Heat Stress Reduces Metabolic Rate While Increasing Respiratory Exchange Ratio in Growing Pigs

 ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Indirect Calorimetry
2.3. Skeletal Muscle Metabolic Measures
2.4. Statistical Analysis
3. Results
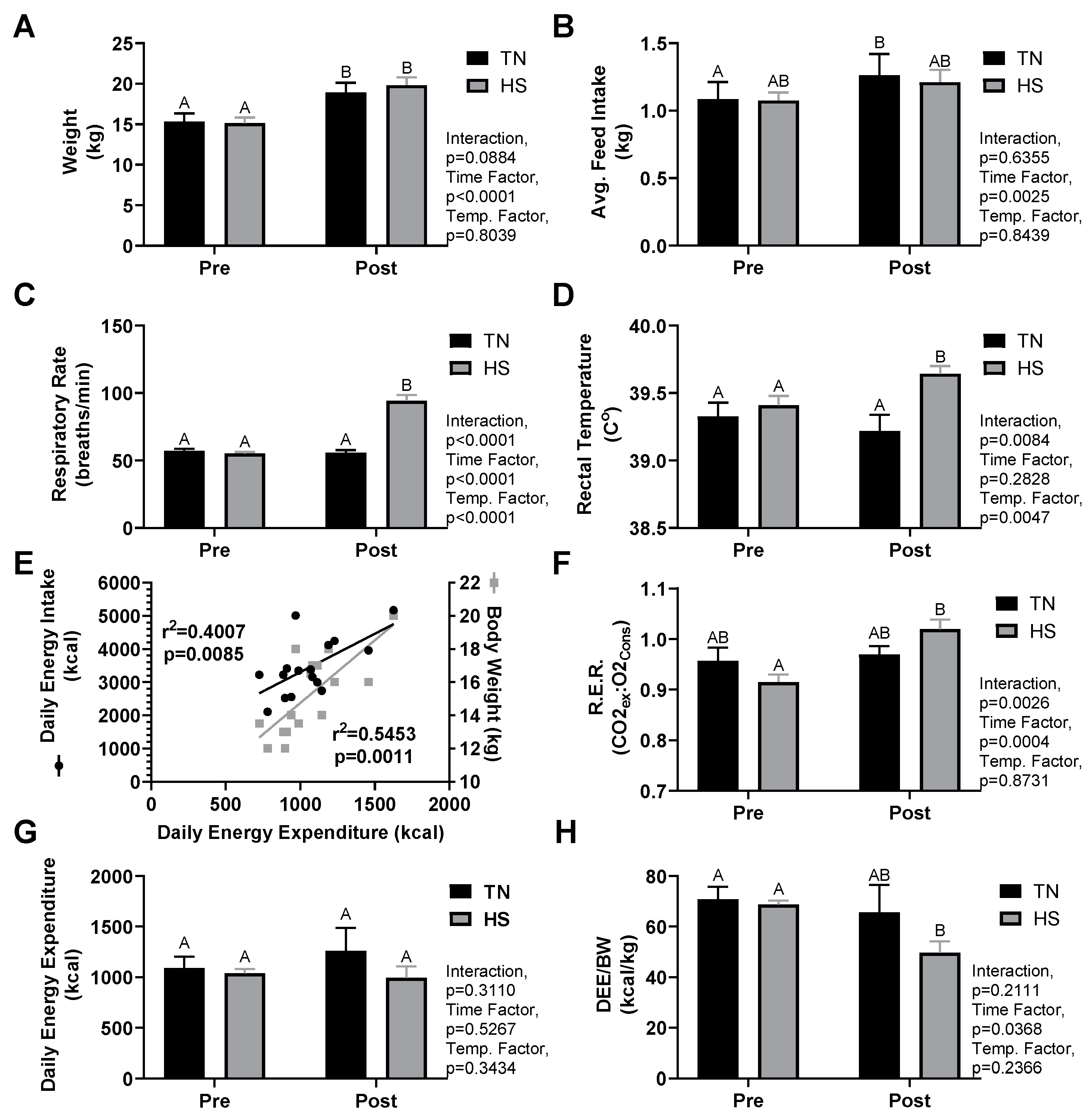
3.1. Growth and Thermoregulatory Response
3.2. Indirect Calorimetry
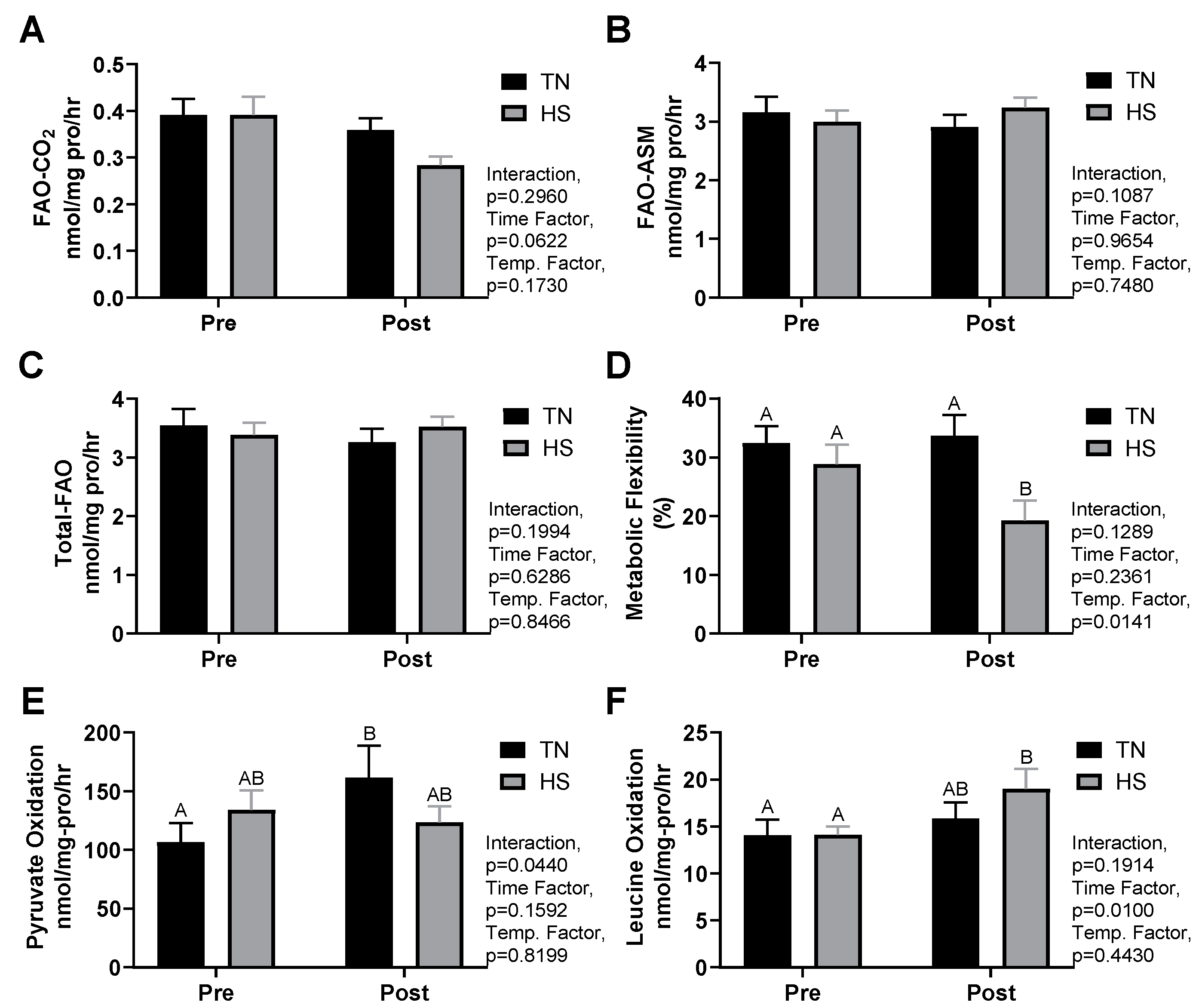
3.3. Substrate Metabolism
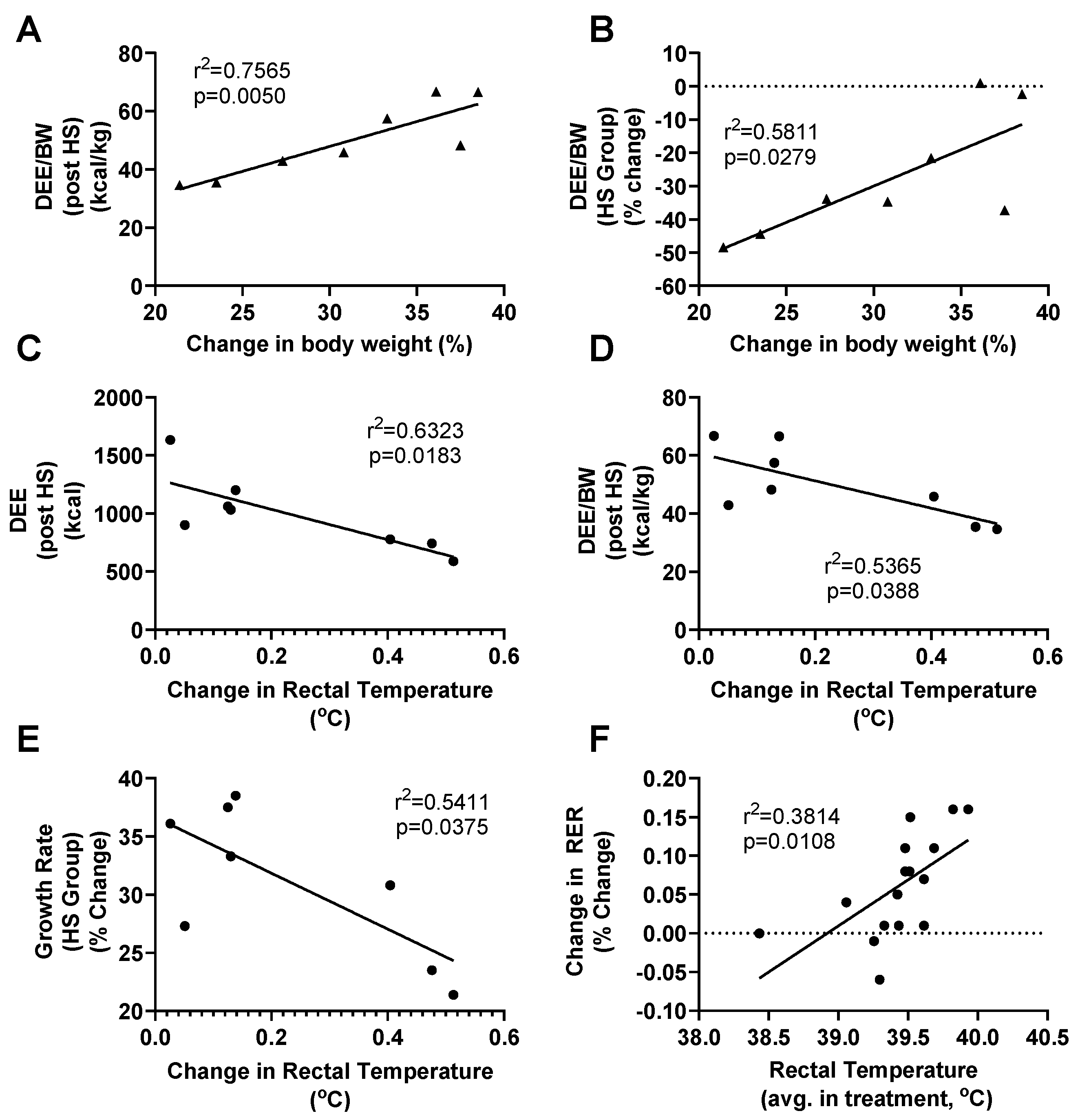
3.4. HS Metabolism and Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, H.; Jiang, R.; Xu, S.; Zhang, Z.; Xu, G.; Zheng, J.; Qu, L. Transcriptome responses to heat stress in hypothalamus of a meat-type chicken. J. Anim. Sci. Biotechnol. 2015, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; McMillan, R.P.; Xie, G.; Giridhar, S.; Baumgard, L.H.; El-Kadi, S.; Selsby, J.; Ross, J.; Gabler, N.; Hulver, M.W.; et al. Heat stress decreases metabolic flexibility in skeletal muscle of growing pigs. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R1096–R1106. [Google Scholar] [CrossRef] [PubMed]
- He, J.; He, Y.; Pan, D.; Cao, J.; Sun, Y.; Zeng, X. Associations of Gut Microbiota with Heat Stress-Induced Changes of Growth, Fat Deposition, Intestinal Morphology, and Antioxidant Capacity in Ducks. Front. Microbiol. 2019, 10, 903. [Google Scholar] [CrossRef] [PubMed]
- Bernabucci, U.; Lacetera, N.; Baumgard, L.H.; Rhoads, R.P.; Ronchi, B.; Nardone, A. Metabolic and hormonal acclimation to heat stress in domesticated ruminants. Animal 2010, 4, 1167–1183. [Google Scholar] [CrossRef] [PubMed]
- Lkhagvasuren, B.; Nakamura, Y.; Oka, T.; Sudo, N.; Nakamura, K. Social defeat stress induces hyperthermia through activation of thermoregulatory sympathetic premotor neurons in the medullary raphe region. Eur. J. Neurosci. 2011, 34, 1442–1452. [Google Scholar] [CrossRef]
- García-Díaz, D.F.; Campion, J.; Milagro, F.I.; Lomba, A.; Marzo, F.; Martínez, J.A. Chronic mild stress induces variations in locomotive behavior and metabolic rates in high fat fed rats. J. Physiol. Biochem. 2007, 63, 337–346. [Google Scholar] [CrossRef]
- Funakoshi, K.; Nakano, M.; Atobe, Y.; Goris, R.C.; Kadota, T.; Yazama, F. Differential development of TRPV1-expressing sensory nerves in peripheral organs. Cell Tissue Res. 2006, 323, 27–41. [Google Scholar] [CrossRef]
- Bertin, S.; Aoki-Nonaka, Y.; de Jong, P.R.; Nohara, L.L.; Xu, H.; Stanwood, S.R.; Srikanth, S.; Lee, J.; To, K.; Abramson, L.; et al. The ion channel TRPV1 regulates the activation and proinflammatory properties of CD4⁺ T cells. Nat. Immunol. 2014, 15, 1055–1063. [Google Scholar] [CrossRef]
- Wang, L.; Schumann, U.; Liu, Y.; Prokopchuk, O.; Steinacker, J.M. Heat shock protein 70 (Hsp70) inhibits oxidative phosphorylation and compensates ATP balance through enhanced glycolytic activity. J. Appl. Physiol. 2012, 113, 1669–1676. [Google Scholar] [CrossRef]
- Kuan, Y.C.; Hashidume, T.; Shibata, T.; Uchida, K.; Shimizu, M.; Inoue, J.; Sato, R. Heat Shock Protein 90 Modulates Lipid Homeostasis by Regulating the Stability and Function of Sterol Regulatory Element-binding Protein (SREBP) and SREBP Cleavage-activating Protein. J. Biol. Chem. 2017, 292, 3016–3028. [Google Scholar] [CrossRef]
- Marshall, J.P.S.; Estevez, E.; Kammoun, H.L.; King, E.J.; Bruce, C.R.; Drew, B.G.; Qian, H.; Iliades, P.; Gregorevic, P.; Febbraio, M.A.; et al. Skeletal muscle-specific overexpression of heat shock protein 72 improves skeletal muscle insulin-stimulated glucose uptake but does not alter whole body metabolism. Diabetes Obes. Metab. 2018, 20, 1928–1936. [Google Scholar] [CrossRef] [PubMed]
- Goto, A.; Egawa, T.; Sakon, I.; Oshima, R.; Ito, K.; Serizawa, Y.; Sekine, K.; Tsuda, S.; Goto, K.; Hayashi, T. Heat stress acutely activates insulin-independent glucose transport and 5’-AMP-activated protein kinase prior to an increase in HSP72 protein in rat skeletal muscle. Physiol. Rep. 2015, 3, e12601. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Tu, J.; Dou, C.; Zhang, J.; Yang, L.; Liu, X.; Lei, K.; Liu, Z.; Wang, Y.; Li, L.; et al. HSP90 promotes cell glycolysis, proliferation and inhibits apoptosis by regulating PKM2 abundance via Thr-328 phosphorylation in hepatocellular carcinoma. Mol. Cancer 2017, 16, 178. [Google Scholar] [CrossRef] [PubMed]
- Victoria Sanz Fernandez, M.; Johnson, J.S.; Abuajamieh, M.; Stoakes, S.K.; Seibert, J.T.; Cox, L.; Kahl, S.; Elsasser, T.H.; Ross, J.W.; Isom, S.C.; et al. Effects of heat stress on carbohydrate and lipid metabolism in growing pigs. Physiol. Rep. 2015, 3, e12315. [Google Scholar] [CrossRef] [PubMed]
- Pasiakos, S.M.; Vislocky, L.M.; Carbone, J.W.; Altieri, N.; Konopelski, K.; Freake, H.C.; Anderson, J.M.; Ferrando, A.A.; Wolfe, R.R.; Rodriguez, N.R. Acute Energy Deprivation Affects Skeletal Muscle Protein Synthesis and Associated Intracellular Signaling Proteins in Physically Active Adults. J. Nutr. 2010, 140, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Areta, J.L.; Burke, L.M.; Camera, D.M.; West, D.W.D.; Crawshay, S.; Moore, D.R.; Stellingwerff, T.; Phillips, S.M.; Hawley, J.A.; Coffey, V.G. Reduced resting skeletal muscle protein synthesis is rescued by resistance exercise and protein ingestion following short-term energy deficit. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E989–E997. [Google Scholar] [CrossRef]
- Gumucio, J.P.; Qasawa, A.H.; Ferrara, P.J.; Malik, A.N.; Funai, K.; McDonagh, B.; Mendias, C.L. Reduced mitochondrial lipid oxidation leads to fat accumulation in myosteatosis. bioRxiv 2019, 33, 7863–7881. [Google Scholar] [CrossRef]
- Pearce, S.C.; Lonergan, S.M.; Huff-Lonergan, E.; Baumgard, L.H.; Gabler, N.K. Acute Heat Stress and Reduced Nutrient Intake Alter Intestinal Proteomic Profile and Gene Expression in Pigs. PLoS ONE 2015, 10, e0143099. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Rhoads, R.P., Jr. Effects of Heat Stress on Postabsorptive Metabolism and Energetics. Annu. Rev. Anim. Biosci. 2013, 1, 311–337. [Google Scholar] [CrossRef]
- Horton, T.J.; Drougas, H.; Brachey, A.; Reed, G.W.; Peters, J.C.; Hill, J.O. Fat and carbohydrate overfeeding in humans: Different effects on energy storage. Am. J. Clin. Nutr. 1995, 62, 19–29. [Google Scholar] [CrossRef]
- Joosen, A.M.C.P.; Westerterp, K.R. Energy expenditure during overfeeding. Nutr. Metab. 2006, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Cramer, M.N.; Jay, O. Biophysical aspects of human thermoregulation during heat stress. Auton. Neurosci. 2016, 196, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Seibert, J.T.; Graves, K.L.; Hale, B.J.; Keating, A.F.; Baumgard, L.H.; Ross, J.W. Characterizing the acute heat stress response in gilts: I. Thermoregulatory and production variables. J. Anim. Sci. 2018, 96, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Ghassemi Nejad, J.; Sung, K.-I. Behavioral and physiological changes during heat stress in Corriedale ewes exposed to water deprivation. J. Anim. Sci. Technol. 2017, 59, 13. [Google Scholar] [CrossRef] [PubMed]
- Weir, J.B.d.V. New methods for calculating metabolic rate with special reference to protein metabolism. J. Physiol. 1949, 109, 1–9. [Google Scholar] [CrossRef]
- Tarpey, M.D.; Davy, K.P.; McMillan, R.P.; Bowser, S.M.; Halliday, T.M.; Boutagy, N.E.; Davy, B.M.; Frisard, M.I.; Hulver, M.W. Skeletal muscle autophagy and mitophagy in endurance-trained runners before and after a high-fat meal. Mol. Metab. 2017, 6, 1597–1609. [Google Scholar] [CrossRef] [PubMed]
- Sanz Fernandez, M.V.; Stoakes, S.K.; Abuajamieh, M.; Seibert, J.T.; Johnson, J.S.; Horst, E.A.; Rhoads, R.P.; Baumgard, L.H. Heat stress increases insulin sensitivity in pigs. Physiol. Rep. 2015, 3, e12478. [Google Scholar] [CrossRef]
- Kimball, A.L.; McCue, P.M.; Petrie, M.A.; Shields, R.K. Whole body heat exposure modulates acute glucose metabolism. Int. J. Hyperth. 2018, 35, 644–651. [Google Scholar] [CrossRef]
- Aarsland, A.; Chinkes, D.; Wolfe, R.R. Hepatic and whole-body fat synthesis in humans during carbohydrate overfeeding. Am. J. Clin. Nutr. 1997, 65, 1774–1782. [Google Scholar] [CrossRef]
- Minehira, K.; Vega, N.; Vidal, H.; Acheson, K.; Tappy, L. Effect of carbohydrate overfeeding on whole body macronutrient metabolism and expression of lipogenic enzymes in adipose tissue of lean and overweight humans. Int. J. Obes. 2004, 28, 1291–1298. [Google Scholar] [CrossRef]
- Dériaz, O.; Fournier, G.; Tremblay, A.; Després, J.P.; Bouchard, C. Lean-body-mass composition and resting energy expenditure before and after long-term overfeeding. Am. J. Clin. Nutr. 1992, 56, 840–847. [Google Scholar] [CrossRef] [PubMed]
- National Research Council (US) Subcommittee on Environmental Stress. Effect of Environment on Nutrient Requirements of Domestic Animals; National Academies Press (US): Washington, DC, USA, 1981. [Google Scholar]
- Mount, L.E. The metabolic rate of the new-born pig in relation to environmental temperature and to age. J. Physiol. 1959, 147, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Fuller, M.F.; Boyne, A.W. The effects of environmental temperature on the growth and metabolism of pigs given different amounts of food: 2. Energy metabolism. Br. J. Nutr. 1972, 28, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Saxton, C. Effects of severe heat stress on respiration and metabolic rate in resting man. Aviat. Space Environ. Med. 1981, 52, 281–286. [Google Scholar] [PubMed]
- Mayorga, E.J.; Renaudeau, D.; Ramirez, B.C.; Ross, J.W.; Baumgard, L.H. Heat stress adaptations in pigs. Anim. Front. 2018, 9, 54–61. [Google Scholar] [CrossRef]
- Begaye, B.; Vinales, K.L.; Hollstein, T.; Ando, T.; Walter, M.; Bogardus, C.; Krakoff, J.; Piaggi, P. Impaired Metabolic Flexibility to High-Fat Overfeeding Predicts Future Weight Gain in Healthy Adults. Diabetes 2020, 69, 181–192. [Google Scholar] [CrossRef]
- Bastardot, F.; Marques-Vidal, P.; Vollenweider, P. Association of body temperature with obesity. The CoLaus study. Int. J. Obes. 2019, 43, 1026–1033. [Google Scholar] [CrossRef]
- Heikens, M.J.; Gorbach, A.M.; Eden, H.S.; Savastano, D.M.; Chen, K.Y.; Skarulis, M.C.; Yanovski, J.A. Core body temperature in obesity. Am. J. Clin. Nutr. 2011, 93, 963–967. [Google Scholar] [CrossRef]
- Hibbard, K.A.; Hoffman, F.M.; Huntzinger, D.; West, T.O. Changes in land cover and terrestrial biogeochemistry. In Climate Science Special Report: Fourth National Climate Assessment; Wuebbles, D.J., Fahey, D.W., Hibbard, K.A., Dokken, D.J., Stewart, B.C., Maycock, T.K., Eds.; U.S. Global Change Research Program: Washington, DC, USA, 2017; Volume I, pp. 277–302. [Google Scholar] [CrossRef]
- Seelenbinder, K.M.; Zhao, L.D.; Hanigan, M.D.; Hulver, M.W.; McMillan, R.P.; Baumgard, L.H.; Selsby, J.T.; Ross, J.W.; Gabler, N.K.; Rhoads, R.P. Effects of heat stress during porcine reproductive and respiratory syndrome virus infection on metabolic responses in growing pigs. J. Anim. Sci. 2018, 96, 1375–1387. [Google Scholar] [CrossRef]
- Gueugneau, M.; Coudy-Gandilhon, C.; Théron, L.; Meunier, B.; Barboiron, C.; Combaret, L.; Taillandier, D.; Polge, C.; Attaix, D.; Picard, B.; et al. Skeletal Muscle Lipid Content and Oxidative Activity in Relation to Muscle Fiber Type in Aging and Metabolic Syndrome. J. Gerontol. Ser. A 2014, 70, 566–576. [Google Scholar] [CrossRef]
- Zurlo, F.; Lillioja, S.; Puente, A.E.-D.; Nyomba, B.L.; Raz, I.; Saad, M.F.; Swinburn, B.A.; Knowler, W.C.; Bogardus, C.; Ravussin, E. Low ratio of fat to carbohydrate oxidation as predictor of weight gain: Study of 24-h RQ. Am. J. Physiol. Endocrinol. Metab. 1990, 259, E650–E657. [Google Scholar] [CrossRef] [PubMed]
- Colditz, P.; Kellaway, R. The effect of diet and heat stress on feed intake, growth, and nitrogen metabolism in Friesian, and Brahman heifers. Aust. J. Agric. Res. 1972, 23, 717–725. [Google Scholar] [CrossRef]
- Gao, S.T.; Guo, J.; Quan, S.Y.; Nan, X.M.; Fernandez, M.V.S.; Baumgard, L.H.; Bu, D.P. The effects of heat stress on protein metabolism in lactating Holstein cows. J. Dairy Sci. 2017, 100, 5040–5049. [Google Scholar] [CrossRef] [PubMed]
- Simmons, P.S.; Miles, J.M.; Gerich, J.E.; Haymond, M.W. Increased proteolysis. An effect of increases in plasma cortisol within the physiologic range. J. Clin. Investig. 1984, 73, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Chung, N.; Jenkins, G.; Hannun, Y.A.; Heitman, J.; Obeid, L.M. Sphingolipids Signal Heat Stress-induced Ubiquitin-dependent Proteolysis. J. Biol. Chem. 2000, 275, 17229–17232. [Google Scholar] [CrossRef]
- Cruzen, S.M.; Baumgard, L.H.; Gabler, N.K.; Pearce, S.C.; Lonergan, S.M. Temporal proteomic response to acute heat stress in the porcine muscle sarcoplasm1. J. Anim. Sci. 2017, 95, 3961–3971. [Google Scholar] [CrossRef]
- Koch, F.; Lamp, O.; Eslamizad, M.; Weitzel, J.; Kuhla, B. Metabolic Response to Heat Stress in Late-Pregnant and Early Lactation Dairy Cows: Implications to Liver-Muscle Crosstalk. PLoS ONE 2016, 11, e0160912. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Measure | Thermal Neutral | Heat Stress | Interaction | Time Effect | Temperature Effect | ||
|---|---|---|---|---|---|---|---|
| Pre | Post | Pre | Post | ||||
| Weight (kg) | 15.3 ± 1.1 | 18.9 ± 1.3 * | 15.1 ± 0.7 | 19.8 ± 1.0 * | 0.0884 | <0.0001 | 0.8039 |
| Rectal Temp. (°C) | 39.3 ± 0.1 | 39.2 ± 0.1 | 39.4 ± 0.1 | 39.6 ± 0.1 *t | 0.0084 | 0.2828 | 0.0470 |
| Respiratory Rate (B.P.M.) | 57 ± 2 | 56 ± 2 | 55 ± 1 | 94 ± 4 *t | <0.0001 | <0.0001 | <0.0001 |
| Feed Intake (kg/day) | 1.1 ± 0.1 | 1.3 ± 0.2 * | 1.1 ± 0.1 | 1.2 ± 0.1 | 0.6355 | 0.0025 | 0.8439 |
| RER (CO2expired:O2consumed) | 0.95 ± 0.03 | 0.97 ± 0.01 | 0.91 ± 0.02 | 1.02 ± 0.02 * | 0.0026 | 0.0004 | 0.8731 |
| DEE (kcal) | 1090 ± 120 | 1252 ± 238 | 1038 ± 48 | 992 ± 123 | 0.3110 | 0.5267 | 0.3434 |
| DEE/BW (kcal/kg) | 70.9 ± 5.1 | 67.5 ± 11.6 | 68.8 ± 1.7 | 49.7 ± 4.8 * | 0.2111 | 0.0368 | 0.2366 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fausnacht, D.W.; Kroscher, K.A.; McMillan, R.P.; Martello, L.S.; Baumgard, L.H.; Selsby, J.T.; Hulver, M.W.; Rhoads, R.P. Heat Stress Reduces Metabolic Rate While Increasing Respiratory Exchange Ratio in Growing Pigs. Animals 2021, 11, 215. https://doi.org/10.3390/ani11010215
Fausnacht DW, Kroscher KA, McMillan RP, Martello LS, Baumgard LH, Selsby JT, Hulver MW, Rhoads RP. Heat Stress Reduces Metabolic Rate While Increasing Respiratory Exchange Ratio in Growing Pigs. Animals. 2021; 11(1):215. https://doi.org/10.3390/ani11010215
Chicago/Turabian StyleFausnacht, Dane W., Kellie A. Kroscher, Ryan P. McMillan, Luciane S. Martello, Lance H. Baumgard, Joshua T. Selsby, Matthew W. Hulver, and Robert P. Rhoads. 2021. "Heat Stress Reduces Metabolic Rate While Increasing Respiratory Exchange Ratio in Growing Pigs" Animals 11, no. 1: 215. https://doi.org/10.3390/ani11010215
APA StyleFausnacht, D. W., Kroscher, K. A., McMillan, R. P., Martello, L. S., Baumgard, L. H., Selsby, J. T., Hulver, M. W., & Rhoads, R. P. (2021). Heat Stress Reduces Metabolic Rate While Increasing Respiratory Exchange Ratio in Growing Pigs. Animals, 11(1), 215. https://doi.org/10.3390/ani11010215