1. Introduction
Research since the late 1990s has substantially broadened our understanding of the cognitive abilities of the domestic dog—in particular, how dogs perceive the world and interact with others. The greatest majority of research focuses on social cognitive abilities, such as gaze- and point-following [
1,
2] and use of others’ attention [
3,
4]; other research looks at the possible result of such social cognitive skill, such as perspective-taking [
5,
6] and representation of self [
7]. Less well studied are the contributions of foundational perceptual abilities to cognition.
Recent work has begun to investigate the cognitive consequences of olfactory perception [
8,
9]; very few research programs have examined perception of affordances and body-size awareness [
10,
11,
12]. Animals’ information about their body size, especially relative to environmental changes and constraints—body sense—is critical to navigating new environments as well as social success. An animal’s size guides their behavior and, for many species, can be a key determinant in both their short- and long-term survival [
13,
14]. Relative body size also relates to reproductive output [
15,
16] and fighting behavior [
17]. A size sense is also integral to navigating changing environments: it has been proposed that the capacity for self-concept evolved to enable large animals to navigate complex environments [
18]. While most animals’ environments fluctuate with naturalistic changes, such as a fallen tree blocking a habitual route, the environments of species who live among humans are additionally driven by human behavior. For an owned dog living in a human household, various aspects of their environment may be altered daily: a chair pushed away from a table or a door closed partway. In these scenarios, an animal’s sense of their own size would facilitate negotiation of their changing surroundings [
19,
20].
Previous research with dogs has investigated their perception of size primarily of conspecifics, not themselves. In a looking-time paradigm, Taylor et al. [
21] found that dogs could match growls of different pitches with corresponding images of dogs of different sizes. Other work has found that dogs accurately matched food-guarding growls with an image of the source of the growl, but looked toward the image of a dog larger than the source when hearing playful growls [
22]. Some research addresses a dog’s understanding of their body size with respect to the size of other dogs. Dogs self-handicap with smaller dogs in play [
23] and small dogs perform higher raised-leg displays relative to their body size than large dogs do [
24].
Having an understanding of one’s size relative to features in the environment also enables determination of what actions are possible in a given scenario, or the “affordances” of the environment [
20,
25]. Human studies show that perceptual input—primarily visual—is used to guide decisions about what actions can be performed [
20], sensitive to changes of one’s own body and of one’s environment [
26]. Subjects attempt to reach or move through openings larger, but not smaller, than their appendage or body size, and make appropriate adjustments to move through narrow openings [
27,
28]. Moreover, subjects wearing size-increasing prosthetics (such as of the hand or stomach) made appropriate adjustments, with experience [
27,
29]. In research with canine subjects, Wagman et al. [
12] evaluated dog behavior when attempting to acquire a treat placed at increasingly higher points along a wall; they found that the height at which subjects transition from reaching for a treat with their head only to rearing was different across individuals but with the same ratio of shoulder height to treat height [
30]. Two projects looked at dogs’ behavior when approaching differently sized openings. Maeda and Fujita [
11] found that when simultaneously given a larger or smaller doorway opening, subjects preferred to go through the larger opening. Lenkei et al. [
10] found longer latencies for subjects to approach too narrow or short openings than openings through which they could fit; moreover, subjects did not attempt to pass through openings that were too small [
10]. These studies provide preliminary evidence that dogs can perceive affordances of a given environment and integrate information about the external world and their own body to organize their behavior.
The present research extends this literature into dogs’ knowledge of their body size, and further explores their understanding of the affordances of the environment. In the first study, we assess whether dogs understand their size relative to an opening which they are asked to pass through by examining behavioral modifications and approach time as the opening becomes increasingly shorter. As in Lenkei et al. [
10], we assess subjects’ judgment of their ability to fit through the openings by measuring their latency to approach openings of decreasing height. We predict that if dogs have a knowledge of their own body size, then they will accurately judge which openings are passable, showing longer latencies to approach smaller openings. In addition, we analyze dogs’ physical adjustments as they attempt to pass through increasingly smaller openings, and examine how their behavior relates to their height at withers [
12], eye-height [
31], or elbow joint-height (which may differ by breed [
32]). As the opening is adjusted incrementally along one dimension, we can determine the ratio at which dogs first adjust their behavior to negotiate the opening, relative to subjects’ height at withers. We predict that the order of physical adjustments will be invariant across dogs, and thus consistent across different body sizes, as will the ratio of body-height to the height of the opening at which dogs make their first bodily adjustment.
Given the importance of tactile information, acquired via experience, to subject behavior [
29], in a second study, we examine subject behavior when their body size is effectively increased. To naturalistically and temporarily increase their width, we offer dogs sticks of various lengths to hold in their mouths while passing through a fixed-width opening. We measure subject modifications in approach time and body position with variously sized sticks, and offer subjects the chance to choose a stick that will easily fit through the aperture. We predict subjects will inaccurately judge affordances as stick length increases—leading to an inability to fit through the opening—but will be able to update their perception of possible actions with experience, and alter their approach so to fit.
3. Results
3.1. Study 1
Forty-three dogs participated in 424 trials (
n = 1 dog refused to participate in any of the trials). Subjects were categorized into five size bins on the basis of height at withers: XS (
n = 5), S (
n = 9), M (
n = 13), L (
n = 16), XL (
n = 1) (see
Table 2 for height ranges per category, and
Table 1 for individual participant characteristics). For analysis, the single XL dog was included in the L bin category.
Subjects attempted to pass through the aperture an average of 9.65 times (SD = 2.14), and successfully passed through the aperture an average of 7.57 times (SD = 1.98). A linear regression of subjects’ age and the number of successful trials completed showed no significant correlation between age and performance (
r2 = 0.01,
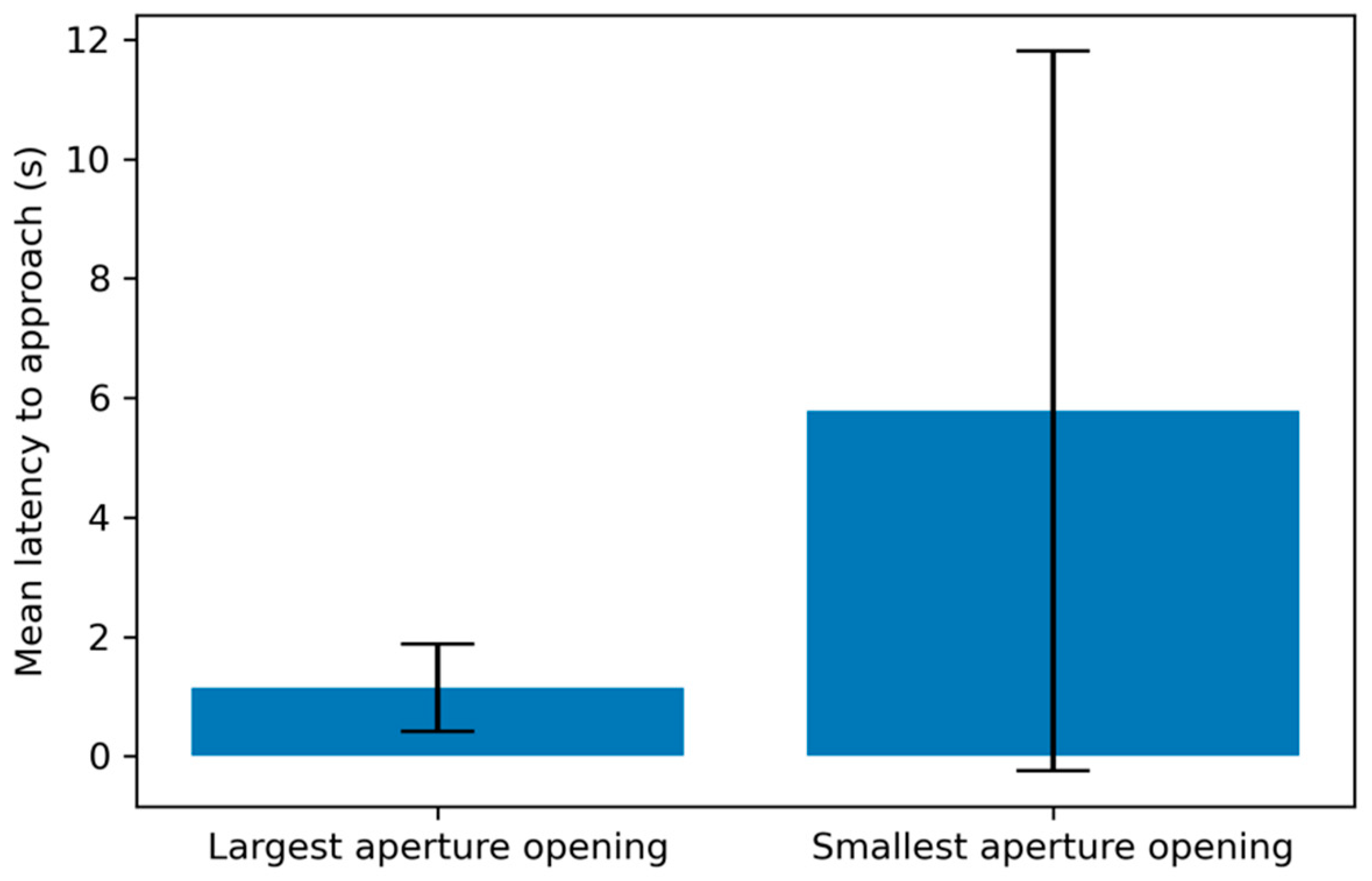
p > 0.05). The subjects modified their behavior on an average of 6.09 trials (SD = 1.76), or an average of 61.8% of all the trials they completed (SD = 15.6%). Subject latency to reach the aperture depended on its height: latency was significantly longer on subjects’ final successful trial (M = 5.78 s, SD = 6.02) than on the first trial (M = 1.15 s, SD = 0.73) t(42) = −4.95,
p < 0.001) (
Figure 4). There was no significant difference in latency between the first trial and the first trial with a behavioral modification (t(42) = −1.40,
p = 0.16).
Across subjects, the final aperture height negotiated varied according to the subjects’ size. A linear regression of subjects’ height at withers and the aperture height on the final successful trial—the trial on which the subject passed through the aperture—showed a positive correlation (
r2 = 0.52,
p < 0.001;
Figure 5). Eye height (
r2 = 0.44,
p < 0.001) and elbow height (
r2 = 0.50,
p < 0.001) were also positively correlated, less strongly, with final aperture height.
The final aperture height successfully negotiated was significantly higher for dogs in the tall group (
M = 62.22 cm, SD = 4.90) than for dogs in the short group (
M = 37.58 cm, SD = 10.54),
t(41) = −8.57,
p < 0.001;
Figure 6). Considered by their bin category, the final aperture height successfully negotiated was significantly higher for dogs in the large group (M = 35.66 cm, SD = 11.70) than for dogs in the medium group (M = 20.5 cm, SD = 3.15), t(41) = −4.40,
p < 0.001), the small group (M = 19.0 cm, SD = 7.42), t(41) = −3.70,
p = 0.001), and the extra-small group (M = 14.4 cm, SD = 3.26), t(41) = −3.82,
p = 0.001). Furthermore, the final aperture height was significantly higher for dogs in the medium group than dogs in the extra-small group (t(41) = −3.41,
p = 0.003).
To determine if the height at which dogs made their first adjustment was the same across dogs of different size, we calculated the mean aperture height for the first adjustment, divided by dog’s HW, for both short and tall dogs. Comparison of these ratios found no significant difference for dog sizes (Mshort = 0.91; Mtall = 0.91; t(41) = −0.07, p > 0.05).
To examine whether dogs used a common strategy to negotiate the aperture, Chi-squared goodness of fit tests were computed on frequencies of the use of specific behavioral adjustments to fit through the aperture—as well as the frequencies of overall adjustment sequences across trials. Subjects’ first behavioral adjustment was significantly more likely to be a head duck than any of the other behaviors (X2(3, n = 43) = 97.05, p < 0.001). The second adjustment was often either a front-elbow bend or a back-elbow bend (X2(3, n = 43) = 34.02, p < 0.001), with no significant difference between the two (X2(1, n = 39) = 0.02, p = 0.63). Similarly, the third behavior was most often either a front elbow bend or back elbow bend (X2(3, n = 43) = 28.56, p < 0.001), with no significant difference in frequency between the two (X2(1, n = 37) = 0.68, p = 0.41). The fourth behavioral adjustment was significantly more likely to be the body turn (X2(3, n = 43) = 39.78, p < 0.001) than any other behavior.
Moreover, adjustment sequence “head duck–front elbow–back elbow–body turn” and “head duck–back elbow–front elbow–body turn” were significantly more likely to occur than any other order of behavioral adjustments (
X2(3,
n = 41) = 23.10,
p < 0.001) (
Figure 7). There was no significant difference between the frequency of these two sequences. An independent Chi-square test revealed no significant relationship between the adjustment sequence chosen and subject body size (
X2(16,
n = 41) = 7.94,
p = 0.54).
A linear regression of subjects’ height at withers on the aperture height when subjects made specific behavioral adjustments revealed a strong positive correlation with height at head duck, with height at withers accounting for 75% of variance in the height of aperture (r
2 = 0.75,
p < 0.001) (
Figure 8). A significant correlation is also seen between height at withers and aperture height at which subjects first bent their elbows (either front or back) (r
2 = 0.87,
p < 0.001) (
Figure 9).
Three dogs used the alternate route on one trial: two on their final height, after failing to go through the aperture, and one who used it but later returned to navigating the opening on the next trial.
3.2. Study 2
Thirteen dogs were selected to participate in Study 2; three dogs did not successfully complete the training trials, and four dogs did not successfully complete the first experimental trial. Thus, data were analyzed from n = 6 dogs. Four of the six dogs fully completed the procedure: one did not make it through the aperture successfully on all trials.
Due to the high attrition rate, inferential statistical analysis was not conducted; all analyses are descriptive. All six subjects successfully passed through the aperture with the small and large stick; all but one successfully passed through with the medium stick. Subjects attempted to pass through the aperture an average of 1.5 times with the small stick, 1.17 times with the medium stick, and 1.67 times with the large stick.
Latency to reach the aperture with the small stick was 21.79 s (SD = 27.58), excluding one subject outlier who had an approach time of 1020 s. Latency to approach with the medium stick was 1.54 s (SD = 0.76); with the large stick, 4.06 s (SD = 5.21).
Behavioral adjustments on these trials included head turns (small stick: 1; medium stick: 1; large stick, 2) and dropping the stick (mean for small stick: 1; medium stick: 0.83; large stick, 1.16). Subjects more often hit the side of the aperture with the large stick (mean = 1.33) than with the small or medium sized sticks (mean = 0.5). When all six subjects were given a final choice of stick on the final trial, three chose the large stick, one chose the small, one chose the medium, and one made no choice.
4. Discussion
These studies were designed to address subject dogs’ behavior when asked to go through an opening (aperture) of decreasing height, until they no longer fit through the opening. The results speak to dogs’ perception of their body size with respect to their environment. This suggests that dogs not only perceive others’ body sizes, as previously found [
21,
22,
24], but also their own. The dogs in this study used knowledge of their body size when judging whether they would fit through an opening: the latency for subjects to reach an aperture they could easily fit through was significantly shorter than to one which was almost too small to fit through. This finding is consistent with Lenkei et al. [
10], who also found a longer latency to approach a too-small opening. Both our results suggest that dogs distinguish between easily passable and difficult to pass openings before acting.
As expected, the final height of the aperture differed significantly between short and tall dogs and across size bins (XS, S, M, L) and correlated with several measurements: the dog’s height at withers, elbows, and eyes. This indicates that what is considered “too small” to fit through is dependent on the dog’s sense of their own size. Moreover, as all measurements of the dog’s body appear to influence their perception of affordances, it is possible that the dog uses a crude representation of their body to determine what actions are possible. Notably, at the same time, subjects’ increase in latency across trials cannot be explained by, for instance, a decrease in motivation as the aperture diminished in height, as most subjects continued to attempt to squeeze through the opening even when it was impossible for them to pass through it.
Interestingly, in the present study, 12 subjects passed through an aperture shorter than one-third of their height at withers; this is smaller in height than what was considered “too small” in Lenkei et al. [
10]. Many more subjects unsuccessfully attempted to pass through an opening that was in fact too small for them to fit through even with bodily modifications. This difference between subject behavior in the present study and in Lenkei et al. [
10] might be explained as resulting from the latter’s adjustment of both the height and width of the aperture (in their Experiment 2), while the present study only adjusted the height. An aperture which is not wide enough to pass through may be more obviously “too small”, as subjects cannot adjust their behavior to significantly reduce their width. By contrast, dogs can adjust their height by, for example, ducking their head or bending their elbows. In addition, decreasing the aperture size incrementally may have enabled more precision in determination of what size was too small for the subjects.
Additionally, the subjects’ different behavior may be related to the cost or benefit associated with negotiating the experimental apparatus in the two studies. Studies with human infants have found that subjects decide whether to use a small doorway according to the related risk: many infants err and get stuck when squeezing into too small openings, but they do not attempt a too small aperture when such an error would result in falling [
33]. This indicates that infants can perceive the affordances allotted by an aperture, but also use information about the cost of using the aperture when planning their actions. Dogs may be similarly evaluating the associated risk during their decision-making process: there was very little risk in passing through the similar apparatuses used in Lenkei et al. [
10] and in this study. However, in addition, in the present study, there was an added benefit: their owners held a treat on the opposite side of the apparatus and were reachable only by successfully negotiating the apparatus. While Lenkei et al. [
10] suggested that dogs’ motivation to get through their opening was high even without the owners (Experiment 1) or without a treat (Experiment 2 and 3), the present study points to the added motivation present when both owner and treat are reached by completing the task.
To further understand the dog’s decision-making process, we sought to determine not only when a dog decides whether they can or cannot fit through an aperture, but also
how dogs fit through an aperture. Thus, we extended our analyses to the order of behavioral adjustments both across and within trials. Through examination of the sequence of bodily modifications made by subjects as they negotiated ever-smaller openings, we can suggest that subjects’ perception of affordances to fit through an aperture is action-scaled. Our results revealed that subjects’ first adjustment came at the same ratio of dogs’ height to aperture height across sizes. Moreover, we found a preferred sequence to adjustments: subjects most often first ducked their heads, then bent front or back elbows, then turned their body. This sequence was relatively invariant across individuals, regardless of body size, providing evidence that dogs know which actions are necessary in the face of environmental constraints. Across subjects, there was a positive correlation between the size of the aperture opening and the size of the subjects’ withers, eye-height, and elbow height. Dog behavior reflects an appreciation of the task—navigating the aperture—relative to the capabilities and constraints of their own bodies. The timing of specific bodily adjustments correlated with subject size: dogs’ height at withers predicts when subjects would adjust their bodies by ducking their head or bending their elbow to fit through ever-diminishing apertures. Similarly, Wagman et al. [
12] found that dogs’ perception of the affordances for reaching was action-scaled, with the ratio of the stimulus height to body height at which subjects moved from reaching to rearing the same for tall dogs and small dogs. The present work adds to the small body of literature examining non-human animals’ navigation of novel environments, which has found, for instance, that snakes change behavioral strategies depending on the affordances of their environment [
34], and frogs prefer jumping through horizontal openings than same-size vertical openings, in line with their physical profile [
35].
Our results in Study 2 must be considered preliminary because of the small sample size. Indeed, subject data provide no clear message. Subject latencies to reach the aperture were longer with a stick that fit than with a stick that did not fit, against expectations. Subjects made multiple attempts to pass through the aperture with all stick sizes. In line with our prediction that subjects may inaccurately judge affordances when their bodies were effectively vertically extended, the sticks often hit the side of the aperture—more with the large-sized stick than the small- or medium-sized sticks. Certainly, subject behavior was not consistent with the idea that they had foreknowledge of the way the stick had changed their size. Similarly, it has been reported that dogs often bump into objects when navigating a familiar environment with an unfamiliar neck appendage like the Elizabethan collar, a flexible cone usually of plastic that is secured around a dog’s neck after surgeries in order to prevent the dog’s oral manipulation of a surgical site. This is suggestive that dogs’ perception of their size does not extend to appendages adding to their height or width.
Instead, modifications to their body (by turning their head) were performed only after knocking the apparatus: possible evidence that subjects updated their perception of affordances with experience. Future research examining these questions may want to include training for dogs to reliably hold or carry sticks in order to assure a robust sample size.
These results indicate that, much like humans [
26,
28], dogs integrate complex information about their body size with knowledge of how adjustment behaviors can alter their size when determining which actions are possible. Future research may explore the role of specific experiences in developing this “size sense”. Humans require experience with their artificially altered body size to update their perception of affordances [
29], and preliminary evidence from Experiment 2 suggests that dogs are, at least initially, similarly inaccurate at determining affordances when their width is effectively increased.
This work suggests that dogs’ representation of their body size extends beyond the olfactory modality [
8] and past actions [
7]. Prior research on self-representation, using the mirror-mark test, has found mixed results with non-humans, some of which may be due to problems resulting from adapting primate-centric cognitive paradigms to non-primates [
8]. Insofar as a sense of one’s size may be described as a representation of oneself [
10,
18], testing an animal’s sense of their size in a changing environment may be a novel methodological way to investigate the notoriously elusive sense of self.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}