, E.; Berlinzani, A.; Gamba, D.; de Simone, F. Development of Two Novel Monoclonal Antibody-Based Elisas for the Detection of Antibodies and the Identification of Swine Isotypes against Swine Vesicular Disease Virus. J. Virol. Methods 1995, 52, 155–167. [Google Scholar] [CrossRef]
- Escribano-Romero, E.; Jimenez-Clavero, M.A.; Gomes, P.; García-Ranea, J.A.; Ley, V. Heparan Sulphate Mediates Swine Vesicular Disease Virus Attachment to the Host Cell. 2004, 85, 653–663. J. Gen. Virol. 2004, 85, 653–663. [Google Scholar] [CrossRef][Green Version]
- Fry, E.E.; Knowles, N.J.; Newman, J.W.I.; Wilsden, G.; Rao, Z.; King, A.M.Q.; Stuart, D.I. Crystal Structure of Swine Vesicular Disease Virus and Implications for Host Adaptation. J. Virol. 2003, 77, 5475. [Google Scholar] [CrossRef][Green Version]
- Martín-Acebes, M.A.; González-Magaldi, M.; Rosas, M.F.; Borrego, B.; Brocchi, E.; Armas-Portela, R.; Sobrino, F. Subcellular Distribution of Swine Vesicular Disease Virus Proteins and Alterations Induced in Infected Cells: A Comparative Study with Foot-and-Mouth Disease Virus and Vesicular Stomatitis Virus. Virology 2008, 374, 432–443. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Burrows, R.; Greig, A.; Goodridge, D. Swine Vesicular Disease. Res. Vet. Sci. 1973, 15, 141–144. [Google Scholar] [CrossRef]
- Chu, R.M.; Moore, D.M.; Conroy, J.D. Experimental Swine Vesicular Disease, Pathology and Immunofluorescence Studies. Can. J. Comp. Med. 1979, 43, 29–38. [Google Scholar]
- Lin, F.; Mackay, D.K.J.; Knowles, N.J. The Persistence of Swine Vesicular Disease Virus Infection in Pigs. Epidemiol. Infect. 1998, 121, 459–472. [Google Scholar] [CrossRef]
- Mann, J.A.; Hutchings, G.H. Swine Vesicular Disease: Pathways of Infection. J. Hyg. 1980, 84, 355–363. [Google Scholar] [CrossRef]
- Mowat, G.N.; Prince, M.J.; Spier, R.E.; Staple, R.F. Preliminary Studies on the Development of a Swine Vesicular Disease Vaccine. Arch. Gesamte Virusforsch 1974, 44, 350–360. [Google Scholar] [CrossRef]
- Letchworth, G.J.; Rodriguez, L.L.; del Cbarrera, J. Vesicular Stomatitis. Vet. J. 1999, 157, 239–260. [Google Scholar] [CrossRef]
- Whelan, S.P.J. Vesicular Stomatitis Virus. In Encyclopedia of Virology, 3rd ed.; Mahy, B.W.J., Van Regenmortel, M.H.V., Eds.; Academic Press: Oxford, UK, 2008; pp. 291–299. [Google Scholar]
- Vanleeuwen, J.A.; Rodriguez, L.L.; Waltner-Toews, D. Cow, Farm, and Ecologic Risk Factors of Clinical Vesicular Stomatitis on Costa Rican Dairy Farms. Am. J. Trop. Med. Hyg. 1995, 53, 342–350. [Google Scholar] [CrossRef]
- Webb, P.A.; Monath, T.P.; Reif, J.S.; Smith, G.C.; Kemp, G.E.; Lazuick, J.S.; Walton, T.E. Epizootic Vesicular Stomatitis in Colorado, 1982: Epidemiologic Studies along the Northern Colorado Front Range. Am. J. Trop. Med. Hyg. 1987, 36, 183–188. [Google Scholar] [CrossRef]
- Webb, P.A.; Holbrook, F.R. Vesicular Stomatitis. In The Arboviruses: Epidemiology and Ecology; Monath, T.P., Ed.; CRC Press: Boca Raton, FL, USA, 1989. [Google Scholar]
- Fultz, P.N.; Holland, J.J. Differing Responses of Hamsters to Infection by Vesicular Stomatitis Virus Indiana and New Jersey Serotypes. Virus Res. 1985, 3, 129–140. [Google Scholar] [CrossRef]
- Gresser, I.; Tovey, M.G.; Bourali-Maury, C. Efficacy of Exogenous Interferon Treatment Initiated after Onset of Multiplication of Vesicular Stomatitis Virus in the Brains of Mice. J. Gen. Virol. 1975, 27, 395–398. [Google Scholar] [CrossRef] [PubMed]
- Huneycutt, B.S.; Plakhov, I.V.; Shusterman, Z.; Bartido, S.M.; Huang, A.; Reiss, C.S.; Aoki, C. Distribution of Vesicular Stomatitis Virus Proteins in the Brains Of BALB/C Mice Following Intranasal Inoculation: An Immunohistochemical Analysis. Brain Res. 1994, 635, 81–95. [Google Scholar] [CrossRef]
- Leder, R.R.; Maas, J.; Lane, V.M.; Evermann, J.F. Epidemiologic Investigation of Vesicular Stomatitis in a Dairy and Its Economic Impact. Bov. Pract. 1983, 1983, 45–49. [Google Scholar]
- Clarke, D.K.; Hendry, R.M.; Singh, V.; Rose, J.K.; Seligman, S.J.; Klug, B.; Kochhar, S.; Mac, L.M.; Carbery, B.; Chen, R.T. Live Virus Vaccines Based on a Vesicular Stomatitis Virus (VSV) Backbone: Standardized Template with Key Considerations for a Risk/Benefit Assessment. Vaccine 2016, 34, 6597–6609. [Google Scholar] [CrossRef] [PubMed]
- Campbell, Z.; Coleman, P.; Guest, A.; Kushwaha, P.; Ramuthivheli, T.; Osebe, T.; Perry, B.; Salt, J. Prioritizing Smallholder Animal Health Needs in East. Africa, West. Africa, and South. Asia Using Three Approaches: Literature Review, Expert Workshops, and Practitioner Surveys. Prev. Vet. Med. 2021, 189, 105279. [Google Scholar] [CrossRef] [PubMed]
- Hopker, A.; Pandey, N.; Saikia, D.; Goswami, J.; Hopker, S.; Saikia, R.; Sargison, N. Spread and Impact of Goat Pox (“Sagolay Bohonta”) in a Village Smallholder Community around Kaziranga National Park, Assam, India. Trop. Anim. Health Prod. 2019, 51, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Holzmann, H.; Hengel, H.; Tenbusch, M.; Doerr, H.W. Eradication of Measles: Remaining Challenges. Med. Microbiol. Immunol. 2016, 205, 201–208. [Google Scholar] [CrossRef]
- Food and Drug Administration (FDA) New Drug and Biological Drug Products; Evidence Needed to Demonstrate Effectiveness of New Drugs when Human Efficacy Studies Are Not Ethical or Feasible. Final Rule Fed. Regist. 2002, 67, 37988–37998.
- Gronvall, G.K.; Trent, D.; Borio, L.; Brey, R.; Nagao, L. The FDA Animal Efficacy Rule and Biodefense. Nat. Biotechnol. 2007, 25, 1084–1087. [Google Scholar]
- Domingo, E.; Perales, C. Viral Quasispecies. PLoS Genet. 2019, 15, e1008271. [Google Scholar] [CrossRef]


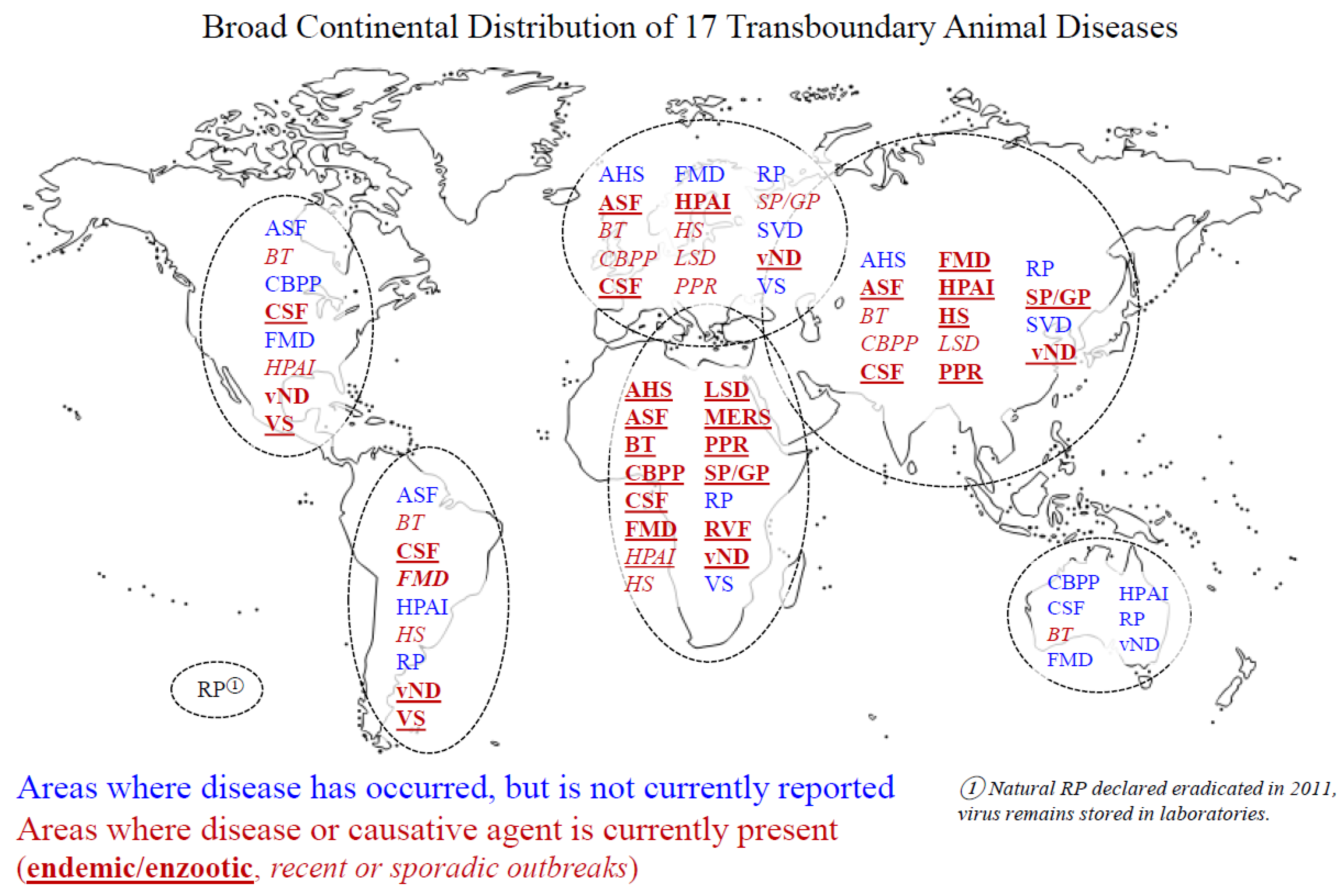{kind=link}
| Disease | Causative Agent | Species Affected | Symptoms | Other |
|---|---|---|---|---|
| African horse sickness | African horse sickness virus (Orbivirus) | Equids; primarily horses | Horses—acute (pulmonary) and chronic (cardiac) with high morbidity and mortality, mules and donkeys—mild disease, zebras—usually asymptomatic | |
| African swine fever | African swine fever virus (Asfivirus) | Domestic and wild suids | Sudden death, shock, hemorrhagic fever, pulmonary edema, depression, anorexia, thrombocytopenia, lymphopenia | |
| Avian influenza | Avian influenza virus (Influenza A) | Domestic poultry; birds and mammals | Highly pathogenic avian influenza (H5 and H7) causes high rates of mortality, respiratory symptoms, sinus or head swelling, depression, anorexia, cyanosis, incoordination, neurologic symptoms, diarrhea | Zoonotic |
| Bluetongue | Bluetongue virus (Orbivirus) | Domestic and wild ruminants; primarily sheep | Fever, swelling, vascular injury and hemorrhage, ulceration, pulmonary edema, muscle necrosis; or asymptomatic | |
| Classical swine fever | Classical swine fever virus (Pestivirus) | Domestic and wild suids | Acute, chronic, and prenatal presentations; sudden death, hemorrhagic fever, stillbirth, abortion, mummification, malformations, persistent infection, congenital tremor | |
| Contagious bovine pleuropneumonia | Mycoplasma mycoides subsp. Mycoides | Domestic and wild large ruminants; primarily cattle | Fever, inappetence, depression, labored breathing, coughing, nasal discharge, salivation (may vary from sudden death to chronic subclinical carrier) | |
| Foot and mouth disease | Foot and mouth disease virus (Aphthovirus) | Cloven-hooved animals | Fever, vesicles on the feet, tongue, snout, muzzle, mammary glands, genital mucosa, or other mucosal sites, inappetence, lameness | |
| Hemorrhagic septicemia | Pasteurella multocida | Cattle and buffalo | Fever, submandibular and brisket edema, respiratory distress and mucopurulent or bloody nasal discharge (may vary from sudden death to chronic subclinical carrier) | |
| Lumpy skin disease | Lumpy skin disease virus (Capripoxvirus) | Cattle and buffalo | Lacrimation, nasal discharge, inappetence, lymphadenopathy, fever, drop in milk production, lameness, nodular skin lesions, sometimes death | |
| Middle East respiratory syndrome | Middle East respiratory syndrome coronavirus (Betacoronavirus) | Camels | Mild upper respiratory disease | Zoonotic |
| Newcastle disease | Newcastle disease virus (Avulavirus) | Primarily domestic chickens; reptiles, birds, and mammals possible | Five pathotypes (viscerotropic velogenic, neurotropic velogenic, mesogenic, lentogenic, and asymptomatic enteric); neurologic and respiratory disease; asymptomatic | Zoonotic |
| Peste des petits ruminants | Peste des petits ruminants virus (Morbillivirus) | Domestic and wild ruminants; primarily goats and sheep | Inappetence, emaciation, depression, fever, diarrhea, nasal and ocular discharge, pneumonia and erosive and necrotic stomatitis, death | Targeted for eradication by 2030 |
| Rift Valley fever | Rift Valley fever virus (Phlebovirus) | Ruminants | Abortion, fever, lymphadenopathy and inappetence; weakness, nasal discharge, and bloody diarrhea (sheep); hypersalivation, diarrhea, and decreased milk production (cattle) | Zoonotic |
| Rinderpest | Rinderpest virus (Morbillivirus) | Domestic and wild ungulates | Fever, erosive mouth lesions, nasal and ocular discharge, bloody diarrhea, dehydration, weakness, death | Eradicated, 2011 |
| Sheeppox and goatpox | Sheeppox virus and Goatpox virus (Capripoxvirus) | Sheep and goats | Fever, labored breathing, depression, inappetence and lymphadenopathy, progressive skin lesions: macular, popular, vesicular, pustular and scabbing; sometimes death | |
| Swine vesicular disease | Swine vesicular disease virus (Enterovirus) | Suids | Infrequent and mild; influenza-like | |
| Vesicular stomatitis | Vesicular stomatitis virus (Vesiculovirus) | Horses, cattle and suids; rarely sheep and goats | Vesicular lesions on feet, snout, lips, and tongue |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clemmons, E.A.; Alfson, K.J.; Dutton, J.W., III. Transboundary Animal Diseases, an Overview of 17 Diseases with Potential for Global Spread and Serious Consequences. Animals 2021, 11, 2039. https://doi.org/10.3390/ani11072039
Clemmons EA, Alfson KJ, Dutton JW III. Transboundary Animal Diseases, an Overview of 17 Diseases with Potential for Global Spread and Serious Consequences. Animals. 2021; 11(7):2039. https://doi.org/10.3390/ani11072039
Chicago/Turabian StyleClemmons, Elizabeth A., Kendra J. Alfson, and John W. Dutton, III. 2021. "Transboundary Animal Diseases, an Overview of 17 Diseases with Potential for Global Spread and Serious Consequences" Animals 11, no. 7: 2039. https://doi.org/10.3390/ani11072039
APA StyleClemmons, E. A., Alfson, K. J., & Dutton, J. W., III. (2021). Transboundary Animal Diseases, an Overview of 17 Diseases with Potential for Global Spread and Serious Consequences. Animals, 11(7), 2039. https://doi.org/10.3390/ani11072039