
Monitoring Campaign over an Edible Dormouse Population (Glis glis; Rodentia: Gliridae) in Sicily: First Report of Mesocestodiasis

 , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Parasitological Investigations
2.3. Investigation for Tularemia
2.4. Bacterial Isolation
2.5. Anatomopathological and Histological Examination
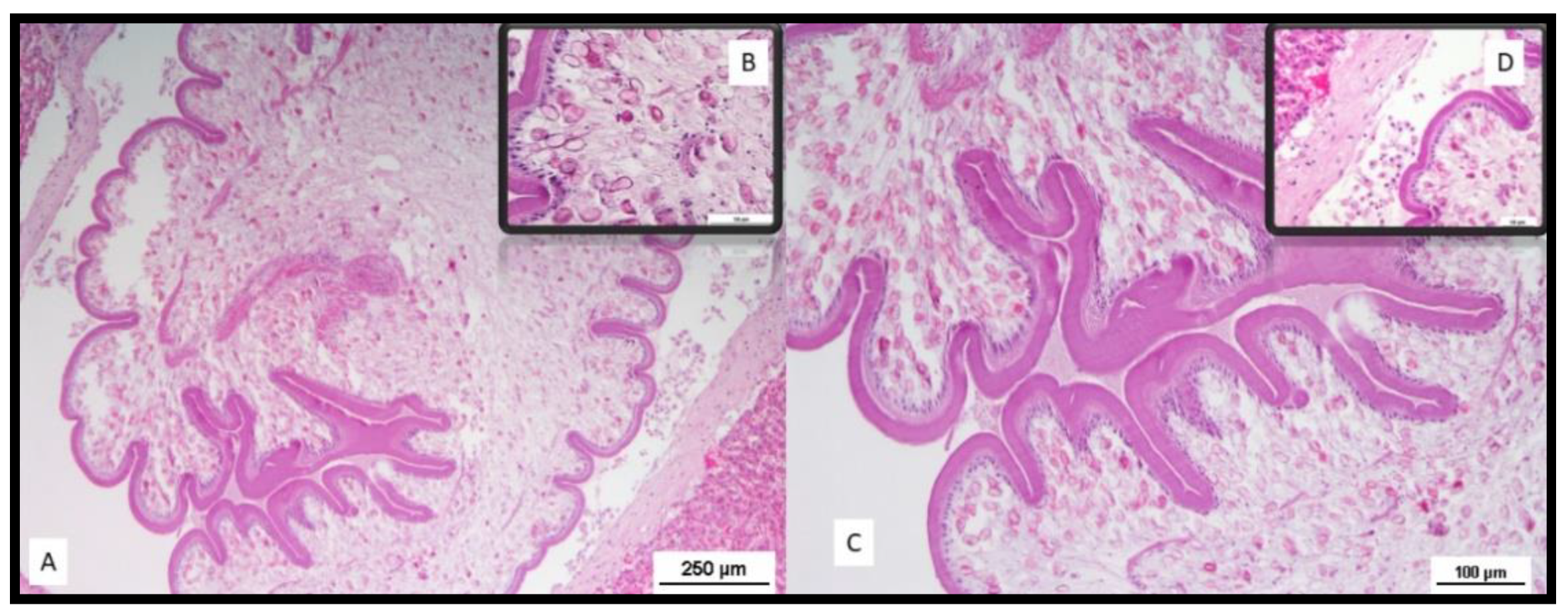
3. Results
- (1)
- Characteristics related to superfamilies and families:Posterior hip without small spiny bristles on the underside of the inner side;Wipe abdominals, each with more than one row of bristles (characteristic of the Ceratophylloidea superfamily);Of the two apical bristles of the anterior femur, the external one is longer;Methane with small marginal spines.
- (2)
- Characteristics of the genus:Comb of the pronotum consisting of less than 24 spines;Labial palp consisting of at least four segments and reaching the anterior trochanter as much as possible;Last item of all tarsi with five pairs of lateral plantar bristles;Head of the spermatheca as long as the tail (and a tail without papilla).
- (3)
- Characteristics of the spermatheca:Spermatheca’s head is piriform, and the copulating bag is non-spiral shaped: Monopsyllus sciurorum;Spermatheca’s head is globular and shows a spiral-shaped copulating bag: other species of Monopsyllus genus.
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Lipatova, I.; Stanko, M.; Paulauskas, A.; Spakovaite, S.; Gedminas, V. Fleas (Siphonaptera) in the Nests of Dormice (Gliridae: Rodentia) in Lithuania. J. Med. Entomol. 2015, 52, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Bieber, C. Population dynamics, sexual activity, and reproduction failure in the fat dormouse (Myoxus glis). J. Zool. 1998, 244. [Google Scholar] [CrossRef]
- Schlund, W.; Scharfe, F.; Ganzhorn, J.U. Long-term comparison of food availability and reproduction in the edible dormouse (Glis glis). Mamm. Biol. 2002, 67, 219–232. [Google Scholar] [CrossRef]
- Pilastro, A.; Tavecchia, G.; Marin, G. Long living and reproduction skipping in the fat dormouse. Ecology 2003, 84, 1784–1792. [Google Scholar] [CrossRef]
- Lebl, K.; Bieber, C.; Adamík, P.; Fietz, J.; Morris, P.; Pilastro, A.; Ruf, T. Survival rates in a small hibernator, the edible dormouse: A comparison across Europe. Ecography 2011, 34, 683–692. [Google Scholar] [CrossRef]
- Ruf, T.; Fietz, J.; Schlund, W.; Bieber, C. High survival in poor years: Life history tactics adapted to mast seeding in the edible dormouse. Ecology 2006, 87, 372–381. [Google Scholar] [CrossRef]
- Bieber, C.; Ruf, T. Habitat differences affect life history tactics of a pulsed resource consumer, the edible dormouse (Glis glis). Popul. Ecol. 2009, 51, 481–492. [Google Scholar] [CrossRef]
- Schlund, W.; Scharfe, F.; Stauss, M.J.; Burkhardt, J.F. Habitat fidelity and habitat utilization of an arboreal mammal (Myoxus glis) in two different forests. Z. Saugetierkd. 1997, 62, 158–171. [Google Scholar]
- Morris, P.A.; Hoodless, A. Movements and hibernaculum site in the fat dormouse (Glis glis). J. Zool. 1992, 228, 685–687. [Google Scholar] [CrossRef]
- Stanojevic, M.; Nikolic, V.; Stajkovic, N.; Stamenkovic, G.; Bozovic, B.; Cekanac, R.; Marusic, P.; Gligic, A. Genetic detection of Dobrava-Belgrade hantavirus in the edible dormouse (Glis glis) in central Serbia. Epidemiol. Infect. 2015, 143, 400–404. [Google Scholar] [CrossRef]
- Earl, P.L.; Americo, J.L.; Cotter, C.A.; Moss, B. Comparative live bioluminescence imaging of monkeypox virus dissemination in a wild-derived inbred mouse (Mus musculus castaneus) and outbred African dormouse (Graphiurus kelleni). Virology 2015, 475, 150–158. [Google Scholar] [CrossRef]
- Ehlers, B.; Anoh, A.E.; Ben Salem, N.; Broll, S.; Couacy-Hymann, E.; Fischer, D.; Gedvilaite, A.; Ingenhütt, N.; Liebmann, S.; Martin, M.; et al. Novel Polyomaviruses in Mammals from Multiple Orders and Reassessment of Polyomavirus Evolution and Taxonomy. Viruses 2019, 11, 930. [Google Scholar] [CrossRef] [PubMed]
- Matuschka, F.R.; Eiffert, H.; Ohlenbusch, A.; Spielman, A. Amplifying role of edible dormice in Lyme disease transmission in central Europe. J. Infec. Dis. 1994, 170, 122–127. [Google Scholar] [CrossRef]
- Fietz, J.; Langer, F.; Havenstein, N.; Matuschka, F.R.; Richter, D. The vector tick Ixodes ricinus feeding on an arboreal rodent-the edible dormouse Glis glis. Parasitol. Res. 2016, 115, 1435–1442. [Google Scholar] [CrossRef]
- Matuschka, F.R.; Allgöwer, R.; Spielman, A.; Richter, D. Characteristics of garden dormice that contribute to their capacity as reservoirs for lyme disease spirochetes. Appl. Environ. Microbiol. 1999, 65, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Faber, M.; Heuner, K.; Jacob, D.; Grunow, R. Tularemia in Germany-A Re-emerging Zoonosis. Front. Cell Infect Microbiol. 2018, 8, 40. [Google Scholar] [CrossRef] [PubMed]
- Bártová, E.; Kučerová, H.L.; Žákovská, A.; Budíková, M.; Nejezchlebová, H. Coxiella burnetii and Francisella tularensis in wild small mammals from the Czech Republic. Ticks Tick Borne Dis. 2020, 11, 101350. [Google Scholar] [CrossRef]
- Berlinguer, G. Aphaniptera d’Italia: Studio Monografico, 2nd ed.; Pensiero Scientifico: Rome, Italy, 1964. [Google Scholar]
- Bowles, J.; McManus, D.P. Genetic characterization of the asian Taenia, a newly described taeniid cestode of humans. Am. J. Trop. Med. Hyg. 1994, 50, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Gasser, R.B.; Zhu, X.; McManus, D.P. NADH dehydrogenase subunit 1 and cytochrome c oxidase subunit I sequences compared for members of the genus Taenia (Cestoda). Int. J. Parasitol. 1999, 29, 1965–1970. [Google Scholar] [CrossRef]
- Versage, J.L.; Severin, D.D.M.; Chu, M.C.; Petersen, J.M. Development of a Multitarget Real-Time TaqMan PCR Assay for Enhanced Detection of Francisella tularensis in Complex Specimens. J. Clin. Microbiol. 2003, 41, 5492–5499. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.R. Diagnostic Procedure in Veterinary Bacteriology and Mycology, 5th ed.; Carter, G., Cole, J., Jr., Eds.; Academic Press: San Diego, CA, USA, 1990. [Google Scholar]
- Makarikov, A.A. A taxonomic review of hymenolepidids (Eucestoda, Hymenolepididae) from dormice (Rodentia, Gliridae), with descriptions of two new species. Acta Parasitol. 2017, 62, 1–21. [Google Scholar] [CrossRef]
- Mas-Coma, S.; Gallego, J. Pseudophysaloptera kahmanni n. sp. (Nematoda: Physalopteridae) parasite de l’estomac du Lérot Eliomys quercinus ophiusae Thomas, 1925 (Rodentia: Gliridae) à Formentera (Baléares). Ann. Parasitol. Hum. Comp. 1977, 52, 19–24. [Google Scholar] [CrossRef]
- Mas-Coma, S. Gongylonema pithyusensis n. sp. (Nematoda: Spiruridae), parasite œsophagien du Lérot Eliomys quercinus ophiusae Thomas, 1925 (Rodentia: Gliridae) à Formentera (Baléares). Ann. Parasitol. Hum. Comp. 1977, 52, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, M.; Sainsbury, A.W.; Sayers, G. Nematode infestation with Heligmosomoides polygyrus in captive common dormice (Muscardinus avellanarius). Vet. Rec. 2006, 158. [Google Scholar] [CrossRef]
- Friedl, A.; Heinzer, I.; Fankhauser, H. Tularemia after a dormouse bite in Switzerland. Eur. J. Clin. Microbiol. Infect. Dis. 2005, 24, 352–354. [Google Scholar] [CrossRef]
- Medved, S.G. Geographical distribution of families of fleas (Siphonaptera). Entomol. Rev. 1996, 76, 978–992. [Google Scholar]
- Morrone, J.J.; Acosta, R. A synopsis of the fleas (Insecta: Siphonaptera) parasitizing New World species of Soricidae (Mammalia: Insectivora). Zootaxa 2006, 1354, 1–30. [Google Scholar]
- Peus, F. Flöhe aus dem Mittelmeergebiet (Insecta, Siphonaptera) III. Sizilien. Mitt. Mus. Nat. Berl. Zool. Mus. Inst. Spez. Zool. 1959, 35, 79–106. [Google Scholar] [CrossRef]
- Krampitz, H.E. Present-day Sicily from a hygienic-zoological point of view. Z. Trop. Parasitol. 1958, 9, 111–128. [Google Scholar]
- Hawlena, H.; Khokhlova, I.S.; Abramsky, Z.; Krasnov, B.R. Age, intensity of infestation by flea parasites and body mass loss in a rodent host. Parasitology 2006, 133, 187–193. [Google Scholar] [CrossRef]
- Kammeyer, P.; Hartmann, D.; Busch, A.; Tomaso, H.; Braune, S.; Runge, M.; Kleinschmidt, S. Glis glis–two cases of acute tularemia. Berl. Münch. Tierärztl. Wochenschr. 2019, 132, 454–458. [Google Scholar] [CrossRef]
- Gambino, D.; Vicari, D.; Vitale, M.; Schirò, G.; Mira, F.; Giglia, M.; Riccardi, A.; Gentile, A.; Giardina, S.; Carrozzo, A.; et al. Study on Bacteria Isolates and Antimicrobial Resistance in Wildlife in Sicily, Southern Italy. Microorganisms 2021, 9, 203. [Google Scholar] [CrossRef]
- Sarvi, S.; Daryani, A.; Sharif, M.; Rahimi, M.T.; Kohansal, M.H.; Mirshafiee, S.; Siyadatpanah, A.; Hosseini, S.-A.; Gholami, S. Zoonotic intestinal parasites of carnivores: A systematic review in Iran. Vet. World 2018, 11, 58–65. [Google Scholar] [CrossRef]
- Fuentes, M.V.; Galán-Puchades, M.T.; Malone, J.B. Short report: A new case report of human mesocestoides infection in the united states. Am. J. Trop. Med. Hyg. 2003, 68, 566–567. [Google Scholar] [CrossRef] [PubMed]
- Eom, K.S.; Kim, S.H.; Rim, H.J. Second case of human infection with Mesocestoides lineatus in Korea. Korean J. Parasitol. 1992, 30, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.G.; Yi, S.H.; Liu, Z. The first case of human infections with Mesocestoideslineatus (Goeze, 1782) in Jilin Province. J. Norman Bethune Univ. Med. Sci. 1991, 4, 360–361. [Google Scholar]
- Schultz, L.J.; Roberto, R.R.; Rutherford, G.W.; Hummert, B.; Lubell, I. Mesocestoides (cestoda) infection in a california child. Pediatr. Infect. Dis. J. 1992, 11, 332–334. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schirò, G.; Pieri, D.; Lo Valvo, M.; Gradoni, L.; Cacciò, S.M.; Severini, F.; Marucci, G.; Galuppo, L.; Cumbo, V.; Puleio, R.; et al. Monitoring Campaign over an Edible Dormouse Population (Glis glis; Rodentia: Gliridae) in Sicily: First Report of Mesocestodiasis. Animals 2021, 11, 934. https://doi.org/10.3390/ani11040934
Schirò G, Pieri D, Lo Valvo M, Gradoni L, Cacciò SM, Severini F, Marucci G, Galuppo L, Cumbo V, Puleio R, et al. Monitoring Campaign over an Edible Dormouse Population (Glis glis; Rodentia: Gliridae) in Sicily: First Report of Mesocestodiasis. Animals. 2021; 11(4):934. https://doi.org/10.3390/ani11040934
Chicago/Turabian StyleSchirò, Giorgia, Domenico Pieri, Mario Lo Valvo, Luigi Gradoni, Simone Mario Cacciò, Francesco Severini, Gianluca Marucci, Lucia Galuppo, Valentina Cumbo, Roberto Puleio, and et al. 2021. "Monitoring Campaign over an Edible Dormouse Population (Glis glis; Rodentia: Gliridae) in Sicily: First Report of Mesocestodiasis" Animals 11, no. 4: 934. https://doi.org/10.3390/ani11040934
APA StyleSchirò, G., Pieri, D., Lo Valvo, M., Gradoni, L., Cacciò, S. M., Severini, F., Marucci, G., Galuppo, L., Cumbo, V., Puleio, R., & Loria, G. R. (2021). Monitoring Campaign over an Edible Dormouse Population (Glis glis; Rodentia: Gliridae) in Sicily: First Report of Mesocestodiasis. Animals, 11(4), 934. https://doi.org/10.3390/ani11040934