
Bayesian Analysis of the Effects of Olive Oil-Derived Antioxidants on Cryopreserved Buck Sperm Parameters

 ,
,  , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
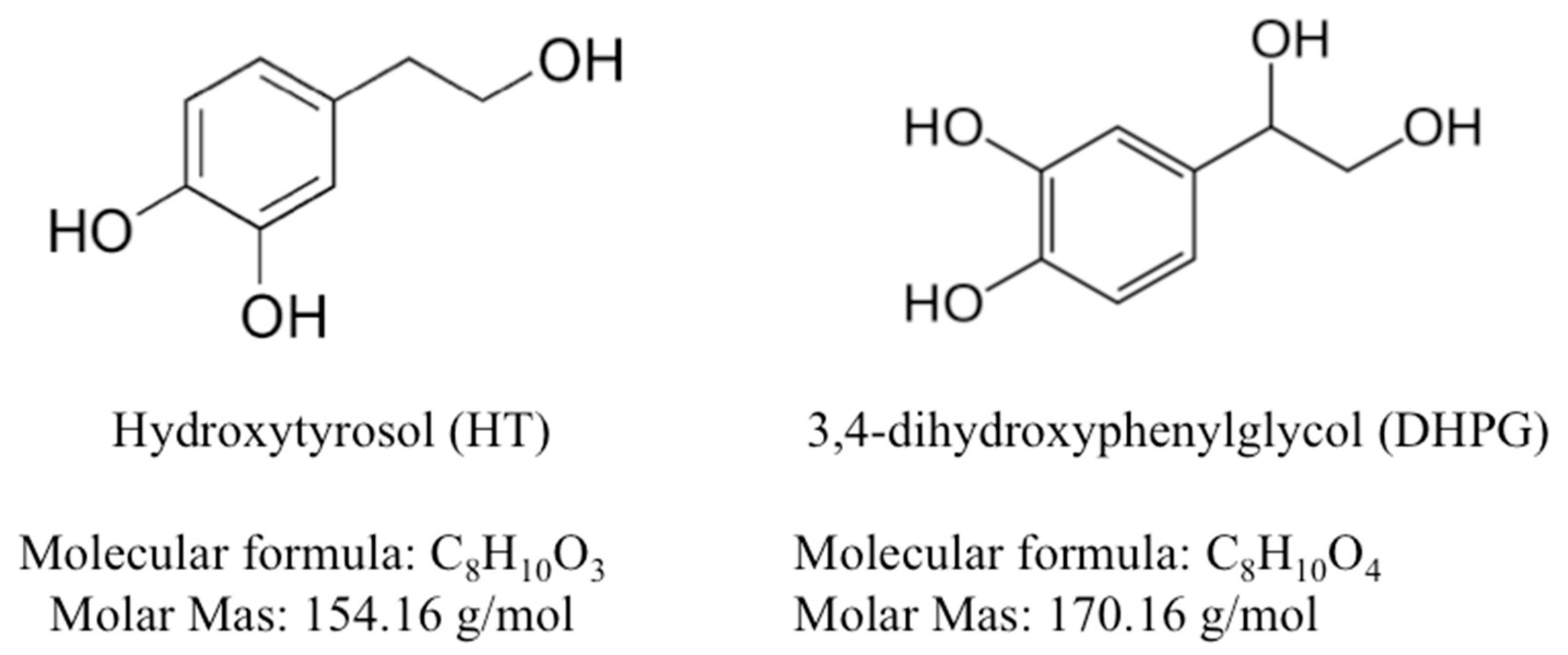
2.1. Chemicals
2.2. Animals and Semen Collection
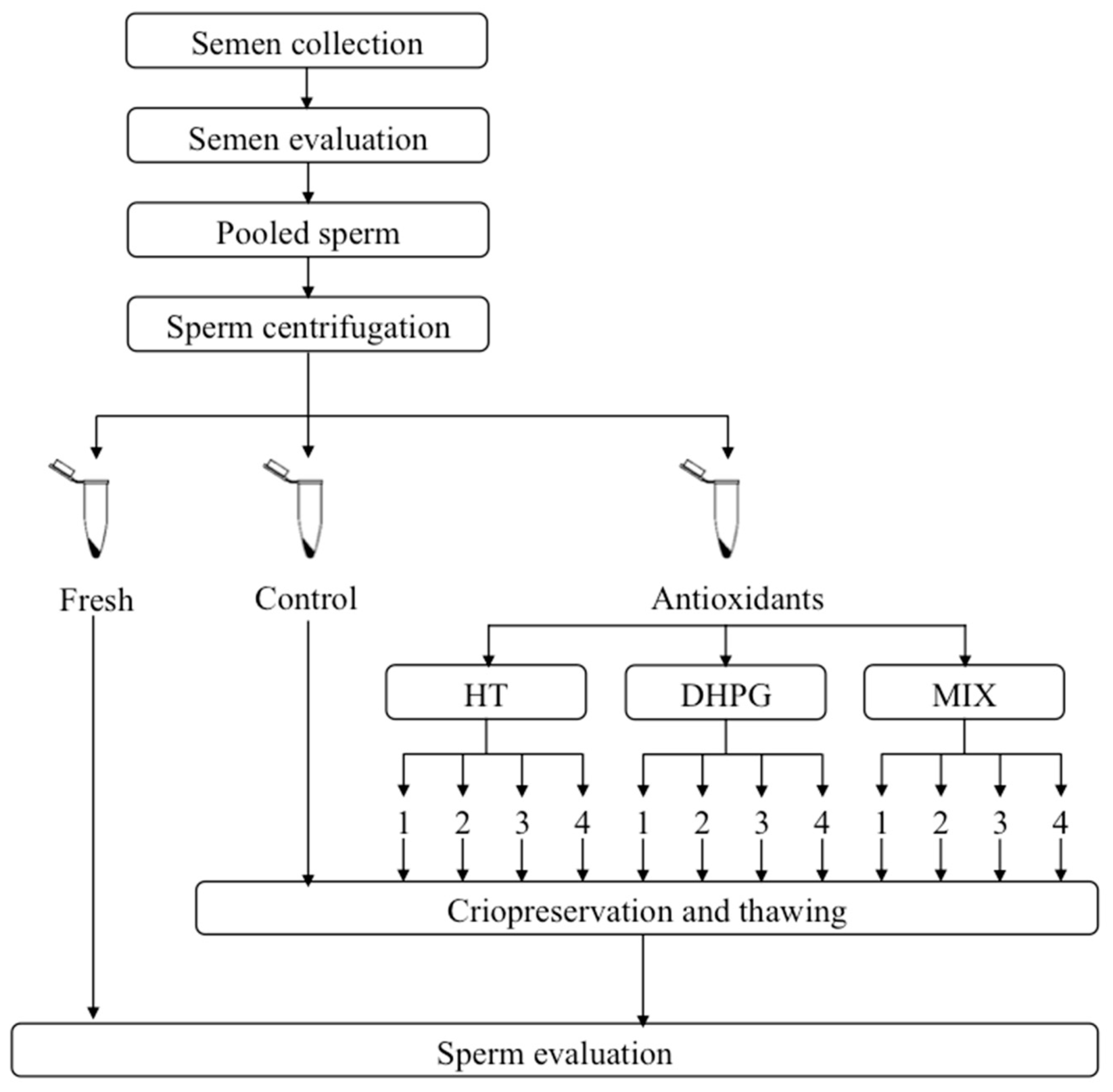
2.3. Experimental Design
2.4. Semen Dilution and Freezing
2.5. Sperm Quality Assessment
2.5.1. Motility
2.5.2. Flow Cytometer
2.5.3. Viability
2.5.4. Acrosome Integrity
2.5.5. Mitochondrial Membrane Potential
2.5.6. Lipid Peroxidation
2.6. Data Analysis
3. Results
3.1. Prior Descriptive Statistics
3.2. Bayesian Inference of Olive Oil Derived Antioxidant Effect
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yoon, S.J.; Rahman, M.S.; Kwon, W.S.; Park, Y.J.; Pang, M.G. Addition of Cryoprotectant Significantly Alters the Epididymal Sperm Proteome. PLoS ONE 2016, 11, e0152690. [Google Scholar]
- Kargar, R.; Forouzanfar, M.; Ghalamkari, G.; Nasr Esfahani, M.H. Dietary flax seed oil and/or Vitamin E improve sperm parameters of cloned goats following freezing-thawing. Cryobiology 2017, 74, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Bucak, M.N.; Sarıözkan, S.; Tuncer, P.B.; Ulutaş, P.A.; Akçadağ, H.İ. Effect of antioxidants on microscopic semen parameters, lipid peroxidation and antioxidant activities in Angora goat semen following cryopreservation. Small Rumin. Res. 2009, 81, 90–95. [Google Scholar] [CrossRef]
- Mata-Campuzano, M.; Álvarez-Rodríguez, M.; Álvarez, M.; Tamayo-Canul, J.; Anel, L.; de Paz, P.; Martínez-Pastor, F. Post-thawing quality and incubation resilience of cryopreserved ram spermatozoa are affected by antioxidant supplementation and choice of extender. Theriogenology 2015, 83, 520–528. [Google Scholar] [CrossRef] [PubMed]
- De Lamirande, E.; Gagnon, C. Impact of reactive oxygen species on spermatozoa: A balancing act between beneficial and detrimental effects. Hum. Reprod. 1995, 10 (Suppl. 1), 15–21. [Google Scholar] [CrossRef]
- Aziz, N.; Saleh, R.A.; Sharma, R.K.; Lewis-Jones, I.; Esfandiari, N.; Thomas, A.J.; Agarwal, A. Novel association between sperm reactive oxygen species production, sperm morphological defects, and the sperm deformity index. Fertil. Steril. 2004, 81, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Saleh, R.A. Role of oxidants in male infertility: Rationale, significance, and treatment. Urol. Clin. N. Am. 2002, 29, 817–828. [Google Scholar] [CrossRef]
- Shafiei, M.; Forouzanfar, M.; Hosseini, S.M.; Nasr Esfahani, M.H. The effect of superoxide dismutase mimetic and catalase on the quality of postthawed goat semen. Theriogenology 2015, 83, 1321–1327. [Google Scholar] [CrossRef] [PubMed]
- Alcay, S.; Gokce, E.; Toker, M.B.; Onder, N.T.; Ustuner, B.; Uzabacı, E.; Gul, Z.; Cavus, S. Freeze-dried egg yolk based extenders containing various antioxidants improve post-thawing quality and incubation resilience of goat spermatozoa. Cryobiology 2016, 72, 269–273. [Google Scholar] [CrossRef]
- Yousefian, I.; Emamverdi, M.; Karamzadeh-Dehaghani, A.; Sabzian-Melei, R.; Zhandi, M.; Zare-Shahneh, A. Attenuation of cryopreservation-induced oxidative stress by antioxidant: Impact of Coenzyme Q10 on the quality of post-thawed buck spermatozoa. Cryobiology 2018, 81, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Ren, F.; Feng, T.; Dai, G.; Wang, Y.; Zhu, H.; Hu, J. Lycopene and alpha-lipoic acid improve semen antioxidant enzymes activity and cashmere goat sperm function after cryopreservation. Cryobiology 2018, 84, 27–32. [Google Scholar] [CrossRef]
- Haydari, M.; Maresca, V.; Rigano, D.; Taleei, A.; Shahnejat-Bushehri, A.A.; Hadian, J.; Sorbo, S.; Guida, M.; Manna, C.; Piscopo, M.; et al. Salicylic Acid and Melatonin Alleviate the Effects of Heat Stress on Essential Oil Composition and Antioxidant Enzyme Activity in Mentha × piperita and Mentha arvensis L. Antioxidants 2019, 8, 547. [Google Scholar] [CrossRef] [Green Version]
- Tortora, F.; Notariale, R.; Maresca, V.; Good, K.V. Phenol-Rich Feijoa sellowiana (Pineapple Guava) Extracts Protect Human Red Blood Cells from Mercury-Induced Cellular Toxicity. Antioxidants 2019, 8, 220. [Google Scholar] [CrossRef] [Green Version]
- Boo, Y.C. Human Skin Lightening Efficacy of Resveratrol and Its Analogs: From in Vitro Studies to Cosmetic Applications. Antioxidants 2019, 8, 332. [Google Scholar] [CrossRef] [Green Version]
- Arando, A.; Delgado, J.V.; Bermúdez-Oria, A.; León, J.M.; Fernández-Prior, Á.; Nogales, S.; Pérez-Marín, C.C. Effect of olive-derived antioxidants (3,4-dihydroxyphenylethanol and 3,4 dihydroxyphenylglycol) on sperm motility and fertility in liquid ram sperm stored at 15 °C or 5 °C. Reprod. Domest. Anim. 2020, 55, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Bolanos, J.G.; Lopez, O.; Fernandez-Bolanos, J.; Rodriguez-Gutierrez, G. Hydroxytyrosol and derivatives: Isolation, synthesis, and biological properties. Curr. Org. Chem. 2008, 12, 442–463. [Google Scholar] [CrossRef]
- Bermúdez-Oria, A.; Rodríguez-Gutiérrez, G.; Rubio-Senent, F.; Lama-Muñoz, A.; Fernández-Bolaños, J. Complexation of hydroxytyrosol and 3,4-dihydroxyphenylglycol with pectin and their potential use for colon targeting. Carbohydr. Polym. 2017, 163, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Choucair, F.; Saliba, E.; Jaoude, I.A.; Hazzouri, M. Antioxidants modulation of sperm genome and epigenome damage: Fact or fad? Converging evidence from animal and human studies. Middle East Fertil Soc. J. 2018, 23, 85–90. [Google Scholar] [CrossRef]
- Roche, M.; Dufour, C.; Loonis, M.; Reist, M.; Carrupt, P.-A.; Dangles, O. Olive phenols efficiently inhibit the oxidation of serum albumin-bound linoleic acid and butyrylcholine esterase. Biochim. Biophys. Acta Gen. Subj. 2009, 1790, 240–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, T.; Ibi, M.; Matsuno, K.; Nakashima, S.; Tanigawa, T.; Yoshikawa, T.; Yabe-Nishimura, C. An endogenous metabolite of dopamine, 3, 4-dihydroxyphenylethanol, acts as a unique cytoprotective agent against oxidative stress-induced injury. Free Radic. Biol. Med. 2004, 36, 555–564. [Google Scholar] [CrossRef]
- Rodríguez, G.; Rodríguez, R.; Fernández-Bolaños, J.; Guillén, R.; Jiménez, A. Antioxidant activity of effluents during the purification of hydroxytyrosol and 3, 4-dihydroxyphenyl glycol from olive oil waste. Eur. Food Res. Technol. 2007, 224, 733–741. [Google Scholar] [CrossRef]
- Hamden, K.; Allouche, N.; Damak, M.; Carreau, S.; Elfeki, A. Potential protective effect of hydroxytyrosol against glucose-induced toxicity in spermatozoa in vitro. Asia Life Sci. 2010, 19, 333–344. [Google Scholar]
- Kedechi, S.; Zribi, N.; Louati, N.; Menif, H.; Sellami, A.; Lassoued, S.; Ben Mansour, R.; Keskes, L.; Rebai, T.; Chakroun, N. Antioxidant effect of hydroxytyrosol on human sperm quality during in vitro incubation. Andrologia 2017, 49, e12595. [Google Scholar] [CrossRef] [PubMed]
- Krishnappa, B.; Srivastava, S.; Kumar, D.; Ghosh, S.; Kalyan, D.; Paul, R.; Bahire, S.; Naqvi, S. Effect of hydroxytyrosol on sperm post-thaw motion and velocity of cryopreserved ram semen. Indian J. Small Rumin. 2018, 24, 75–79. [Google Scholar] [CrossRef]
- Arando, A.; Delgado, J.; Fernández-Prior, A.; León, J.; Bermúdez-Oria, A.; Nogales, S.; Pérez-Marín, C. Effect of different olive oil-derived antioxidants (hydroxytyrosol and 3, 4-dihydroxyphenylglycol) on the quality of frozen-thawed ram sperm. Cryobiology 2019, 86, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Bolaños, J.; Rodríguez, G.; Rodríguez, R.; Heredia, A.; Guillén, R.; Jiménez, A. Production in large quantities of highly purified hydroxytyrosol from liquid− solid waste of two-phase olive oil processing or “Alperujo”. J. Agric. Food Chem. 2002, 50, 6804–6811. [Google Scholar] [CrossRef]
- Fernández-Bolaños Guzmán, J.; Rodríguez-Gutiérrez, G.; Lama Muñoz, A.; Fernández-Bolaños Guzmán, J.M.; Maya, I.; López López, Ó.; Marset Castro, A. Method for obtaining hydroxytyrosol extract, mixture of hydroxytyrosol and 3, 4-dihydroxyphenylglycol extract, and hydroxytyrosyl acetate extract, from by-products of the olive tree, and the purification thereof. International Patent Application No. PCT/ES2012/070491. U.S. Patent Application No. WO 2013/007850, 3 July, 2013. [Google Scholar]
- Evans, G.; Maxwell, W.C. Salamons’ Artificial Insemination of Sheep and Goats; Butterworths: Sydney, Australia, 1987. [Google Scholar]
- Lopes, S.A.F.; Rosa, H.J.D.; Chaveiro, A.; da Silva, F.M. Influence of Different Freezing Curves on the Acrosome Integrity of Male Goat Sperm Cells. Am. J. Anim. Vet. Sci. 2021, 16, 56–61. [Google Scholar] [CrossRef]
- Lee, J.A.; Spidlen, J.; Boyce, K.; Cai, J.; Crosbie, N.; Dalphin, M.; Furlong, J.; Gasparetto, M.; Goldberg, M.; Goralczyk, E.M.; et al. MIFlowCyt: The minimum information about a flow cytometry experiment. Cytom. Part A 2008, 73A, 926–930. [Google Scholar] [CrossRef] [Green Version]
- Arando, A.; Gonzalez, A.; Delgado, J.; Arrebola, F.; Perez-Marín, C. Storage temperature and sucrose concentrations affect ram sperm quality after vitrification. Anim. Reprod. Sci. 2017, 181, 175–185. [Google Scholar] [CrossRef]
- Petrunkina, A.; Waberski, D.; Bollwein, H.; Sieme, H. Identifying non-sperm particles during flow cytometric physiological assessment: A simple approach. Theriogenology 2010, 73, 995–1000. [Google Scholar] [CrossRef]
- Santiani, A.; Ugarelli, A.; Evangelista-Vargas, S. Characterization of functional variables in epididymal alpaca (Vicugna pacos) sperm using imaging flow cytometry. Anim. Reprod. Sci. 2016, 173, 49–55. [Google Scholar] [CrossRef]
- Hallap, T.; Nagy, S.; Jaakma, Ü.; Johannisson, A.; Rodriguez-Martinez, H. Mitochondrial activity of frozen-thawed spermatozoa assessed by MitoTracker Deep Red 633. Theriogenology 2005, 63, 2311–2322. [Google Scholar] [CrossRef]
- Thuwanut, P.; Axner, E.; Johanisson, A.; Chatdarong, K. Detection of Lipid Peroxidation Reaction in Frozen-Thawed Epididymal Cat Spermatozoa Using BODIPY581/591 C11. Reprod. Domest. Anim. 2009, 44, 373–376. [Google Scholar] [CrossRef]
- Cleophas, T.J.; Zwinderman, A.H. Bayesian Analysis of Variance (Anova). In Modern Bayesian Statistics in Clinical Research; Cleophas, T.J., Zwinderman, A.H., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 83–89. [Google Scholar]
- Martins-Bessa, A.; Quaresma, M.; Leiva, B.; Calado, A.; Navas González, F.J. Bayesian Linear Regression Modelling for Sperm Quality Parameters Using Age, Body Weight, Testicular Morphometry, and Combined Biometric Indices in Donkeys. Animals 2021, 11, 176. [Google Scholar] [CrossRef]
- IBM Corp. IBM SPSS Statistics Algorithms; IBM Corp: Armonk, NY, USA, 2017; p. 110. [Google Scholar]
- IBM Corp. IBM SPSS Statistics for Windows, 25.0; IBM Corp: Armonk, NY, USA, 2017. [Google Scholar]
- Bayarri, M.J.; Berger, J.O.; Forte, A.; García-Donato, G. Criteria for Bayesian model choice with application to variable selection. Ann. Stat. 2012, 40, 1550–1577. [Google Scholar] [CrossRef] [Green Version]
- Seifi-Jamadi, A.; Ahmad, E.; Ansari, M.; Kohram, H. Antioxidant effect of quercetin in an extender containing DMA or glycerol on freezing capacity of goat semen. Cryobiology 2017, 75, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Sikka, S.C.; Rajasekaran, M.; Hellstrom, W.J. Role of oxidative stress and antioxidants in male infertility. J. Androl. 1995, 16, 464–468. [Google Scholar]
- Flora, S.J. Structural, chemical and biological aspects of antioxidants for strategies against metal and metalloid exposure. Oxid. Med. Cell. Longev. 2009, 2, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Aboagla, E.M.-E.; Maeda, T. Arbutin‘s suppression of cryodamage in goat sperm and its mechanism of cryoprotection. Theriogenology 2011, 76, 538–546. [Google Scholar] [CrossRef]
- Zanganeh, Z.; Zhandi, M.; Zare-Shahneh, A.; Najafi, A.; Nabi, M.M.; Mohammadi-Sangcheshmeh, A. Does rosemary aqueous extract improve buck semen cryopreservation? Small Rumin. Res. 2013, 114, 120–125. [Google Scholar] [CrossRef]
- Silva, E.C.B.; Cajueiro, J.F.P.; Silva, S.V.; Soares, P.C.; Guerra, M.M.P. Effect of antioxidants resveratrol and quercetin on in vitro evaluation of frozen ram sperm. Theriogenology 2012, 77, 1722–1726. [Google Scholar] [CrossRef]
- Salehi, B.; Martorell, M.; Arbiser, J.L.; Sureda, A.; Martins, N.; Maurya, P.K.; Sharifi-Rad, M.; Kumar, P.; Sharifi-Rad, J. Antioxidants: Positive or Negative Actors? Biomolecules 2018, 8, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rietjens, S.J.; Bast, A.; Haenen, G.R. New insights into controversies on the antioxidant potential of the olive oil antioxidant hydroxytyrosol. J. Agric. Food Chem. 2007, 55, 7609–7614. [Google Scholar] [CrossRef] [PubMed]
- Hashem, N.M.; Gonzalez-Bulnes, A.; Simal-Gandara, J. Polyphenols in Farm Animals: Source of Reproductive Gain or Waste? Antioxidants 2020, 9, 1023. [Google Scholar] [CrossRef] [PubMed]
- Lancellotti, T.E.S.; Boarelli, P.V.; Romero, A.A.; Funes, A.K.; Cid-Barria, M.; Cabrillana, M.E.; Monclus, M.A.; Simón, L.; Vicenti, A.E.; Fornes, M.W. Semen quality and sperm function loss by hypercholesterolemic diet was recovered by addition of olive oil to diet in rabbit. PLoS ONE 2013, 8, e52386. [Google Scholar] [CrossRef] [PubMed]
- Mansour, S.W.; Sangi, S.; Harsha, S.; Khaleel, M.A.; Ibrahim, A. Sensibility of male rats fertility against olive oil, Nigella sativa oil and pomegranate extract. Asian Pac. J. Trop. Biomed. 2013, 3, 563–568. [Google Scholar] [CrossRef] [Green Version]
- Banihani, S.A. Semen quality as affected by olive oil. Int. J. Food Prop. 2017, 20, 1901–1906. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Gupta, S.; Sharma, R.K. Role of oxidative stress in female reproduction. Reprod. Biol. Endocrin. 2005, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, S.F.; Osman, K.; Das, S.; Othman, A.M.; Majid, N.A.; Rahman, M.P.A. A study of the antioxidant effect of alpha lipoic acids on sperm quality. Clinics 2008, 63, 545–550. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B. The antioxidant paradox: Less paradoxical now? Br. J. Clin. Pharmacol. 2013, 75, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Rahmatzadeh, M.; Kohram, H.; Zare Shahneh, A.; Seifi-Jamadi, A.; Ahmad, E. Antioxidative effect of BHA in soya bean lecithin-based extender containing glycerol or DMSO on freezing capacity of goat semen. Reprod. Domest. Anim. 2017, 52, 985–991. [Google Scholar] [CrossRef]
- Ahmad, E.; Aksoy, M.; Serin, İ.; Küçük, N.; Ceylan, A.; Uçan, U. Cholesterol-loaded cyclodextrin pretreatment of ram spermatozoa protects structural integrity of plasma membrane during osmotic challenge and reduces their ability to undergo acrosome reaction in vitro. Small Rumin. Res. 2013, 115, 77–81. [Google Scholar] [CrossRef]
- Daramola, J.O.; Adekunle, E.O.; Oke, O.E.; Onagbesan, O.M.; Iyasere, O.S.; Williams, T.J.; James, I.J.; Oyewusi, I.K.; Oyewusi, J.A. Effects of pyridoxine supplementation or in combination with other antioxidants on motility, in vitro capacitation and acrosome reaction of goat buck spermatozoa during cryopreservation. Small Rumin. Res. 2015, 131, 113–117. [Google Scholar] [CrossRef]
- Falchi, L.; Pau, S.; Pivato, I.; Bogliolo, L.; Zedda, M.T. Resveratrol supplementation and cryopreservation of buck semen. Cryobiology 2020, 95, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Huang, T., Jr.; Yanagimachi, R. Inner acrosomal membrane of mammalian spermatozoa: Its properties and possible functions in fertilization. Am. J. Anat. 1985, 174, 249–268. [Google Scholar] [CrossRef]
- Zadeh Hashem, E.; Haddad, R.; Eslami, M. Evaluation of ram semen enrichment with oleic acid on different spermatozoa parameters during low temperature liquid storage. Small Rumin. Res. 2017, 150, 30–39. [Google Scholar] [CrossRef]
- Gangwar, C.; Kharche, S.D.; Ranjan, R.; Kumar, S.; Goel, A.K.; Jindal, S.K.; Agarwal, S.K. Effect of vitamin C supplementation on freezability of Barbari buck semen. Small Rumin. Res. 2015, 129, 104–107. [Google Scholar] [CrossRef]
- Agarwal, A.; Virk, G.; Ong, C.; du Plessis, S.S. Effect of oxidative stress on male reproduction. World J. Mens Health 2014, 32, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Zhang, Y.; Bai, H.; Liu, J.; Li, J.; Wu, B. Mitochondria-targeted antioxidant MitoTEMPO improves the post-thaw sperm quality. Cryobiology 2018, 80, 26–29. [Google Scholar] [CrossRef]
- Sharafi, M.; Zhandi, M.; Akbari Sharif, A. Supplementation of soybean lecithin-based semen extender by antioxidants: Complementary flowcytometric study on post-thawed ram spermatozoa. Cell Tissue Bank. 2015, 16, 261–269. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Viability (%) | Acrosome Integrity (%) | HMMP (%) | LPO (%) | |
|---|---|---|---|---|
| FRESH | 84.4 ± 4.5 | 72.0 ± 6.4 | 79.8 ± 7.1 | 1.9 ± 0.8 |
| CONTROL | 43.9 ± 7.2 | 43.0 ± 7.7 | 40.3 ± 6.6 | 2.1 ± 0.5 |
| HT1 | 50.2 ± 6.5 | 40.2 ± 9.8 | 32.8 ± 4.4 | 1.7 ± 0.6 |
| HT2 | 44.5 ± 5.9 | 37.7 ± 5.5 | 32.6 ± 7.0 | 2.2 ± 0.4 |
| HT3 | 44.5 ± 8.8 | 42.2 ± 11.4 | 38.2 ± 5.6 | 2.5 ± 0.7 |
| HT4 | 43.1 ± 9.0 | 42.3 ± 6.9 | 40.3 ± 7.1 | 2.2 ± 0.8 |
| DHPG1 | 49.9 ± 14.2 | 47.5 ± 11.3 | 40.8 ± 9.0 | 2.2 ± 1.3 |
| DHPG2 | 43.8 ± 6.7 | 41.7 ± 6.4 | 35.8 ± 8.2 | 1.6 ± 0.5 |
| DHPG3 | 43.6 ± 7.0 | 38.8 ± 6.2 | 36.8 ± 9.1 | 1.8 ± 0.9 |
| DHPG4 | 47.0 ± 8.5 | 35.0 ± 8.3 | 40.0 ± 6.0 | 1.3 ± 0.7 |
| MIX1 | 43.4 ± 7.9 | 39.0 ± 5.2 | 35.8 ± 7.3 | 2.9 ± 1.5 |
| MIX2 | 44.2 ± 8.6 | 39.9 ± 7.9 | 36.5 ± 7.3 | 1.6 ± 0.7 |
| MIX3 | 42.6 ± 13.3 | 41.7 ± 12.7 | 38.0 ± 3.1 | 1.6 ± 0.6 |
| MIX4 | 43.2 ± 8.5 | 38.8 ± 8.9 | 34.0 ± 8.1 | 1.6 ± 0.7 |
| MT (%) | PM (%) | VLC (μm/s) | VSL (μm/s) | VAP (%) | LIN (%) | STR (%) | WOB (%) | ALH (μm) | BFC (Hz) | |
|---|---|---|---|---|---|---|---|---|---|---|
| FRESH | 91.6 ± 3.4 | 38.9 ± 4.8 | 105.4 ± 9.4 | 36.9 ± 4.9 | 64.3 ± 4.3 | 35.2 ± 4.7 | 57.4 ± 5.7 | 61.1 ± 3.3 | 3.3 ± 0.4 | 10.1 ± 1.1 |
| CONTROL | 60.2 ± 9.4 | 31.6 ± 6.5 | 82.5 ± 14.3 | 39.7 ± 8.9 | 54.5 ± 11.7 | 47.7 ± 3.8 | 72.7 ± 2.2 | 65.7 ± 4.0 | 3.0 ± 0.3 | 11.0 ± 0.6 |
| HT1 | 55.7 ± 6.4 | 30 ± 8.5 | 78.1 ± 10.0 | 37.6 ± 7.8 | 51.2 ± 9.0 | 48.0 ± 7.1 | 73.1 ± 4.7 | 65.4 ± 6.6 | 2.8 ± 0.3 | 11.4 ± 1.0 |
| HT2 | 60.0 ± 8.9 | 35.2 ± 5.3 | 79.7 ± 6.8 | 38.9 ± 4.9 | 52.3 ± 5.2 | 49.0 ± 6.7 | 74.3 ± 4.2 | 65.8 ± 6.3 | 2.9 ± 0.4 | 11.2 ± 1.0 |
| HT3 | 63.7 ± 8.1 | 33.9 ± 5.8 | 77.0 ± 10.2 | 36.1 ± 6.4 | 49.6 ± 9.1 | 46.9 ± 5.6 | 73.0 ± 3.2 | 64.2 ± 6.6 | 2.9 ± 0.3 | 11.1 ± 1.2 |
| HT4 | 54.5 ± 7.9 | 27.7 ± 8.2 | 85.7 ± 9.2 | 36.2 ± 4.6 | 52.9 ± 6.8 | 42.2 ± 2.9 | 68.5 ± 3.1 | 61.6 ± 2.6 | 3.0 ± 0.2 | 11.9 ± 0.2 |
| DHPG1 | 58.9 ± 9.8 | 32.8 ± 8 | 83.7 ± 10.8 | 37.6 ± 3.3 | 52.8 ± 5.4 | 45.3 ± 3.9 | 71.4 ± 2.3 | 63.3 ± 3.8 | 3.1 ± 0.3 | 12.1 ± 0.9 |
| DHPG2 | 55.5 ± 9.8 | 31.8 ± 4.7 | 85.0 ± 10.0 | 39.8 ± 5.6 | 55.1 ± 8.3 | 46.8 ± 3.2 | 72.5 ± 2.3 | 64.6 ± 3.7 | 2.9 ± 0.2 | 11.8 ± 0.4 |
| DHPG3 | 61.6 ± 9.1 | 32.4 ± 7.9 | 78.5 ± 12.4 | 35.2 ± 5.1 | 49.0 ± 6.2 | 45.2 ± 4.9 | 71.9 ± 4.4 | 62.7 ± 3.4 | 3.0 ± 0.3 | 11.6 ± 1.0 |
| DHPG4 | 59.7 ± 9.8 | 33.9 ± 4.4 | 84.9 ± 8.5 | 37.9 ± 3.6 | 52.5 ± 4.7 | 44.7 ± 3.1 | 72.2 ± 2.7 | 62.0 ± 4.0 | 3.1 ± 0.3 | 12.2 ± 1.1 |
| MIX1 | 59.8 ± 9.4 | 30.7 ± 8.3 | 81.0 ± 10.7 | 36.3 ± 6.0 | 50.7 ± 6.4 | 44.9 ± 4.3 | 71.4 ± 4.1 | 62.8 ± 2.9 | 3.0 ± 0.2 | 11.4 ± 1.3 |
| MIX2 | 60.0 ± 6.6 | 31.5 ± 7.8 | 80.1 ± 6.6 | 35.3 ± 4.0 | 49.6 ± 3.5 | 44.1 ± 2.8 | 71.1 ± 3.5 | 62.0 ± 1.6 | 3.1 ± 0.2 | 11.4 ± 0.9 |
| MIX3 | 57.4 ± 9.0 | 32.3 ± 9.1 | 76.8 ± 5.5 | 34.4 ± 3.1 | 47.2 ± 4.2 | 44.8 ± 3.5 | 72.9 ± 5.0 | 61.4 ± 2.0 | 3.0 ± 0.2 | 11.7 ± 1.0 |
| MIX4 | 58.1 ± 8.1 | 31.4 ± 3.6 | 77.3 ± 5.5 | 34.5 ± 2.6 | 47.5 ± 3.9 | 44.6 ± 2.0 | 72.7 ± 1.9 | 61.4 ± 2.8 | 3.1 ± 0.1 | 11.9 ± 0.7 |
| BG Sum of Squares | BG df | BG Mean Square | WG Sum of Squares | WG df | WG Mean Square | F | p Value | Bayes Factor | |
|---|---|---|---|---|---|---|---|---|---|
| Viability (%) | 9146.527 | 12 | 703.579 | 5301.865 | 70 | 75.741 | 9.289 | 0.001 | 14,617,034.109 |
| Acrosome integrity (%) | 6166.972 | 12 | 474.382 | 5044.732 | 70 | 72.068 | 6.582 | 0.001 | 11,102.852 |
| HMMP (%) | 10,748.051 | 12 | 826.773 | 3465.358 | 70 | 49.505 | 16.701 | 0.001 | 24,196,033,653,804.700 |
| LPO (%) | 13.991 | 12 | 1.076 | 48.149 | 70 | 0.688 | 1.565 | 0.117 | 0.000 |
| TM (%) | 6445.744 | 12 | 495.826 | 4985.240 | 70 | 71.218 | 6.962 | 0.001 | 33,431.056 |
| PM (%) | 530.067 | 12 | 40.774 | 3293.809 | 70 | 47.054 | 0.867 | 0.591 | 0.000 |
| VCL (μm/s) | 4134.502 | 12 | 318.039 | 6422.702 | 70 | 91.753 | 3.466 | 0.001 | 0.252 |
| VSL (μm/s) | 247.939 | 12 | 19.072 | 1998.821 | 70 | 28.555 | 0.668 | 0.787 | 0.000 |
| VAP (μm/s) | 1402.105 | 12 | 107.854 | 3178.839 | 70 | 45.412 | 2.375 | 0.011 | 0.002 |
| LIN (%) | 869.610 | 12 | 66.893 | 1361.286 | 70 | 19.447 | 3.440 | 0.001 | 0.227 |
| STR (%) | 1344.017 | 12 | 103.386 | 960.978 | 70 | 13.728 | 7.531 | 0.001 | 163,617.988 |
| WOB (%) | 222.771 | 12 | 17.136 | 1195.420 | 70 | 17.077 | 1.003 | 0.458 | 0.000 |
| ALH (μm) | 0.842 | 12 | 0.065 | 4.724 | 70 | 0.067 | 0.960 | 0.499 | 0.000 |
| BCF (Hz) | 23.095 | 12 | 1.777 | 60.923 | 70 | 0.870 | 2.041 | 0.029 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arando Arbulu, A.; Navas González, F.J.; Bermúdez-Oria, A.; Delgado Bermejo, J.V.; Fernández-Prior, Á.; González Ariza, A.; León Jurado, J.M.; Pérez-Marín, C.C. Bayesian Analysis of the Effects of Olive Oil-Derived Antioxidants on Cryopreserved Buck Sperm Parameters. Animals 2021, 11, 2032. https://doi.org/10.3390/ani11072032
Arando Arbulu A, Navas González FJ, Bermúdez-Oria A, Delgado Bermejo JV, Fernández-Prior Á, González Ariza A, León Jurado JM, Pérez-Marín CC. Bayesian Analysis of the Effects of Olive Oil-Derived Antioxidants on Cryopreserved Buck Sperm Parameters. Animals. 2021; 11(7):2032. https://doi.org/10.3390/ani11072032
Chicago/Turabian StyleArando Arbulu, Ander, Francisco Javier Navas González, Alejandra Bermúdez-Oria, Juan Vicente Delgado Bermejo, África Fernández-Prior, Antonio González Ariza, Jose Manuel León Jurado, and Carlos Carmelo Pérez-Marín. 2021. "Bayesian Analysis of the Effects of Olive Oil-Derived Antioxidants on Cryopreserved Buck Sperm Parameters" Animals 11, no. 7: 2032. https://doi.org/10.3390/ani11072032
APA StyleArando Arbulu, A., Navas González, F. J., Bermúdez-Oria, A., Delgado Bermejo, J. V., Fernández-Prior, Á., González Ariza, A., León Jurado, J. M., & Pérez-Marín, C. C. (2021). Bayesian Analysis of the Effects of Olive Oil-Derived Antioxidants on Cryopreserved Buck Sperm Parameters. Animals, 11(7), 2032. https://doi.org/10.3390/ani11072032