
Environmental Streptococcus uberis Associated with Clinical Mastitis in Dairy Cows: Virulence Traits, Antimicrobial and Biocide Resistance, and Epidemiological Typing
 ,
,  ,
,  , and
, and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Lactating Cows and Husbandry Practices
2.2. Milk Sampling and Isolation of S. uberis
2.3. Antimicrobial Susceptibility Testing
2.4. DNA Extraction and Molecular Identification of S. uberis
2.5. PCR Amplifications of Virulence Attributes and Antimicrobial and Biocide Resistance Genes
2.6. PCR–RFLP
2.7. Bioinformatics and Statistical Analysis
3. Results
3.1. Infection Rate and Characterization of S. uberis in Clinically Mastitic Dairy Cows
3.2. Antimicrobial Resistance Patterns of S. uberis Isolates
3.3. Molecular Characteristics and Virulence Gene Profiling of S. uberis
3.4. Detection of Antimicrobial Resistance Genes in S. uberis Isolates
3.5. Biocide Resistance Genes and Biocide–Antimicrobial Cross-Resistance
3.6. Typing of Virulent S. uberis Isolates Using RFLP–PCR
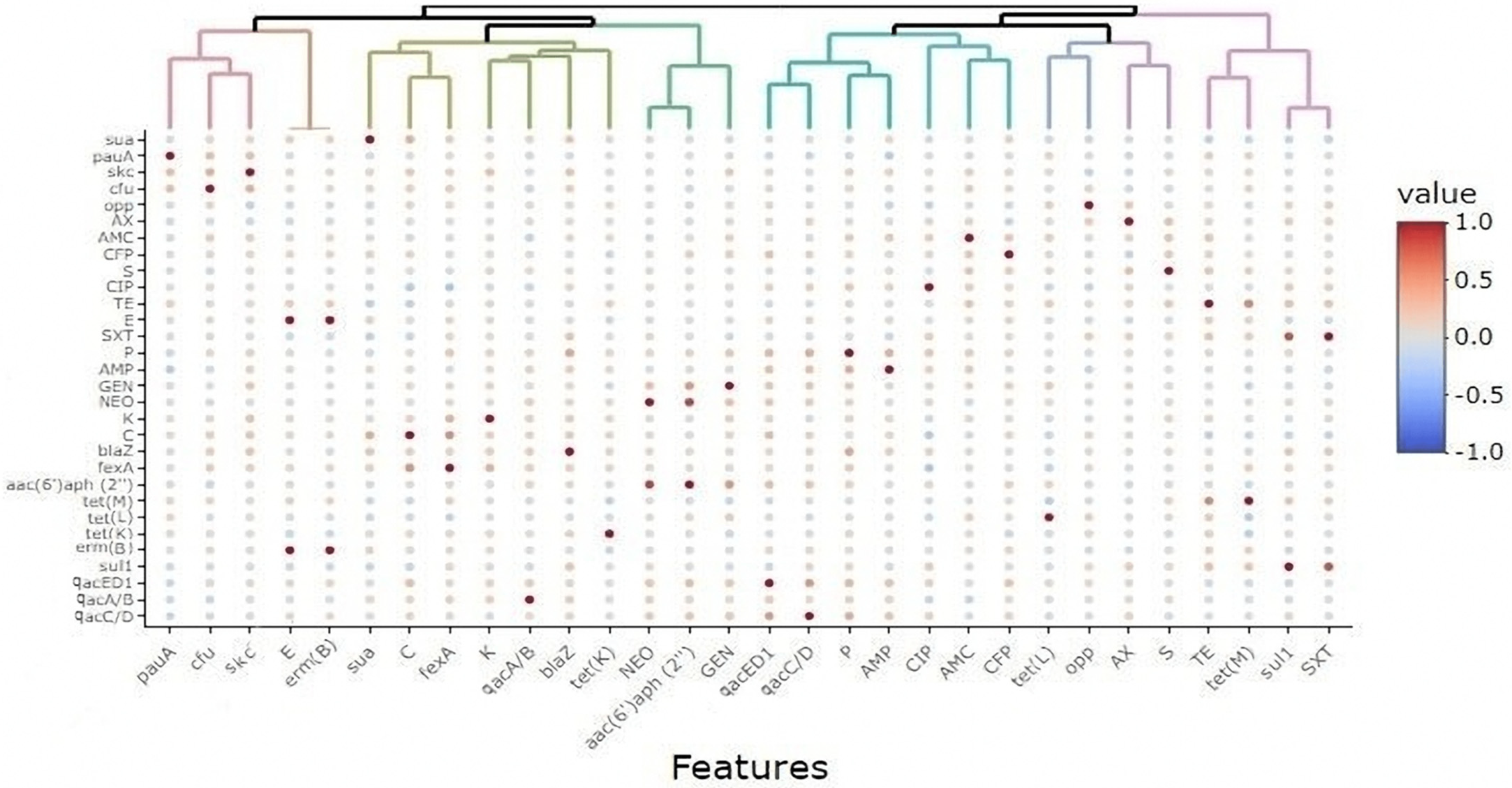
3.7. Association between the Existence of Virulence Traits, Antimicrobial and Biocide Resistance, and Hygienic Interventions for Dairy Cows
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Collenburg, L.; Beyersdorf, N.; Wiese, T.; Arenz, C.; Saied, E.M.; Becker-Flegler, K.A.; Schneider-Schaulies, S.; Avota, E. The Activity of the Neutral Sphingomyelinase Is Important in T Cell Recruitment and Directional Migration. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Abebe, R.; Hatiya, H.; Abera, M.; Megersa, B.; Asmare, K. Bovine mastitis: Prevalence, risk factors and isolation of Staphylococcus aureus in dairy herds at Hawassa milk shed, South Ethiopia. BMC Vet. Res. 2016, 12, 270. [Google Scholar] [CrossRef]
- Lone, M.A.; Hülsmeier, A.J.; Saied, E.M.; Karsai, G.; Arenz, C.; von Eckardstein, A.; Hornemann, T. Subunit Composition of the Mammalian Serine-Palmitoyltransferase Defines the Spectrum of Straight and Methyl-Branched Long-Chain Bases. Proc. Natl. Acad. Sci. USA 2020, 117, 15591–15598. [Google Scholar] [CrossRef] [PubMed]
- Käppeli, N.; Morach, M.; Zurfluh, K.; Corti, S.; Nüesch-Inderbinen, M.; Stephan, R. Sequence types and antimicrobial resistance profiles of Streptococcus uberis isolated from bovine mastitis. Front. Vet. Sci. 2019, 6, 234. [Google Scholar] [CrossRef]
- Vasiliauskaité-Brooks, I.; Healey, R.D.; Rochaix, P.; Saint-Paul, J.; Sounier, R.; Grison, C.; Waltrich-Augusto, T.; Fortier, M.; Hoh, F.; Saied, E.M.; et al. Structure of a Human Intramembrane Ceramidase Explains Enzymatic Dysfunction Found in Leukodystrophy. Nat. Commun. 2018, 9, 5437. [Google Scholar] [CrossRef] [PubMed]
- Facklam, R. What happened to the streptococci: Overview of taxonomic and nomenclature changes. Clin. Microbiol. Rev. 2002, 15, 613–630. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.U.; Hassan, A.A.; Abdulmawjood, A.; Lanimler, C.; Wolter, W.; Zschock, M. Identification and epidemiological characterization of Streptococcus uberis isolated from bovine mastitis using conventional methods. J. Vet. Sci. 2003, 4, 213–224. [Google Scholar] [CrossRef]
- Almeida, R.A.; Luther, D.A.; Park, H.M.; Oliver, S.P. Identification, isolation and partial characterization of a novel Streptococcus uberis adhesion molecule (SUAM). Vet. Microbiol. 2006, 115, 183–191. [Google Scholar] [CrossRef]
- Smith, A.J.; Kitt, A.J.; Ward, P.N.; Leigh, J.A. Isolation and characterization of a mutant strain of Streptococcus uberis, which fails to utilize a plasmin derived beta-casein peptide for the acquisition of methionine. J. Appl. Microbiol. 2002, 93, 631–639. [Google Scholar] [CrossRef]
- Ward, P.N.; Leigh, J.A. Genetic analysis of Streptococcus uberis plasminogen activators. Indian J. Med. Res. 2004, 119, 136–140. [Google Scholar]
- Reinoso, E.B.; Lasagno, M.C.; Dieser, S.A.; Odierno, L.M. Distribution of virulence-associated genes in Streptococcus uberis isolated from bovine mastitis. FEMS Microbiol. Lett. 2011, 318, 183–188. [Google Scholar] [CrossRef]
- Palma, E.; Tilocca, B.; Roncada, P. Antimicrobial resistance in veterinary medicine: An overview. Int. J. Mol. Sci. 2020, 21, 1914. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Aziz, N.K.; Ammar, A.M.; El-Naenaeey, E.Y.M.; El Damaty, H.M.; Elazazy, A.A.; Hefny, A.A.; Shaker, A.; Eldesoukey, I.E. Antimicrobial and antibiofilm potentials of cinnamon oil and silver nanoparticles against Streptococcus agalactiae isolated from bovine mastitis: New avenues for countering resistance. BMC Vet. Res. 2021, 17, 1–14. [Google Scholar] [CrossRef]
- Tomazi, T.; Freu, G.; Alves, B.G.; de Souza Filho, A.F.; Heinemann, M.B.; Veiga Dos Santos, M. Genotyping and antimicrobial resistance of Streptococcus uberis isolated from bovine clinical mastitis. PLoS ONE 2019, 14, e0223719. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, F.; Li, X.-P.; Luo, J.-Y.; Wang, L.; Zhou, Y.-L.; Yan, Y.; Wang, X.-R.; Li, H.S. Detection of antimicrobial resistance and virulence-related genes in Streptococcus uberis and Streptococcus parauberis isolated from clinical bovine mastitis cases in northwestern China. J. Integr. Agric. 2020, 19, 2784–2791. [Google Scholar] [CrossRef]
- Pitkälä, A.; Koort, J.; Björkroth, J. Identification and antimicrobial resistance of Streptococcus uberis and Streptococcus parauberis isolated from bovine milk samples. J. Dairy Sci. 2008, 91, 4075–4081. [Google Scholar] [CrossRef]
- Roussel, P.; Porcherie, A.; Répérant-Ferter, M.; Cunha, P.; Gitton, C.; Rainard, P.; Germon, P. Escherichia coli mastitis strains: In Vitro phenotypes and severity of infection In Vivo. PLoS ONE 2017, 12, e0178285. [Google Scholar] [CrossRef]
- Abd El-Aziz, N.K.; Abd El-Hamid, M.I.; Bendary, M.M.; El-Azazy, A.A.; Ammar, A.M. Existence of vancomycin resistance among methicillin resistant S. aureus recovered from animal and human sources in Egypt. Slov. Vet. Res. 2018, 55, 221–230. [Google Scholar]
- Bjorland, J.; Steinum, T.; Kvitle, B.; Waage, S.; Sunde, M.; Heir, E. Widespread distribution of disinfectant resistance genes among staphylococci of bovine and caprine origin in Norway. J. Clin. Microbiol. 2005, 43, 4363–4368. [Google Scholar] [CrossRef]
- Quinn, P.J.; Carter, M.E.; Markey, B.; Carter, G.R. Clinical Veterinary Microbiology; Mosby: London, UK, 1999; pp. 21–66. [Google Scholar]
- Hardie, J.M. Genus Streptococcus. In Bergey’s Manual of Systematic Bacteriology; Sneath, P.H.A., Ed.; Williams and Wilkins: Baltimore, MD, USA, 1986; Volume 2, pp. 1043–1071. [Google Scholar]
- Christie, R.; Atkins, N.E.; Munch-Petersen, E. A note on a lytic phenomenon shown by group B streptococci. Aust. J. Exp. Biol. Med. Sci. 1944, 22, 197–200. [Google Scholar] [CrossRef]
- Domig, K.J.; Mayer, H.K.; Kneifel, W. Methods used for the isolation, enumeration, characterization and identification of Enterococcus spp. 2. Pheno- and genotypic criteria. Int. J. Food Microbiol. 2003, 88, 165–188. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standard single disc method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standard for Antimicrobial Susceptibility Testing: Nineteenth Informational Supplement M100; CLSI: Wayne, PA, USA, 2019. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Tambekar, D.; Dhanorkar, D.; Gulhane, S.; Khandelwal, V.; Dudhane, M. Antibacterial susceptibility of some urinary tract pathogens to commonly used antibiotics. Afr. J. Biotechnol. 2006, 5, 1562–1565. [Google Scholar]
- Picard, F.J.; Ke, D.; Boudreau, D.K.; Boissinot, M.; Huletsky, A.; Richard, D.; Ouellette, M.; Roy, P.H.; Bergeron, M.G. Use of tuf sequences for genus-specific PCR detection and phylogenetic analysis of 28 Streptococcal species. J. Clin. Microbiol. 2004, 42, 3686–3695. [Google Scholar] [CrossRef]
- Nithin Prabhu, K.; Isloor, S.; Hegde, R.; Rathnamma, D.; Veeregowda, B.M.; Narasimha Murthy, H.; Shome, R.; Suryanarayana, V.V.S. Development of polymerase chain reaction for detection of predominant streptococcal isolates causing subclinical bovine mastitis. Indian J. Biotechnol. 2013, 12, 208–212. [Google Scholar]
- Nithin Prabhu, K.; Isloor, S.K.; Hegde, R.; Suryanarayana, W. Standardization of PCR and phylogenetic analysis for predominant streptococcal species isolated from subclinical mastitis. In Proceedings of the International Symposium on “Role of Biotechnology in Conserving Biodiversity and Livestock Development for Food Security and Poverty Alleviation” and XVII Annual Convention of Indian Society of Veterinary Immunology & Biotechnology (ISVIB), Bikaner, India, 23 March 2010; Volume 50, p. 47. [Google Scholar]
- Vesterholm-Nielsen, M.; Ølholm Larsen, M.; Elmerdahl Olsen, J.; Møller Aarestrup, E. Occurrence of the blaZ Gene in penicillin resistant Staphylococcus aureus isolated from bovine mastitis in Denmark. Acta Vet. Scand. 1999, 40, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Kehrenberg, C.; Schwarz, S. Distribution of florfenicol resistance genes fexA and cfr among chloramphenicol resistant Staphylococcus isolates. Antimicrob. Agents Chemother. 2006, 50, 1156–1163. [Google Scholar] [CrossRef] [PubMed]
- Duran, N.; Ozer, B.; Duran, G.G.; Onlen, Y.; Demir, C. Antibiotic resistance genes & susceptibility patterns in staphylococci. Indian J. Med. Res. 2012, 135, 389–396. [Google Scholar]
- Ng, L.K.; Martin, I.; Alfa, M.; Mulvey, M. Multiplex PCR for the detection of tetracycline resistant genes. Mol. Cell Probes. 2001, 15, 209–215. [Google Scholar] [CrossRef]
- Malhotra-Kumar, S.; Lammens, C.; Piessens, J.; Goossens, H. Multiplex PCR for simultaneous detection of macrolide and tetracycline resistance determinants in streptococci. Antimicrob. Agents Chemother. 2005, 49, 4798–4800. [Google Scholar] [CrossRef]
- Lina, G.; Quaglia, A.; Reverdy, M.E.; Leclercq, R.; Vandenesch, F.; Etienne, J. Distribution of genes encoding resistance to macrolides, lincosamides, and streptogramins among staphylococci. Antimicrob. Agents Chemother. 1999, 43, 1062–1066. [Google Scholar] [CrossRef] [PubMed]
- Schlegelova, J.; Vlkova, H.; Babak, V.; Holasova, M.; Jaglic, Z.; Stosova, T.; Sauer, P. Resistance to erythromycin of Staphylococcus spp. isolates from the food chain. Vet. Med. 2008, 53, 307–314. [Google Scholar] [CrossRef]
- Jensen, L.B.; Frimodt-Møller, N.; Aarestrup, F.M. Presence of erm gene classes in gram-positive bacteria of animal and human origin in Denmark. FEMS Microbiol. Lett. 1999, 170, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Ibekwe, A.M.; Murinda, S.E.; Graves, A.K. Genetic diversity and antimicrobial resistance of Escherichia coli from human and animal sources uncovers multiple resistances from human sources. PLoS ONE 2011, 6, e20819. [Google Scholar] [CrossRef] [PubMed]
- Grape, M.; Motakefi, A.; Pavuluri, S.; Kahlmeter, G. Standard and real-time multiplex PCR methods for detection of trimethoprim resistance dfr genes in large collections of bacteria. Clin. Microbiol. Infect. 2007, 13, 1112–1118. [Google Scholar] [CrossRef]
- Noguchi, N.; Suwa, J.; Narui, K.; Sasatsu, M.; Ito, T.; Hiramatsu, K.; Song, J. Susceptibilities to antiseptic agents and distribution of antiseptic-resistance genes qacA/B and smr of methicillin-resistant Staphylococcus aureus isolated in Asia during 1998 and 1999. J. Med. Microbiol. 2005, 54, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Chuanchuen, R.; Khemtong, S.; Padungtod, P. Occurrence of qacE/qacEDelta1 genes and their correlation with class 1 integrons in Salmonella enterica isolates from poultry and swine. Southeast Asian J. Trop. Med. Public Health. 2007, 38, 855–862. [Google Scholar]
- Kolde, R. Package ‘Pheatmap’: Pretty Heat Map. 2018. Available online: https://rdrr.io/cran/pheatmap/ (accessed on 12 May 2020).
- Friendly, M. Corrgrams: Exploratory displays for correlation matrices. Am. Stat. 2002, 56, 316–324. [Google Scholar] [CrossRef]
- Galili, T.; O’Callaghan, A.; Sidi, J.; Sievert, C. Heatmaply: An R package for creating interactive cluster heatmaps for online publishing. Bioinformatics 2018, 34, 1600–1602. [Google Scholar] [CrossRef]
- Harrel, F.E., Jr. Package ‘Hmisc’. Available online: https://cran.r-project.org/web/packages/Hmisc/Hmisc.pdf (accessed on 4 May 2020).
- SAS. SAS Statistics User’s Guide. Statistical Analytical System, 5th rev. SAS ed.; Institute Inc.: Cary, NC, USA, 2012. [Google Scholar]
- Palanivel, K.M.; Suresh, R.V.; Jayakumar, R.; Ganesan, P.I.; Dhanapalan, P. Retrospective study of sub-clinical mastitis in buffaloes. Indian J. Vet. Med. 2008, 28, 34–36. [Google Scholar]
- El Damaty, H.M.; Mahmmod, Y.S.; Gouda, S.M.; Sobhy, N.M. Epidemiological and ultrasonographic investigation of bovine fascioliasis in smallholder production system in Eastern Nile Delta of Egypt. Prev. Vet. Med. 2018, 158, 35–42. [Google Scholar] [CrossRef]
- El Damaty, H.M.; Fawzi, E.M.; Neamat-Allah, A.N.F.; Elsohaby, I.; Abdallah, A.; Farag, G.K.; El-Shazly, Y.A.; Mahmmod, Y.S. Characterization of foot and mouth disease virus serotype SAT-2 in swamp water buffaloes (Bubalus bubalis) under the Egyptian smallholder production system. Animals 2021, 11, 1697. [Google Scholar] [CrossRef]
- Bradley, A.J.; Leach, K.A.; Breen, J.E.; Green, L.E.; Green, M.J. Survey of the incidence and etiology of mastitis in dairy’ farms in England and Wales. Vet. Rec. 2007, 160, 253–257. [Google Scholar] [CrossRef]
- Shome, B.R.; Bhuvana, M.; Mitra, S.D.; Krithiga, N.; Shome, R.; Velu, D.; Banerjee, A.; Barbuddhe, S.B.; Prabhudas, K.; Rahman, H. Molecular characterization of Streptococcus agalactiae and Streptococcus uberis isolates from bovine milk. Trop. Anim. Health Prod. 2012, 44, 1981–1992. [Google Scholar] [CrossRef]
- Eldesouky, I.E.; Refae, M.A.; Nada, H.S.; Elnaby, G.R.H. Molecular detection of Streptococcus species isolated from cows with mastitis. World Vet. J. 2016, 6, 193–202. [Google Scholar] [CrossRef]
- Amin, B.; Deneke, Y.; Abdela, N. Bovine mastitis: Prevalence, risk factors, and isolation of Streptococcus species from small holders dairy farms in and around Haramaya town, Eastern Ethiopia. Glob. J. Med. Res. 2017, 17, 27–38. [Google Scholar]
- Amosun, E.A.; Ajuwape, A.T.P.; Adetosoye, A.I. Bovine streptococcal mastitis in Southwest and Northern States of Nigeria. Afr. J. Biomed. Res. 2010, 13, 33–37. [Google Scholar]
- Vasiľ, M. Etiology, course, and reduction of incidence of environmental mastitis in the herd of dairy cows. Slovak J. Anim. Sci. 2009, 3, 136–144. [Google Scholar]
- Abozaid, A.A.; El Balkemy, F.A.; El Damaty, H.M. Impact of risk factors on the prevalence of mastitis in dairy cattle. Zag. Vet. J. 2013, 41, 162–168. [Google Scholar] [CrossRef]
- Jayarao, B.M.; Pillai, S.R.; Sawant, A.A.; Wolfgang, D.R.; Hegde, N.V. Guidelines for monitoring bulk tank milk somatic cell and bacterial counts. J. Dairy Sci. 2004, 87, 3561–3573. [Google Scholar] [CrossRef]
- Kivaria, F.M.; Noordhuizen, J.P.T.M.; Msamia, H.M. Risk factors associated with the incidence rate of clinical mastitis in smallholder dairy cows in the Dar es Salaam region of Tanzania. Vet. J. 2007, 173, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Haenni, M.; Galofaro, L.; Ythier, M.; Giddey, M.; Majcherczyk, P.; Moreillon, P.; Madec, J.Y. Penicillin-bindingprotein gene alterations in Streptococcus uberis isolates presenting decreased susceptibility to penicillin. Antimicrob. Agents Chemother. 2010, 54, 1140–1145. [Google Scholar] [CrossRef]
- Cameron, M.; Saab, M.; Heider, L.; McClure, J.T.; Rodriguez-Lecompte, J.C.; Sanchez, J. Antimicrobial susceptibility patterns of environmental streptococci recovered from bovine milk samples in the Maritime Provinces of Canada. Front. Vet. Sci. 2016, 3, 1–14. [Google Scholar] [CrossRef]
- Erskine, R.J.; Walker, R.D.; Bolin, C.A.; Bartlett, P.C.; White, D.G. Trends in antibacterial susceptibility of mastitis pathogens during a seven-year period. J. Dairy Sci. 2002, 85, 1111–1118. [Google Scholar] [CrossRef]
- Gianneechini, R.E.; Concha, C.; Franklin, A. Antimicrobial susceptibility of udder pathogens isolated from dairy herds in the West Littoral region of Uruguay. Acta Vet. Scand. 2002, 43, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Minst, K.; Märtlbauer, E.; Miller, T.; Meyer, C. Short communication: Streptococcus species isolated from mastitis milk samples in Germany and their resistance to antimicrobial agents. J. Dairy Sci. 2012, 95, 6957–6962. [Google Scholar] [CrossRef] [PubMed]
- Schröder, A.; Hoedemacker, M.; Klein, G. Susceptibility of mastitis pathogens in northern Germany. Berl. Munch. Tierarztl. Wochenschr. 2005, 118, 393–398. [Google Scholar] [PubMed]
- Rato, M.G.; Bexiga, R.; Florindo, C.; Cavaco, L.M.; Vilela, C.L.; Santos-Sanches, I. Antimicrobial resistance and molecular epidemiology of streptococci from bovine mastitis. Vet. Microbiol. 2013, 161, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, H. Results of the antimicrobial agent susceptibility study raised in a representative, cross-sectional monitoring study on a national basis. Int. J. Med. Microbiol. 2006, 296, 69–79. [Google Scholar] [CrossRef]
- Lollai, S.A.; Ziccheddu, M.; Duprè, I.; Piras, D. Characterization of resistance to tetracyclines and aminoglycosides of sheep mastitis pathogens: Study of the effect of gene content on resistance. J. Appl. Microbiol. 2016, 121, 941–951. [Google Scholar] [CrossRef]
- Abd El-Aziz, N.K.; Gharib, A.A. Coexistence of plasmid-mediated quinolone resistance determinants and AmpC-beta- lactamases in Escherichia coli strains in Egypt. Cell Mol. Biol. 2015, 61, 29–35. [Google Scholar] [PubMed]
- Guérin-Faublée, V.; Tardy, F.; Bouveron, C.; Carret, G. Antimicrobial susceptibility of Streptococcus species isolated from clinical mastitis in dairy cows. Int. J. Antimicrob. Agents 2002, 19, 219–226. [Google Scholar] [CrossRef]
- Denamiel, G.; Llorente, P.; Carabella, M.; Rebuelto, M.; Gen-tilini, E. Anti-microbial susceptibility of Streptococcus spp. isolated from bovine mastitis in Argentina. J. Vet. Med. B Infect. Dis. Vet. Public Health 2005, 52, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Boireau, C.; Cazeau, G.; Jarrige, N.; Calavas, D.; Madec, J.Y.; Leblond, A.; Haenni, M.; Gay, É. Antimicrobial resistance in bacteria isolated from mastitis in dairy cattle in France, 2006–2016. J. Dairy Sci. 2018, 101, 9451–9462. [Google Scholar] [CrossRef]
- Abd El-Aziz, N.K.; Ammar, A.M.; Hamdy, M.M.; Gobouri, A.A.; Azab, E.; Sewid, A.H. First report of aacC5-aadA7_4 gene cassette array and phage tail tape measure protein on class 1 integrons of Campylobacter species isolated from animal and human sources in Egypt. Animals 2020, 10, 2067. [Google Scholar] [CrossRef]
- Lasagno, M.C.; Reinoso, E.B.; Dieser, S.A.; Calvinho, L.F.; Buzzola, F.; Vissio, C.; Bogni, C.; Odierno, L.M. Phenotypic and genotypic characterization of Streptococcus uberis isolated from bovine subclinical mastitis in Argentinean dairy farms. Rev. Argent. Microbiol. 2011, 43, 212–217. [Google Scholar]
- Zadoks, R.N.; Schukken, Y.H.; Wiedmann, M. Multilocus sequence typing of Streptococcus uberis provides sensitive and epidemiologically relevant subtype information and reveals positive selection in the virulence gene pauA. J. Clin. Microbiol. 2005, 43, 2407–2417. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ward, P.N.; Field, T.R.; Rapier, C.D.; Leigh, J.A. The activation of bovine plasminogen by pauA is not required for virulence of Streptococcus uberis. Infect. Immun. 2003, 71, 7193–7196. [Google Scholar] [CrossRef]
- Bragg, R.; Jansen, A.; Coetzee, M.; van der Westhuizen, W.; Boucher, C. Bacterial resistance to quaternary ammonium compounds (qac) disinfectants. In Infectious Diseases and Nanomedicine II. Advances in Experimental Medicine and Biology; Adhikari, R., Thapa, S., Eds.; Springer: New Delhi, India, 2014; Volume 808. [Google Scholar]
- Haggag, Y.N.; Nossair, M.A.; Mansour, A.M.; Abd El Rahman, A.H. Streptococci in dairy farms: Isolation, antibiogram pattern and disinfectant sensitivity. Alex. J. Vet. Sci. 2018, 59, 85–92. [Google Scholar] [CrossRef]
- Levy, S.B.; Marshall, B. Antibacterial resistance worldwide: Causes, challenges and responses. Nat. Med. 2004, 10, S122–S129. [Google Scholar] [CrossRef] [PubMed]
- Cambau, E.; Guillard, T. Antimicrobials that affect the synthesis and conformation of nucleic acids. Rev. Sci. Tech. 2012, 31, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Oggioni, M.R.; Coelho, J.R.; Furi, L.; Knight, D.R.; Viti, C.; Orefici, G.; Martinez, J.L.; Freitas, A.T.; Coque, T.M.; Morrissey, I.; et al. Significant differences characterise the correlation coefficients between biocide and antibiotic susceptibility profiles in Staphylococcus aureus. Curr. Pharm. Des. 2015, 21, 2054–2057. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Class | AMA | No. of Resistant Isolates (%) | MAR Index | Fisher Exact p-Value * |
|---|---|---|---|---|
| Beta-lactams | CX | 69 (100.00) | 0.048 | NE |
| AMP | 62 (89.85) | 0.043 | 0.007 | |
| AX | 48 (69.57) | 0.033 | <0.001 | |
| P | 55 (79.71) | 0.038 | <0.001 | |
| Beta-lactamase inhibitor | AMC | 24 (34.78) | 0.017 | <0.001 |
| Cephalosporins | CRO | 69 (100.00) | 0.048 | NE |
| CFP | 18 (26.09) | 0.012 | <0.001 | |
| CL | 69 (100.00) | 0.048 | NE | |
| FEP | 69 (100.00) | 0.048 | NE | |
| Non-beta Lactams (Carbapenems) | IPM | 0 (00.00) | 0.00 | <0.001 |
| Lincomycins | DA | 69 (100.00) | 0.048 | NE |
| Fluoroquinlones | CIP | 15 (21.74) | 0.010 | <0.001 |
| Tetracyclines | TE | 45 (65.22) | 0.031 | <0.001 |
| Macrolides | E | 51(73.91) | 0.035 | 0.001 |
| Aminoglycosides | S | 60 (86.96) | 0.041 | 0.002 |
| GEN | 14 (20.28) | 0.009 | <0.001 | |
| NEO | 28 (40.57) | 0.019 | <0.001 | |
| K | 21 (30.43) | 0.014 | <0.001 | |
| Phenicols | C | 38 (55.07) | 0.026 | <0.001 |
| Aminocoumarins | NV | 69 (100.00) | 0.048 | NE |
| Sulfonamides | SXT | 33 (47.83) | 0.023 | <0.001 |
| Isolate No. | Herd | Virulence Pattern | Antimicrobial Resistance Profile | Biocide Resistance Genes | |
|---|---|---|---|---|---|
| Phenotype | Resistance Genes | ||||
| 1 | A | ND | CX, CRO, CL, FEP, S, DA, TE, E, SXT, AMP, NV | tet(M), erm(B), sul1 | ND |
| 2 | pauA, cfu, opp | AX, CX, CRO, CL, FEP, S, DA, TE, E, SXT, AMP, NV, C | tet(L), erm(B) | qacED1 | |
| 3 | pauA, cfu, opp | AX, CX, CRO, CL, FEP, DA, TE, E, SXT, NV | tet(M), erm(B), sul1 | ND | |
| 4 | pauA, cfu, opp | AX, AMC, CX, CRO, CL, FEP, S, DA, TE, E, SXT, P, AMP, NV, K, C | blaZ, fexA, tet(M), sul1 | qacED1, qacC/D | |
| 5 | sua | AX, AMC, CX, CRO, CL, FEP, S, DA, E, P, AMP, NV | blaZ, erm(B) | ND | |
| 6 | pauA | CX, CRO, CL, FEP, S, DA, TE, NV | tet(M) | ND | |
| 7 | pauA | AX, CX, CRO, CL, FEP, S, DA, CIP, TE, SXT, P, AMP, NV, NEO | blaZ, aac(6′)aph(2″), tet(K), sul1 | ND | |
| 8 | B | ND | AX, AMC, CX, CRO, CL, FEP, S, DA, TE, E, SXT, AMP, NV | tet(M), erm(B) | ND |
| 9 | pauA | AX, CX, CRO, CL, FEP, S, DA, SXT, P, NV | blaZ, sul1 | qacC/D | |
| 10 | sua, pauA | AX, AMC, CX, CRO, CL, FEP, S, DA, TE, AMP, NV, K | tet(M) | ND | |
| 11 | skc | AX, CX, CRO, CL, FEP, S, DA, SXT, AMP, NV, C | sul1 | qacED1, qacC/D | |
| 12 | sua, pauA | CX, CRO, CL, FEP, S, DA, E, NV, NEO | aac(6′)aph(2″), erm(B) | ND | |
| 13 | sua, pauA | CX, CRO, CFP, CL, FEP, S, DA, E, AMP, NV, K, C | erm(B) | qacED1 | |
| 14 | sua, pauA | CX, CRO, CL, FEP, DA, E, AMP, NV, NEO, C | aac(6′)aph(2″), erm(B) | qacED1, qacC/D | |
| 15 | pauA | AX, CX, CRO, CL, FEP, S, DA, TE, E, SXT, AMP, NV | tet(M), erm(B), sul1 | qacC/D | |
| 16 | skc | AMC, CX, CRO, CL, FEP, S, DA, TE, SXT, P, AMP, NV, K | blaZ, tet(L), sul1 | ND | |
| 17 | C | ND | CX, CRO, CFP, CL, FEP, S, DA, TE, E, SXT, P, AMP, NV, NEO | blaZ, aac(6′)aph(2″), tet(M), erm(B), sul1 | qacA/B, qacED1, qacC/D |
| 18 | ND | AX, CX, CRO, CL, FEP, S, DA, E, SXT, P, AMP, NV, C | blaZ, fexA, erm(B), sul1 | qacED1, qacC/D | |
| 19 | sua | CX, CRO, CL, FEP, DA, E, P, AMP, NV, C | fexA, erm(B) | qacC/D | |
| 20 | sua | AX, CX, CRO, CL, FEP, S, DA, CIP, E, P, AMP, NV, K | erm(B) | qacED1, qacC/D | |
| 21 | sua | AX, AMC, CX, CRO, CL, FEP, S, DA, TE, E, SXT, P, AMP, NV, C | blaZ, fexA, tet(M), erm(B), sul1 | qacED1 | |
| 22 | sua | AX, CX, CRO, CL, FEP, S, DA, E, AMP, NV, NEO, C | aac(6′)aph(2″), erm(B) | qacA/B, qacED1, qacC/D | |
| 23 | sua | AX, CX, CRO, CL, FEP, S, DA, AMP, NV, C | ND | qacED1, qacC/D | |
| 24 | sua | AX, CX, CRO, CL, FEP, S, DA, TE, E, P, AMP, NV, C | fexA, tet(M), erm(B), sul1 | qacC/D | |
| 25 | sua | AX, CX, CRO, CFP, CL, FEP, S, DA, TE, E, SXT, NV, NEO, C | fexA, aac(6′)aph(2″), tet(M), erm(B), sul1 | qacED1, qacC/D | |
| 26 | sua, pauA, skc, cfu | CX, CRO, CL, FEP, S, DA, E, SXT, P, AMP, NV, NEO, K, C | blaZ, fexA, aac(6′)aph(2″), erm(B) | qacA/B, qacED1 | |
| 27 | sua, pauA, skc, cfu | AX, AMC, CX, CRO, CL, FEP, S, DA, TE, E, P, AMP, NV, C | blaZ, fexA, tet(M), erm(B), sul1 | qacC/D | |
| 28 | sua, pauA, skc, cfu | AX, CX, CRO, CFP, CL, FEP, S, DA, TE, E, SXT, P, AMP, NV, K, C | blaZ, fexA, tet(M), erm(B) | qacED1, qacC/D | |
| 29 | pauA | CX, CRO, CL, FEP, DA, E, P, AMP, NV, GEN, C | fexA, aac(6′)aph(2″), erm(B) | qacED1, qacC/D | |
| 30 | pauA | AX, CX, CRO, CL, FEP, S, DA, TE, AMP, NV, K, C | tet(M) | qacED1, qacC/D | |
| 31 | opp | AX, AMC, CX, CRO, CL, FEP, S, DA, TE, E, P, AMP, NV, C | tet(K), erm(B) | qacED1, qacC/D | |
| 32 | pauA, skc | AX, CX, CRO, CL, FEP, S, DA, TE, E, P, AMP, NV, K, C | fexA, tet(M), erm(B) | qacA/B, qacED1 | |
| 33 | pauA | AX, CX, CRO, CFP, CL, FEP, S, DA, TE, E, SXT, P, NV, GEN, NEO, K, C | fexA, aac(6′)aph(2″), tet(M), erm(B), sul1 | qacED1, qacC/D | |
| 34 | cfu | AMC, CX, CRO, CFP, CL, FEP, S, DA, P, AMP, NV, C | fexA | qacED1 | |
| 35 | opp | AX, CX, CRO, CL, FEP, S, DA, TE, SXT, P, AMP, NV, NEO, C | fexA, aac(6′)aph(2″), tet(M), sul1 | qacED1, qacC/D | |
| 36 | pauA, skc | AMC, CX, CRO, CFP, CL, FEP, S, DA, TE, E, P, AMP, NV, GEN | aac(6′)aph(2″), tet(L), erm(B) | qacED1, qacC/D | |
| 37 | sua | AX, CX, CRO, CFP, CL, FEP, S, DA, TE, E, P, AMP, NV | tet(M), erm(B) | qacA/B, qacED1, qacC/D | |
| 38 | pauA | AX, AMC, CX, CRO, CFP, CL, FEP, S, DA, TE, E, P, AMP, NV, NEO, C | aac(6′)aph(2″), tet(L), erm(B) | qacED1, qacC/D | |
| 39 | cfu | CX, CRO, CL, FEP, DA, P, AMP, NV | tet(M) | qacED1, qacC/D | |
| 40 | pauA, skc | AX, AMC, CX, CRO, CL, FEP, S, DA, CIP, TE, E, SXT, P, AMP, NV | tet(M), erm(B), sul1 | qacED1, qacC/D | |
| 41 | pauA | AX, CX, CRO, CL, FEP, S, DA, TE, P, AMP, NV, GEN, NEO | aac(6′)aph(2″), tet(L) | qacA/B, qacED1, qacC/D | |
| 42 | D | ND | AX, CX, CRO, CL, FEP, S, DA, TE, E, P, AMP, NV, NEO | aac(6′)aph(2″), tet(M), erm(B) | qacED1, qacC/D |
| 43 | ND | AX, CX, CRO, CL, FEP, DA, CIP, SXT, P, AMP, NV, GEN, NEO | aac(6′)aph(2″), sul1 | qacED1, qacC/D | |
| 44 | ND | AMC, CX, CRO, CFP, CL, FEP, S, DA, CIP, TE, E, SXT, P, AMP, NV, NEO | aac(6′)aph(2″), tet(M), erm(B), sul1 | qacED1, qacC/D | |
| 45 | ND | AX, AMC, CX, CRO, CFP, CL, FEP, S, DA, CIP, TE, E, SXT, P, AMP, NV, K | erm(B), sul1 | qacED1, qacC/D | |
| 46 | ND | AX, AMC, CX, CRO, CFP, CL, FEP, S, DA, TE, E, P, AMP, NV, GEN, K | aac(6′)aph(2″), erm(B) | qacED1, qacC/D | |
| 47 | ND | CX, CRO, CL, FEP, S, DA, CIP, TE, E, SXT, P, AMP, NV, K | blaZ, tet(M), erm(B), sul1 | qacED1, qacC/D | |
| 48 | ND | AX, CX, CRO, CL, FEP, S, DA, TE, E, SXT, P, AMP, NV, NEO | blaZ, aac(6′)aph(2″), tet(M), erm(B), sul1 | qacED1, qacC/D | |
| 49 | pauA | AX, CX, CRO, CL, FEP, DA, CIP, TE, E, SXT, P, AMP, NV | blaZ, erm(B), sul1 | qacA/B, qacED1, qacC/D | |
| 50 | sua, opp | AX, AMC, CX, CRO, CL, FEP, S, DA, TE, E, SXT, P, AMP, NV, GEN, NEO | blaZ, aac(6′)aph(2″), tet(L), erm(B), sul1 | qacED1, qacC/D | |
| 51 | cfu | AX, AMC, CX, CRO, CL, FEP, S, DA, CIP, E, P, AMP, NV, NEO | aac(6′)aph(2″), erm(B) | qacED1, qacC/D | |
| 52 | pauA | CX, CRO, CL, FEP, S, DA, TE, E, P, AMP, NV, NEO | aac(6′)aph(2″), tet(M), erm(B) | qacED1, qacC/D | |
| 53 | pauA | AX, CX, CRO, CL, FEP, S, DA, CIP, TE, SXT, P, AMP, NV, NEO | blaZ, aac(6′)aph(2″), tet(M), sul1 | qacED1, qacC/D | |
| 54 | cfu | AX, AMC, CX, CRO, CL, FEP, S, DA, CIP, TE, E, SXT, P, AMP, NV | blaZ, tet(M), erm(B), sul1 | qacED1, qacC/D | |
| 55 | sua, opp | AX, CX, CRO, CFP, CL, FEP, S, DA, SXT, P, AMP, NV, C | blaZ, sul1 | qacA/B, qacED1, qacC/D | |
| 56 | sua | AX, AMC, CX, CRO, CFP, CL, FEP, S, DA, CIP, TE, E, P, AMP, NV, GEN, C | blaZ, aac(6′)aph(2″), erm(B) | qacED1, qacC/D | |
| 57 | sua, pauA, skc, cfu | AX, AMC, CX, CRO, CFP, CL, FEP, S, DA, CIP, SXT, P, AMP, NV, GEN, NEO, C | blaZ, aac(6′)aph(2″), sul1 | qacED1, qacC/D | |
| 58 | sua, pauA, skc, cfu | AMC, CX, CRO, CFP, CL, FEP, S, DA, CIP, TE, E, P, AMP, NV, GEN, NEO, C | blaZ, aac(6′)aph(2″), tet(M), erm(B) | qacED1, qacC/D | |
| 59 | sua, pauA, skc, cfu | CX, CRO, CFP, CL, FEP, S, DA, TE, E, P, AMP, NV, C | blaZ, tet(M), erm(B) | qacED1, qacC/D | |
| 60 | sua, pauA, skc, cfu | CX, CRO, CL, FEP, DA, TE, E, P, AMP, NV, K, C | blaZ, fexA, tet(K), erm(B) | qacED1, qacC/D | |
| 61 | sua, pauA, skc, cfu | AX, CX, CRO, CL, FEP, S, DA, TE, E, SXT, P, AMP, NV, GEN, NEO, K, C | blaZ, fexA, aac(6′)aph(2″), tet(M), erm(B), sul1 | qacED1, qacC/D | |
| 62 | sua | AX, AMC, CX, CRO, CL, FEP, S, DA, TE, SXT, P, AMP, NV, NEO, K, C | blaZ, fexA, aac(6′)aph(2″), tet(K), sul1 | qacA/B, qacED1, qacC/D | |
| 63 | sua | CX, CRO, CFP, CL, FEP, S, DA, TE, E, SXT, P, AMP, NV, NEO, K, C | blaZ, fexA, aac(6′)aph(2″), tet(M), erm(B), sul1 | qacA/B, qacED1, qacC/D | |
| 64 | sua | AX, CX, CRO, CL, FEP, S, DA, E, P, AMP, NV, NEO, C | blaZ, erm(B) | qacA/B, qacED1, qacC/D | |
| 65 | sua | CX, CRO, CL, FEP, DA, E, P, AMP, NV, C | blaZ, aac(6′)aph(2″), erm(B) | qacED1, qacC/D | |
| 66 | sua | AX, CX, CRO, CL, FEP, S, DA, CIP, E, P, AMP, NV, C | blaZ, erm(B) | qacED1, qacC/D | |
| 67 | skc | AX, AMC, CX, CRO, CL, FEP, S, DA, TE, E, P, AMP, NV, GEN, NEO, K, C | blaZ, fexA, aac(6′)aph(2″), tet(M), erm(B) | qacA/B, qacED1, qacC/D | |
| 68 | skc | AX, CX, CRO, CL, FEP, S, DA, E, P, AMP, NV, GEN, NEO, K, C | blaZ, aac(6′)aph(2″), erm(B) | qacED1, qacC/D | |
| 69 | opp | AX, CX, CRO, CL, FEP, S, DA, P, AMP, NV, GEN, NEO, K, C | blaZ, fexA, aac(6′)aph(2″), | qacA/B, qacED1, qacC/D | |
| Molecular Pathotype | Virulence Genes | No. of S. uberis Isolates (%) | Farms |
|---|---|---|---|
| I | sua, pauA, skc, cfu | 8 (11.59) | C, D |
| II | pauA, cfu, opp | 3 (4.35) | A |
| III | sua, pauA | 4 (5.8) | B |
| IV | pauA, skc | 3 (4.35) | C |
| V | sua, opp | 2 (2.9) | D |
| VI | sua | 15 (21.74) | A, C, D |
| VII | pauA | 10 (14.49) | A, B, C, D |
| VIII | skc | 3 (4.35) | B, D |
| IX | cfu | 4 (5.8) | C, D |
| X | opp | 3 (4.35) | C, D |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd El-Aziz, N.K.; Ammar, A.M.; El Damaty, H.M.; Abd Elkader, R.A.; Saad, H.A.; El-Kazzaz, W.; Khalifa, E. Environmental Streptococcus uberis Associated with Clinical Mastitis in Dairy Cows: Virulence Traits, Antimicrobial and Biocide Resistance, and Epidemiological Typing. Animals 2021, 11, 1849. https://doi.org/10.3390/ani11071849
Abd El-Aziz NK, Ammar AM, El Damaty HM, Abd Elkader RA, Saad HA, El-Kazzaz W, Khalifa E. Environmental Streptococcus uberis Associated with Clinical Mastitis in Dairy Cows: Virulence Traits, Antimicrobial and Biocide Resistance, and Epidemiological Typing. Animals. 2021; 11(7):1849. https://doi.org/10.3390/ani11071849
Chicago/Turabian StyleAbd El-Aziz, Norhan K., Ahmed M. Ammar, Hend M. El Damaty, Rehab A. Abd Elkader, Hosam A. Saad, Waleed El-Kazzaz, and Eman Khalifa. 2021. "Environmental Streptococcus uberis Associated with Clinical Mastitis in Dairy Cows: Virulence Traits, Antimicrobial and Biocide Resistance, and Epidemiological Typing" Animals 11, no. 7: 1849. https://doi.org/10.3390/ani11071849
APA StyleAbd El-Aziz, N. K., Ammar, A. M., El Damaty, H. M., Abd Elkader, R. A., Saad, H. A., El-Kazzaz, W., & Khalifa, E. (2021). Environmental Streptococcus uberis Associated with Clinical Mastitis in Dairy Cows: Virulence Traits, Antimicrobial and Biocide Resistance, and Epidemiological Typing. Animals, 11(7), 1849. https://doi.org/10.3390/ani11071849