
Multiparametric Study of Antioxidant Effect on Ram Sperm Cryopreservation—From Field Trials to Research Bench

 , ,
, ,  , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sample Collection
2.3. Experimental Design and Antioxidant Supplementation
2.4. Freezing and Thawing Protocols
2.5. Fertility Trial: Field Inseminations
2.6. In Vitro Assay: Sperm Quality Analyses
2.6.1. Sperm Motility and Kinetic Parameters
2.6.2. Multiparametric Flow Cytometry Analyses
2.6.3. RedoxSYS Analysis
2.7. Statistical Analysis
3. Results
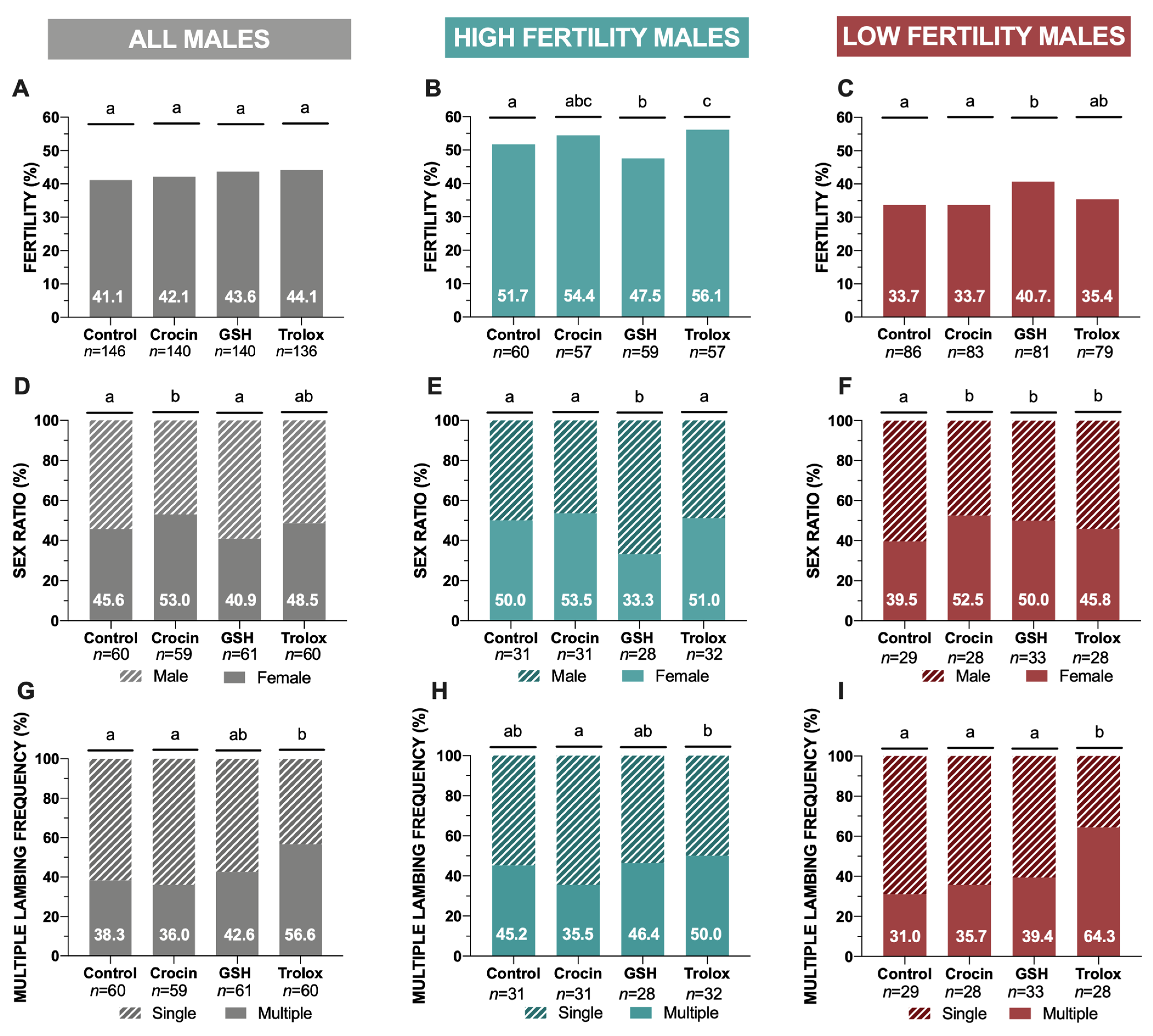
3.1. In Vivo Results: Fertility Trials
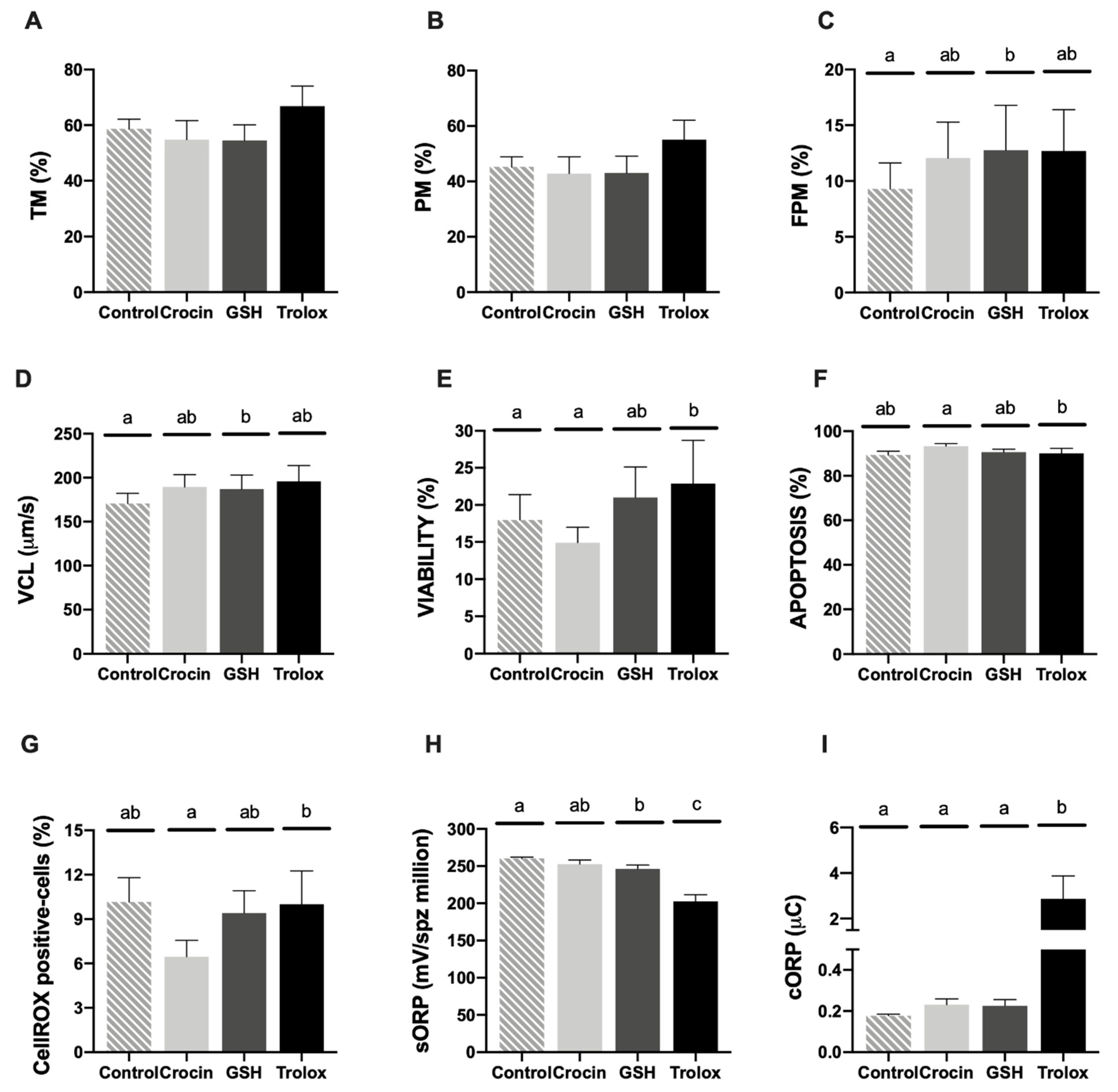
3.2. In Vitro Results: Sperm Quality Assays
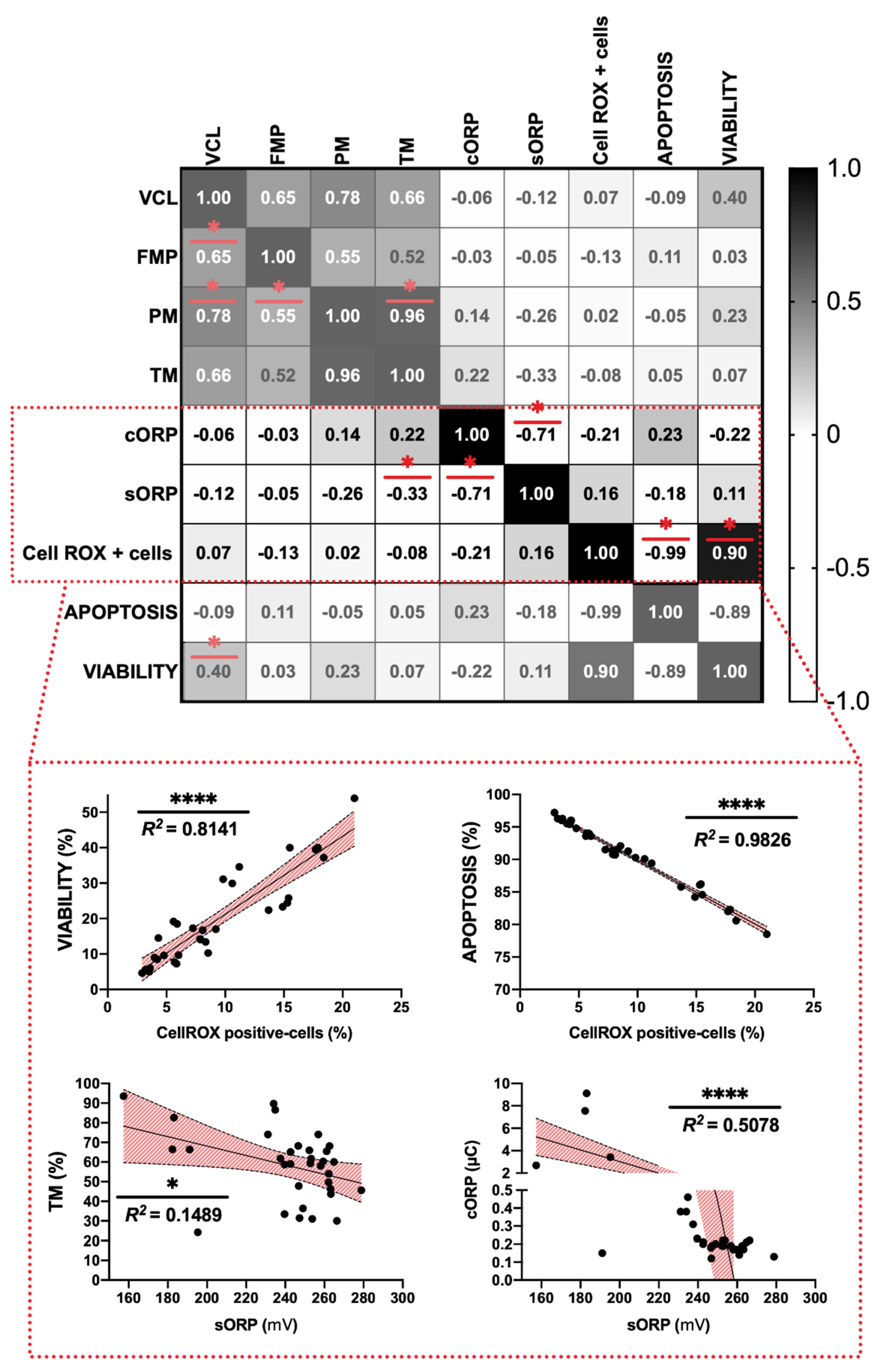
3.3. Correlation of Redox Balance Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Global | Prolificacy | Fecundity | Litter Size Distribution | |||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | ||||
| Control | 1.42 ± 0.07 | 0.58 ± 0.07 | 61.8 | 35.0 | 3.3 | |
| Crocin | 1.41 ± 0.08 | 0.59 ± 0.07 | 64.4 | 30.5 | 5.1 | |
| GSH | 1.44 ± 0.07 | 0.63 ± 0.07 | 57.4 | 41.0 | 1.6 | |
| Trolox | 1.62 ± 0.08 | 0.71 ± 0.08 | 43.3 | 51.7 | 5.0 | |
| HFM | Prolificacy | Fecundity | Litter Size Distribution | |||
| 1 | 2 | 3 | ||||
| Control | 1.52 ± 0.11 | 0.78 ± 0.11 | 69.0 | 31.0 | 0 | |
| Crocin | 1.39 ± 0.10 | 0.75 ± 0.11 | 64.3 | 28.6 | 7.1 | |
| GSH | 1.50 ± 0.11 | 0.71 ± 0.11 | 60.6 | 39.4 | 0 | |
| Trolox | 1.53 ± 0.10 | 0.86 ± 0.12 | 35.7 | 57.1 | 7.1 | |
| LFM | Prolificacy | Fecundity | Litter Size Distribution | |||
| 1 | 2 | 3 | ||||
| Control | 1.31 ± 0.09 | 0.44 ± 0.07 | 54.8 | 38.7 | 6.5 | |
| Crocin | 1.43 ± 0.12 | 0.48 ± 0.08 | 64.5 | 32.3 | 3.2 | |
| GSH | 1.39 ± 0.09 | 0.57 ± 0.08 | 53.8 | 42.9 | 3.6 | |
| Trolox | 1.71 ± 0.11 | 0.61 ± 0.10 | 50.0 | 46.9 | 3.1 | |
References
- Nagata, M.P.B.; Egashira, J.; Katafuchi, N.; Endo, K.; Ogata, K.; Yamanaka, K.; Yamanouchi, T.; Matsuda, H.; Hashiyada, Y.; Yamashita, K. Bovine sperm selection procedure prior to cryopreservation for improvement of post-thawed semen quality and fertility. J. Anim. Sci. Biotechnol. 2019, 10, 91. [Google Scholar] [CrossRef]
- Alvarez, M.; Anel, L.L.; Boixo, J.C.; Chamorro, C.; Neila, M.M.; Montes, G.R.; Paz, P.; Anel, L. Current challenges in sheep artificial insemination: A particular insight. Reprod. Domest. Anim. 2019, 54, 32–40. [Google Scholar] [CrossRef]
- Gibbons, A.E.; Fernandez, J.; Bruno, G.M.M.; Spinelli, M.V.; Cueto, M.I.; Gibbons, A.E.; Fernandez, J.; Bruno, G.M.M.; Spinelli, M.V.; Cueto, M.I.; et al. Technical recommendations for artificial insemination in sheep. Anim. Reprod. 2019, 16, 803–809. [Google Scholar] [CrossRef]
- Lv, C.; Wu, G.; Hong, Q.; Quan, G. Spermatozoa Cryopreservation: State of Art and Future in Small Ruminants. Biopreserv. Biobank. 2019, 17, 171–182. [Google Scholar] [CrossRef]
- Koppers, A.J.; Iuliis, G.N.; Finnie, J.M.; Laughlin, E.A.; Aitken, R.J. Significance of Mitochondrial Reactive Oxygen Species in the Generation of Oxidative Stress in Spermatozoa. J. Clin. Endocrinol. Metab. 2008, 93, 3199–3207. [Google Scholar] [CrossRef]
- Martin, H.D.; Bragado, M.J.; Batista, A.R.; Oliveira, P.F.; Alves, M.G. Antioxidants and male fertility: From molecular studies to clinical evidence. Antioxidants 2019, 8, 89. [Google Scholar] [CrossRef]
- Len, J.S.; Koh, W.S.D.; Tan, S.X. The roles of reactive oxygen species and antioxidants in cryopreservation. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef]
- Peña, F.J.; Flaherty, C.; Ortiz, R.J.M.; Martín, C.F.E.; Gaitskell, P.G.L.; Gil, M.C.; Ferrusola, C.O.; Flaherty, C.; Ortiz, R.J.M.; Martín, C.F.E.; et al. Redox regulation and oxidative stress: The particular case of the stallion spermatozoa. Antioxidants 2019, 8, 567. [Google Scholar] [CrossRef]
- Ollero, M.; Powers, R.D.; Alvarez, J.G. Variation of docosahexaenoic acid content in subsets of human spermatozoa at different stages of maturation: Implications for sperm lipoperoxidative damage. Mol. Reprod. Dev. 2000, 55, 326–334. [Google Scholar] [CrossRef]
- Alahmar, A.T. Role of Oxidative Stress in Male Infertility: An Updated Review. J. Hum. Reprod. Sci. 2019, 12, 4–18. [Google Scholar] [CrossRef]
- Ko, E.Y.; Sabanegh, E.S.; Agarwal, A. Male infertility testing: Reactive oxygen species and antioxidant capacity. Fertil. Steril. 2014, 102, 1518–1527. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Saleh, R.A.; Bedaiwy, M.A. Role of reactive oxygen species in the pathophysiology of human reproduction. Fertil. Steril. 2003, 79, 829–843. [Google Scholar] [CrossRef]
- Rajesh, K.T.; Doreswamy, K.; Muralidhara, S.B. Oxidative stress associated DNA damage in testis of mice: Induction of abnormal sperms and effects on fertility. Mutat. Res.Genet. Toxicol. Environ. Mutagen. 2001, 513, 103–111. [Google Scholar] [CrossRef]
- Aitken, R.J. Impact of oxidative stress on male and female germ cells: Implications for fertility. Reproduction 2020, 159, R189–R201. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Majzoub, A.; Agarwal, A. Oxidative stress and sperm function: A systematic review on evaluation and management. Arab J. Urol. 2019, 17, 87–97. [Google Scholar] [CrossRef]
- Cocuzza, M.; Sikka, S.C.; Athayde, K.S.; Agarwal, A. Clinical relevance of oxidative stress and sperm chromation damage in male infertility: An evidence based analysis. Int. Braz. J. Urol. 2007, 33, 603–621. [Google Scholar] [CrossRef] [PubMed]
- Allai, L.; Benmoula, A.; Silva, M.M.; Nasser, B.; Amiri, B. Supplementation of ram semen extender to improve seminal quality and fertility rate. Anim. Reprod. Sci. 2018, 192, 6–17. [Google Scholar] [CrossRef]
- Agarwal, A.; Durairajanayagam, D.; Plessis, S.S. Utility of antioxidants during assisted reproductive techniques: An evidence based review. Reprod. Biol. Endocrinol. 2014, 12. [Google Scholar] [CrossRef]
- Amidi, F.; Pazhohan, A.; Shabani, N.M.; Khodarahmian, M.; Nekoonam, S. The role of antioxidants in sperm freezing: A review. Cell Tissue Bank. 2016, 17, 745–756. [Google Scholar] [CrossRef]
- Zhang, W.; Yi, K.; Chen, C.; Hou, X.; Zhou, X. Application of antioxidants and centrifugation for cryopreservation of boar spermatozoa. Anim. Reprod. Sci. 2012, 132, 123–128. [Google Scholar] [CrossRef]
- Peris, F.P.; Soler, A.J.; Iniesta, C.M.; Martín, M.A.; Sánchez, A.I.; Medina, C.D.A.; Fernández, S.M.R.; García, Á.O.; Maroto, M.A.; Montoro, V.; et al. Sperm Cryodamage in Ruminants: Understanding the Molecular Changes Induced by the Cryopreservation Process to Optimize Sperm Quality. Int. J. Mol. Sci. 2020, 21, 2781. [Google Scholar] [CrossRef]
- López, A.L.; Alvarez, G.O.; Morales, M.A.; Cuerda, I.M.; Ramón, M.; Soler, A.J.; Santos, F.M.R.; Garde, J.J. Reduced glutathione addition improves both the kinematics and physiological quality of post-thawed red deer sperm. Anim. Reprod. Sci. 2015, 162, 73–79. [Google Scholar] [CrossRef] [PubMed]
- López, A.L.; Álvarez, G.O.; Parrilla, I.; Olmo, D.; Santos, F.M.; Soler, A.; Morales, M.A.; Ortiz, J.; Alkmin, D.; Tarantini, T.; et al. The Effect of Oxidative Stress on Thawed Bulk-Sorted Red Deer Sperm. Reprod. Domest. Anim. 2016, 51, 407–414. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Agarwal, A.; Selvam, M.K.P.; Samanta, L.; Vij, S.C.; Parekh, N.; Sabanegh, E.; Tadros, N.N.; Arafa, M.; Sharma, R. Effect of antioxidant supplementation on the sperm proteome of idiopathic infertile men. Antioxidants 2019, 8, 488. [Google Scholar] [CrossRef]
- Meara, C.M.; Hanrahan, J.P.; Donovan, A.; Fair, S.; Rizos, D.; Wade, M.; Boland, M.P.; Evans, A.C.O.; Lonergan, P. Relationship between in vitro fertilisation of ewe oocytes and the fertility of ewes following cervical artificial insemination with frozen-thawed ram semen. Theriogenology 2005, 64, 1797–1808. [Google Scholar] [CrossRef]
- Agarwal, A.; Roychoudhury, S.; Sharma, R.; Gupta, S.; Majzoub, A.; Sabanegh, E. Diagnostic application of oxidation-reduction potential assay for measurement of oxidative stress: Clinical utility in male factor infertility. Reprod. Biomed. Online 2017, 34, 48–57. [Google Scholar] [CrossRef]
- Campuzano, M.M.; Rodríguez, A.M.; Álvarez, M.; Canul, T.J.; Anel, L.; Paz, P.; Pastor, M.F. Post-thawing quality and incubation resilience of cryopreserved ram spermatozoa are affected by antioxidant supplementation and choice of extender. Theriogenology 2015, 83, 520–528. [Google Scholar] [CrossRef]
- Anel, L.; Paz, P.; Álvarez, M.; Chamorro, C.A.; Boixo, J.C.; Manso, A.; González, M.; Kaabi, M.; Anel, E. Field and in vitro assay of three methods for freezing ram semen. Theriogenology 2003, 60, 1293–1308. [Google Scholar] [CrossRef]
- Riesco, F.M.; Lopez, A.L.; Montero, N.M.; Martinez, P.C.; Garrido, M.R.; Alvarez, M.; Paz, P.; Anel, L. ProAKAP4 as novel molecular marker of sperm quality in ram: An integrative study in fresh, cooled and cryopreserved sperm. Biomolecules 2020, 10, 1046. [Google Scholar] [CrossRef]
- Chávez, C.J.M.; Cerón, H.J.; Flores, A.C.F.; Carlos, L.M.A.; Domínguez, L.R.R.; Casasola, Q.A.; Cháirez, E.F.G. Lambing rate and prolificacy in inseminated hair sheep treated with bovine somatotropin. Trop. Anim. Health Prod. 2016, 48, 699–703. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, F.; Feng, X.; Yang, H.; Zhu, A.; Pang, J.; Han, L.; Zhang, T.; Yao, X.; Wang, F. Genome-wide analysis of DNA Methylation profiles on sheep ovaries associated with prolificacy using whole-genome Bisulfite sequencing. BMC Genom. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, R.; Bostofte, E.; Skakkebæk, N.E.; Hansen, J.; Møller, H. Offspring sex ratio of subfertile men and men with abnormal sperm characteristics. Hum. Reprod. 2000, 15, 2369–2370. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Crespo, P.M.; Pintado, B.; Adán, G.A. Scrotal heat stress effects on sperm viability, sperm DNA integrity, and the offspring sex ratio in mice. Mol. Reprod. Dev. 2008, 75, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Saleh, R.A.; Agarwal, A. Oxidative stress and male infertility: From research bench to clinical practice. J. Androl. 2002, 23, 737–752. [Google Scholar] [CrossRef]
- Ghallab, A.R.M.; Shahat, A.M.; Fadl, A.M.; Ayoub, M.M.; Moawad, A.R. Impact of supplementation of semen extender with antioxidants on the quality of chilled or cryopreserved Arabian stallion spermatozoa. Cryobiology 2017, 79, 14–20. [Google Scholar] [CrossRef]
- Gadani, B.; Bucci, D.; Spinaci, M.; Tamanini, C.; Galeati, G. Resveratrol and Epigallocatechin-3-gallate addition to thawed boar sperm improves in vitro fertilization. Theriogenology 2017, 90, 88–93. [Google Scholar] [CrossRef]
- Eidan, S.M. Effect on post-cryopreserved semen characteristics of Holstein bulls of adding combinations of vitamin C and either catalase or reduced glutathione to Tris extender. Anim. Reprod. Sci. 2016, 167, 1–7. [Google Scholar] [CrossRef]
- Jamadi, S.A.; Ahmad, E.; Ansari, M.; Kohram, H. Antioxidant effect of quercetin in an extender containing DMA or glycerol on freezing capacity of goat semen. Cryobiology 2017, 75, 15–20. [Google Scholar] [CrossRef]
- López, A.L.; Rodríguez, A.M.; García, A.O.; Álvarez, M.; Morales, M.A.; Anel, L.; Paz, P.; Garde, J.J.; Pastor, M.F. Reduced glutathione and Trolox (vitamin E) as extender supplements in cryopreservation of red deer epididymal spermatozoa. Anim. Reprod. Sci. 2012, 135, 37–46. [Google Scholar] [CrossRef]
- Bae, J.; Kim, S.; Chen, Z.; Eisenberg, M.L.; Buck, L.G.M. Human semen quality and the secondary sex ratio. Asian J. Androl. 2016, 18, 374. [Google Scholar] [CrossRef]
- Mossman, J.A.; Slate, J.; Birkhead, T.R.; Moore, H.D.; Pacey, A.A. Sperm speed is associated with sex bias of siblings in a human population. Asian J. Androl. 2013, 15, 152–154. [Google Scholar] [CrossRef] [PubMed]
- Maligaya, M.L.R.; Chan, C.A.; Jacobson, J.D.; Patton, W.C.; Corselli, J.; Chan, P.J. A follow-up expanded study of the correlation of sperm velocity in seminal plasma and offspring gender. Arch. Androl. 2006, 52, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Sailer, B.L.; Sarkar, L.J.; Bjordahl, J.A.; Jost, L.K.; Evenson, D.P. Effects of heat stress on mouse testicular cells and sperm chromatin structure. J. Androl. 1997, 18, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Rathje, C.C.; Johnson, E.E.P.; Drage, D.; Patinioti, C.; Silvestri, G.; Affara, N.A.; Radio, I.C.; Cocquet, J.; Skinner, B.M.; Ellis, P.J.I. Differential Sperm Motility Mediates the Sex Ratio Drive Shaping Mouse Sex Chromosome Evolution. Curr. Biol. 2019, 29, 3692–3698. [Google Scholar] [CrossRef] [PubMed]
- Santolaria, P.; Palacin, I.; Yaniz, J. Management Factors Affecting Fertility in Sheep. In Artificial Insemination in Farm Animals; InTech: London, UK, 2011. [Google Scholar]
- Fiel, V.S.; Palacín, I.; Santolaria, P.; Yániz, J.L. A comparative study of sperm morphometric subpopulations in cattle, goat, sheep and pigs using a computer-assisted fluorescence method (CASMA-F). Anim. Reprod. Sci. 2013, 139, 182–189. [Google Scholar] [CrossRef]
- Brito, L.F.C.; Barth, A.D.; Goeseels, B.S.; Panich, P.L.; Kastelic, J.P. Comparison of methods to evaluate the plasmalemma of bovine sperm and their relationship with in vitro fertilization rate. Theriogenology 2003, 60, 1539–1551. [Google Scholar] [CrossRef]
- Plaza, D.M.; Muñoz, M.P.; Tapia, J.A.; Ortega, F.C.; Silva, B.C.C.; Peña, F.J.; Davila, M.P.; Muñoz, P.M.; Tapia, J.A. Inhibition of mitochondrial complex I leads to decreased motility and membrane integrity related to increased hydrogen peroxide and reduced ATP production, while the inhibition of glycolysis has less impact on sperm motility. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Agarwal, A.; Qiu, E.; Sharma, R. Laboratory assessment of oxidative stress in semen. Arab J. Urol. 2018, 16, 77–86. [Google Scholar] [CrossRef]
- Agarwal, A.; Roychoudhury, S.; Bjugstad, K.B.; Cho, C.L. Oxidation-reduction potential of semen: What is its role in the treatment of male infertility? Ther. Adv. Urol. 2016, 8, 302–318. [Google Scholar] [CrossRef]
- Aitken, R.J. Not every sperm is sacred; a perspective on male infertility. Mol. Hum. Reprod. 2018, 24, 287–298. [Google Scholar] [CrossRef]
- Daas, J.H.G.; Jong, G.; Lansbergen, L.M.T.E.; Wagtendonk, L.A.M. The Relationship between the Number of Spermatozoa Inseminated and the Reproductive Efficiency of Individual Dairy Bulls. J. Dairy Sci. 1998, 81, 1714–1723. [Google Scholar] [CrossRef]
- Ostermeier, G.C.; Sargeant, G.A.; Yandell, B.S.; Evenson, D.P.; Parrish, J.J. Relationship of bull fertility to sperm nuclear shape. J. Androl. 2001, 22, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Saacke, R.G. Sperm morphology: Its relevance to compensable and uncompensable traits in semen. Theriogenology 2008, 70, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Fiel, V.S.; Palacín, I.; Santolaria, P.; Fantova, E.; Casorrán, Q.F.J.; Mur, S.E.; Yániz, J.L. In vitro assessment of sperm quality from rams of high and low field fertility. Anim. Reprod. Sci. 2014, 146, 15–20. [Google Scholar] [CrossRef]
- Aitken, R.J.; Smith, T.B.; Jobling, M.S.; Baker, M.A.; Iuliis, G.N. Oxidative stress and male reproductive health. Asian J. Androl. 2014, 16, 31–38. [Google Scholar] [CrossRef]
- Maia, M.S.; Bicudo, S.D.; Sicherle, C.C.; Rodello, L.; Gallego, I.C.S. Lipid peroxidation and generation of hydrogen peroxide in frozen-thawed ram semen cryopreserved in extenders with antioxidants. Anim. Reprod. Sci. 2010, 122, 118–123. [Google Scholar] [CrossRef]
- Nekoonam, S.; Nashtaei, M.S.; Naji, M.; Zangi, B.M.; Amidi, F. Effect of Trolox on sperm quality in normozospermia and oligozospermia during cryopreservation. Cryobiology 2016, 72, 106–111. [Google Scholar] [CrossRef]
- Silva, S.V.; Soares, A.T.; Batista, A.M.; Almeida, F.C.; Nunes, J.F.; Peixoto, C.A.; Guerra, M.M.P. Vitamin E (Trolox) addition to Tris-egg yolk extender preserves ram spermatozoon structure and kinematics after cryopreservation. Anim. Reprod. Sci. 2013, 137, 37–44. [Google Scholar] [CrossRef]
- Zhu, Z.; Fan, X.; Lv, Y.; Zhang, N.; Fan, C.; Zhang, P.; Zeng, W. Vitamin E analogue improves rabbit sperm quality during the process of cryopreservation through its antioxidative action. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riesco, M.F.; Alvarez, M.; Anel-Lopez, L.; Neila-Montero, M.; Palacin-Martinez, C.; Montes-Garrido, R.; Boixo, J.C.; de Paz, P.; Anel, L. Multiparametric Study of Antioxidant Effect on Ram Sperm Cryopreservation—From Field Trials to Research Bench. Animals 2021, 11, 283. https://doi.org/10.3390/ani11020283
Riesco MF, Alvarez M, Anel-Lopez L, Neila-Montero M, Palacin-Martinez C, Montes-Garrido R, Boixo JC, de Paz P, Anel L. Multiparametric Study of Antioxidant Effect on Ram Sperm Cryopreservation—From Field Trials to Research Bench. Animals. 2021; 11(2):283. https://doi.org/10.3390/ani11020283
Chicago/Turabian StyleRiesco, Marta F., Mercedes Alvarez, Luis Anel-Lopez, Marta Neila-Montero, Cristina Palacin-Martinez, Rafael Montes-Garrido, Juan Carlos Boixo, Paulino de Paz, and Luis Anel. 2021. "Multiparametric Study of Antioxidant Effect on Ram Sperm Cryopreservation—From Field Trials to Research Bench" Animals 11, no. 2: 283. https://doi.org/10.3390/ani11020283
APA StyleRiesco, M. F., Alvarez, M., Anel-Lopez, L., Neila-Montero, M., Palacin-Martinez, C., Montes-Garrido, R., Boixo, J. C., de Paz, P., & Anel, L. (2021). Multiparametric Study of Antioxidant Effect on Ram Sperm Cryopreservation—From Field Trials to Research Bench. Animals, 11(2), 283. https://doi.org/10.3390/ani11020283