Evaluation of Cross-Protection between G1a- and G2a-Genotype Porcine Epidemic Diarrhea Viruses in Suckling Piglets

 ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cells and Viruses
2.3. Growth Kinetics of G2a and G1a Strains of PEDV and Indirect Immunofluorescence Assay (IFA)
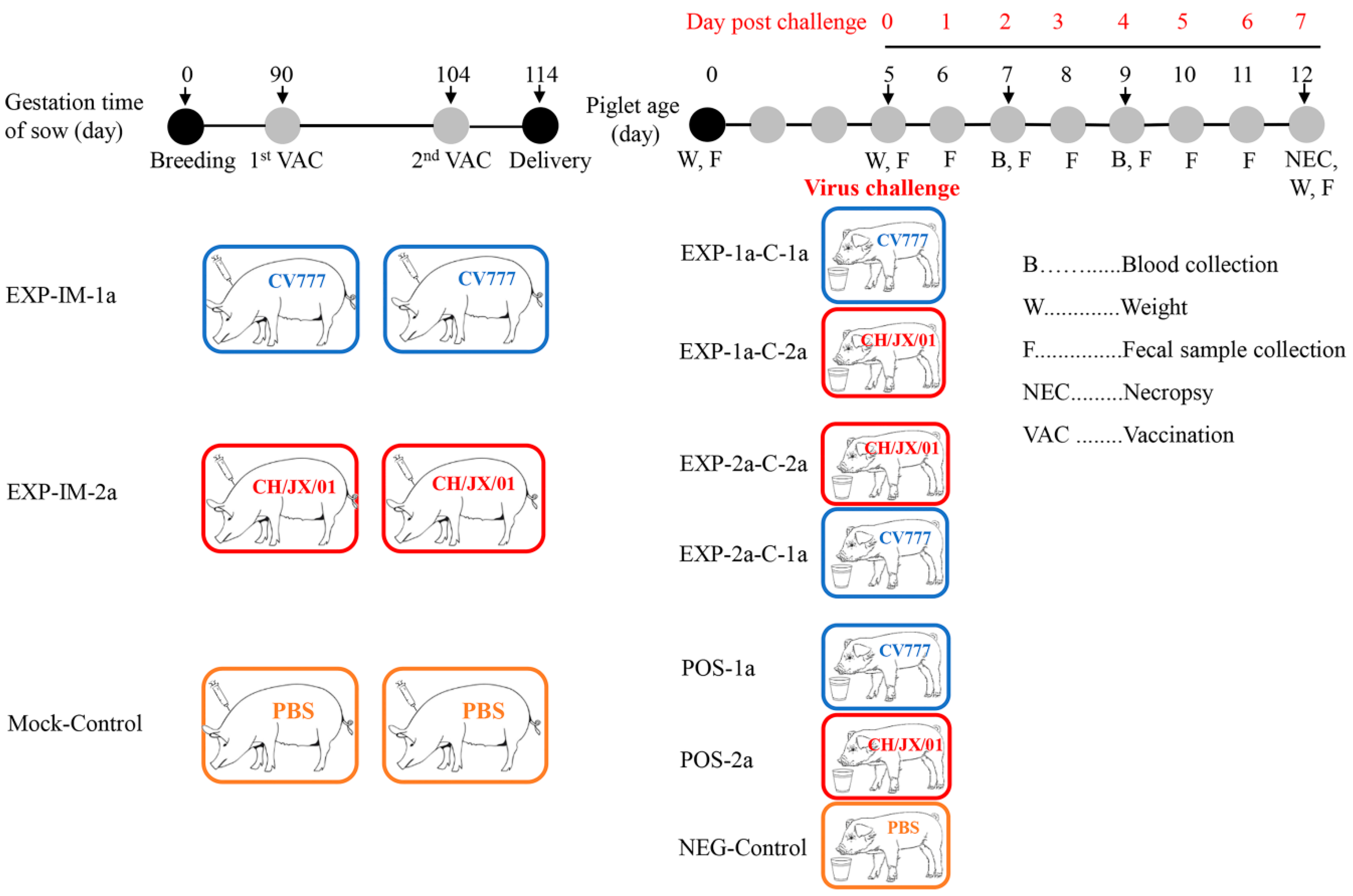
2.4. Animals
2.5. Vaccination and Challenge
2.6. Gross and Histopathological Examinations
2.7. Statistical Analysis
3. Results
3.1. Comparative Growth Kinetics of G2a and G1a PEDV Strains In Vitro
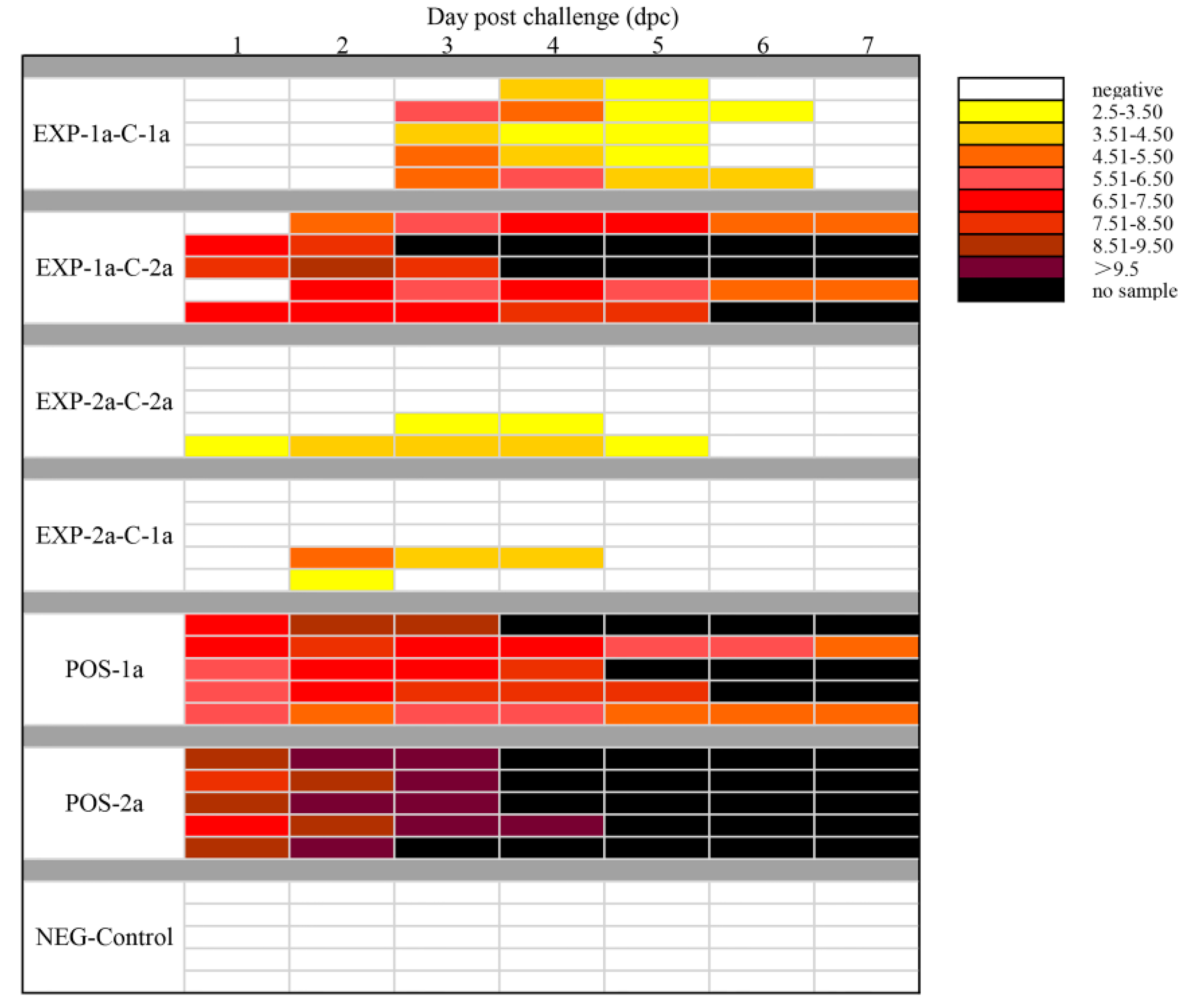
3.2. Clinical Assessment of Piglets under Virus Challenge
3.3. Gross Lesions, Histopathology, and Immunohistochemistry Staining
3.4. Protective Effects of Different Levels of Maternal Antibodies on Piglets
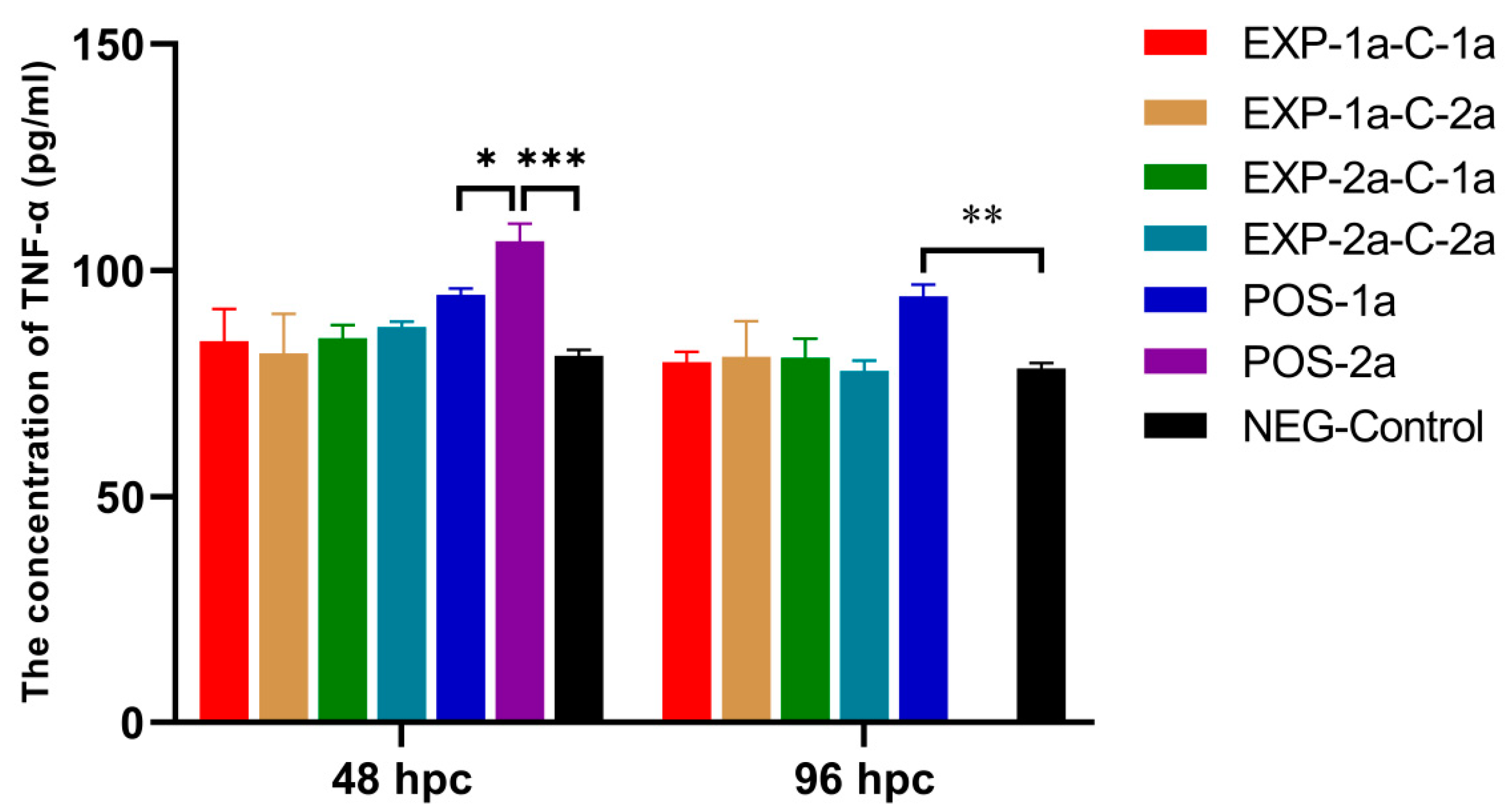
3.5. Serum TNF-α Concentration in Pigs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Daesub, S.; Bongkyun, P. Porcine epidemic diarrhoea virus: A comprehensive review of molecular epidemiology, diagnosis, and vaccines. Virus Genes 2012, 44, 167–175. [Google Scholar]
- Debouck, P.; Pensaert, M. Experimental infection of pigs with a new porcine enteric coronavirus, CV777. Am. J. Vet. Res. 1980, 41, 219. [Google Scholar] [PubMed]
- Ducatelle, R.; Coussement, W.; Charlier, G.; Debouck, P.; Hoorens, J. Three-dimensional Sequential Study of the Intestinal Surface in Experimental Porcine CV 777 Coronavirus Enteritis. Zent. Veterinärmed. Reihe B 2010, 28, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.-Q.; Cai, R.-J.; Chen, Y.-Q.; Liang, P.-S.; Chen, D.-K.; Song, C. Outbreak of Porcine Epidemic Diarrhea in Suckling Piglets, China. Emerg. Infect. Dis. 2012, 18, 161–163. [Google Scholar] [CrossRef]
- Stevenson, G.W.; Hoang, H.; Schwartz, K.J.; Burrough, E.; Madson, D.; Cooper, V.L.; Pillatzki, A.; Gauger, P.; Schmitt, B.J.; Koster, L.G.; et al. Emergence of Porcine epidemic diarrhea virus in the United States: Clinical signs, lesions, and viral genomic sequences. J. Vet. Diagn. Investig. 2013, 25, 649–654. [Google Scholar] [CrossRef]
- Zhang, F.; Luo, S.; Gu, J.; Li, Z.; Li, K.; Yuan, W.; Ye, Y.; Li, H.; Ding, Z.; Song, D.; et al. Prevalence and phylogenetic analysis of porcine diarrhea associated viruses in southern China from 2012 to 2018. BMC Vet. Res. 2019, 15, 1–9. [Google Scholar] [CrossRef]
- Masuda, T.; Murakami, S.; Takahashi, O.; Miyazaki, A.; Ohashi, S.; Yamasato, H.; Suzuki, T. New porcine epidemic diarrhoea virus variant with a large deletion in the spike gene identified in domestic pigs. Arch. Virol. 2015, 160, 2565–2568. [Google Scholar] [CrossRef]
- Dastjerdi, A.; Carr, J.; Ellis, R.J.; Steinbach, F.; Williamson, S. Porcine Epidemic Diarrhea Virus among Farmed Pigs, Ukraine. Emerg. Infect. Dis. 2015, 21, 2235–2237. [Google Scholar] [CrossRef]
- Oka, T.; Saif, L.J.; Marthaler, D.G.; Esseili, M.A.; Meulia, T.; Lin, C.-M.; Vlasova, A.; Jung, K.; Zhang, Y.; Wang, Q. Cell culture isolation and sequence analysis of genetically diverse US porcine epidemic diarrhea virus strains including a novel strain with a large deletion in the spike gene. Vet. Microbiol. 2014, 173, 258–269. [Google Scholar] [CrossRef]
- Suzuki, T.; Murakami, S.; Takahashi, O.; Kodera, A.; Masuda, T.; Itoh, S.; Miyazaki, A.; Ohashi, S.; Tsutsui, T. Molecular characterization of pig epidemic diarrhoea viruses isolated in Japan from 2013 to 2014. Infect. Genet. Evol. 2015, 36, 363–368. [Google Scholar] [CrossRef]
- Chen, Q.; Gauger, P.C.; Stafne, M.R.; Thomas, J.T.; Madson, D.M.; Huang, H.; Zheng, Y.; Li, G.; Zhang, J. Pathogenesis comparison between the United States porcine epidemic diarrhoea virus prototype and S-INDEL-variant strains in conventional neonatal piglets. J. Gen. Virol. 2016, 97, 1107–1121. [Google Scholar] [CrossRef]
- Leyi, W.; Beverly, B.; Yan, Z. New Variant of Porcine Epidemic Diarrhea Virus, United States, 2014. Emerg. Infect. Dis. 2014, 20, 917–919. [Google Scholar]
- Gallien, S.; Moro, A.; Lediguerher, G.; Catinot, V.; Paboeuf, F.; Bigault, L.; Gauger, P.C.; Pozzi, N.; Berri, M.; Authié, E.; et al. Limited shedding of an S-InDel strain of porcine epidemic diarrhea virus (PEDV) in semen and questions regarding the infectivity of the detected virus. Vet. Microbiol. 2019, 228, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Stadler, J.; Zoels, S. Emergence of porcine epidemic diarrhea virus in southern Germany. BMC Vet. Res. 2015, 11, 142. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-M.; Saif, L.J.; Marthaler, D.; Wang, Q. Evolution, antigenicity and pathogenicity of global porcine epidemic diarrhea virus strains. Virus Res. 2016, 226, 20–39. [Google Scholar] [CrossRef] [PubMed]
- Goede, D.; Murtaugh, M.P.; Nerem, J.; Yeske, P.; Rossow, K.; Morrison, R. Previous infection of sows with a ‘‘mild’’ strain of porcine epidemic diarrhea virus confers protection against infection with a ‘‘severe’’ strain. Vet. Microbiol. 2015, 176, 161–164. [Google Scholar] [CrossRef]
- Guo, J.; Fang, L.; Ye, X.; Chen, J.; Xu, S.; Zhu, X.; Miao, Y.; Wang, D.; Xiao, S. Evolutionary and genotypic analyses of global porcine epidemic diarrhea virus strains. Transbound. Emerg. Dis. 2018, 66, 111–118. [Google Scholar] [CrossRef]
- Song, D.; Huang, D.; Peng, Q.; Huang, T.; Chen, Y.; Zhang, T.; Nie, X.; He, H.; Wang, P.; Liu, Q.; et al. Molecular Characterization and Phylogenetic Analysis of Porcine Epidemic Diarrhea Viruses Associated with Outbreaks of Severe Diarrhea in Piglets in Jiangxi, China 2013. PLoS ONE 2015, 10, e0120310. [Google Scholar] [CrossRef]
- Song, D.; Moon, H.; Kang, B. Porcine epidemic diarrhea: A review of current epidemiology and available vaccines. Clin. Exp. Vaccine Res. 2015, 4, 166–176. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, L.; Zhang, Q.; Zhou, P.; Fang, Y.; Zhao, D.; Feng, J.; Li, W.; Zhang, Y.; Wang, Y. Evaluation and comparison of immunogenicity and cross-protective efficacy of two inactivated cell culture-derived GIIa- and GIIb-genotype porcine epidemic diarrhea virus vaccines in suckling piglets. Vet. Microbiol. 2019, 230, 278–282. [Google Scholar] [CrossRef]
- Opriessnig, T.; Gerber, P.F.; Shen, H.; De Castro, A.M.M.G.; Zhang, J.; Chen, Q.; Halbur, P.G. Evaluation of the efficacy of a commercial inactivated genogroup 2b-based porcine epidemic diarrhea virus (PEDV) vaccine and experimental live genogroup 1b exposure against 2b challenge. Vet. Res. 2017, 48, 69. [Google Scholar] [CrossRef]
- Chen, Q.; Thomas, J.T.; Giménez-Lirola, L.; Hardham, J.M.; Gao, Q.; Gerber, P.F.; Opriessnig, T.; Zheng, Y.; Li, G.; Gauger, P.C.; et al. Evaluation of serological cross-reactivity and cross-neutralization between the United States porcine epidemic diarrhea virus prototype and S-INDEL-variant strains. BMC Vet. Res. 2016, 12, 70. [Google Scholar] [CrossRef]
- Sato, T.; Oroku, K.; Ohshima, Y.; Furuya, Y.; Sasakawa, C. Efficacy of genogroup 1 based porcine epidemic diarrhea live vaccine against genogroup 2 field strain in Japan. Virol. J. 2018, 15, 28. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Oh, J.; Kang, B.; Yang, J.; Moon, H.; Yoo, H.; Jang, Y.; Park, B. Oral efficacy of Vero cell attenuated porcine epidemic diarrhea virus DR13 strain. Res. Vet. Sci. 2007, 82, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Li, H.; Bi, Z.; Song, D.; Zhang, F.; Lei, D.; Luo, S.; Li, Z.; Gong, W.; Huang, D.; et al. Significant inhibition of re-emerged and emerging swine enteric coronavirus in vitro using the multiple shRNA expression vector. Antivir. Res. 2019, 166, 11–18. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Q.; Zhang, L.; Zhou, P.; Yang, J.; Fang, Y.; Dong, Z.; Zhao, D.; Li, W.; Feng, J.; et al. A newly isolated Chinese virulent genotype GIIb porcine epidemic diarrhea virus strain: Biological characteristics, pathogenicity and immune protective effects as an inactivated vaccine candidate. Virus Res. 2019, 259, 18–27. [Google Scholar] [CrossRef]
- Zhao, Y.; Qu, H.; Hu, J.; Fu, J.; Chen, R.; Li, C.; Cao, S.; Wen, Y.; Wu, R.; Zhao, Q.; et al. Characterization and Pathogenicity of the Porcine Deltacoronavirus Isolated in Southwest China. Viruses 2019, 11, 1074. [Google Scholar] [CrossRef]
- Wang, X.; Chen, J.; Shi, D.; Shi, H.; Zhang, X.; Yuan, J.; Jiang, S.; Feng, L. Immunogenicity and antigenic relationships among spike proteins of porcine epidemic diarrhea virus subtypes G1 and G2. Arch. Virol. 2015, 161, 537–547. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, T.; Song, D.; Huang, T.; Peng, Q.; Chen, Y.; Li, A.; Zhang, F.; Wu, Q.; Ye, Y.; et al. Comparison and evaluation of conventional RT-PCR, SYBR green I and TaqMan real-time RT-PCR assays for the detection of porcine epidemic diarrhea virus. Mol. Cell. Probes 2017, 33, 36–41. [Google Scholar] [CrossRef]
- Bohac, J.; Derbyshire, J.B.; Thorsen, J. The detection of transmissible gastroenteritis viral antigens by immunodiffusion. Can. J. Comp. Med. 1975, 39, 67–75. [Google Scholar]
- Jung, K.; Wang, Q.; Scheuer, K.A.; Lu, Z.; Zhang, Y.; Saif, L.J. Pathology of US Porcine Epidemic Diarrhea Virus Strain PC21A in Gnotobiotic Pigs. Emerg. Infect. Dis. 2014, 20, 662–665. [Google Scholar] [CrossRef]
- Jung, K.; Saif, L.J. Porcine epidemic diarrhea virus infection: Etiology, epidemiology, pathogenesis and immunoprophylaxis. Vet. J. 2015, 204, 134–143. [Google Scholar] [CrossRef]
- Liu, X.; Lin, C.-M.; Annamalai, T.; Gao, X.; Lu, Z.; Esseili, M.A.; Jung, K.; El-Tholoth, M.; Saif, L.J.; Wang, Q. Determination of the infectious titer and virulence of an original US porcine epidemic diarrhea virus PC22A strain. Vet. Res. 2015, 46, 1–6. [Google Scholar] [CrossRef]
- Madson, D.M.; Arruda, P.H.E.; Magstadt, D.R.; Burrough, E.R.; Hoang, H.; Sun, D.; Bower, L.P.; Bhandari, M.; Gauger, P.C.; Stevenson, G.W. Characterization of Porcine Epidemic Diarrhea Virus Isolate US/Iowa/18984/2013 Infection in 1 Day Old Cesarean Derived Colostrum Deprived Piglets. Vet. Pathol. 2015, 53, 44–52. [Google Scholar] [CrossRef]
- Gerber, P.F.; Opriessnig, T. Detection of immunoglobulin (Ig) A antibodies against porcine epidemic diarrhea virus (PEDV) in fecal and serum samples. MethodsX 2015, 2, 368–373. [Google Scholar] [CrossRef]
- Lin, H.; Chen, L.; Gao, L.; Yuan, X.; Ma, Z.; Fan, H. Epidemic strain YC2014 of porcine epidemic diarrhea virus could provide piglets against homologous challenge. Virol. J. 2016, 13, 68. [Google Scholar] [CrossRef]
- Lin, C.-M.; Annamalai, T.; Liu, X.; Gao, X.; Lu, Z.; El-Tholoth, M.; Hu, H.; Saif, L.J.; Wang, Q. Experimental infection of a US spike-insertion deletion porcine epidemic diarrhea virus in conventional nursing piglets and cross-protection to the original US PEDV infection. Vet. Res. 2015, 46, 134. [Google Scholar] [CrossRef]
- Dinarello, C.A. Proinflammatory and Anti-inflammatory Cytokines as Mediators in the Pathogenesis of Septic Shock. Chest 1997, 112, 321S–329S. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, C.; Chen, Y.; Liu, Z.; Zheng, J.; Wang, T.; Luo, H.; Liu, Y.; Shan, Y.; Fang, W.; et al. Effect of route of inoculation on innate and adaptive immune responses to porcine epidemic diarrhea virus infection in suckling pigs. Vet. Microbiol. 2019, 228, 83–92. [Google Scholar] [CrossRef]
- Xue, M.; Zhao, J.; Ying, L.; Fu, F.; Li, L.; Ma, Y.; Shi, H.; Zhang, J.; Feng, L.; Liu, P. IL-22 suppresses the infection of porcine enteric coronaviruses and rotavirus by activating STAT3 signal pathway. Antivir. Res. 2017, 142, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Van Reeth, K.; Nauwynck, H. Proinflammatory cytokines and viral respiratory disease in pigs. Vet. Res. 2000, 31, 187–213. [Google Scholar] [CrossRef]
- Shen, Y.; Shenk, T.E. Viruses and apoptosis. Curr. Opin. Genet. Dev. 1995, 5, 105–111. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Challenge Group | Body Weight Gain in 7 dpc (kg) | Onset of Diarrhea (hpc) | Fecal Scores Accumulation and Ratio | Mortality | |||
|---|---|---|---|---|---|---|---|
| FS = 3 | FS = 2 | FS = 1 | FS = 0 | ||||
| EXP-1a-C-1a | 1.31 ± 0.38 | None a | 0 (0) | 0 (0) | 3 (4.29%) | 67 (95.71%) | 0 |
| EXP-1a-C-2a | −0.07 ± 0.19 * | 48–144 | 19 (38.78%) | 11 (22.45%) | 10 (20.41%) | 9 (18.37%) | 60% |
| EXP-2a-C-2a | 1.21 ± 0.11 | None b | 0 (0) | 0 (0) | 5 (7.14%) | 65 (78.57%) | 0 |
| EXP-2a-C-1a | 1.25 ± 0.64 | None | 0 (0) | 0 (0) | 4 (5.71%) | 66 (94.29%) | 0 |
| POS-1a | 0.80 ± 0.20 * | 48–144 | 28 (50.00%) | 12 (21.43%) | 10 (17.86%) | 6 (10.71%) | 60% |
| POS-2a | −0.35 ± 0.21 ** | 12–96 | 21 (67.74%) | 5 (16.13%) | 2 (6.45%) | 3 (9.68%) | 100% |
| NEG-Control | 1.41 ± 0.20 | None | 0 (0) | 0 (0) | 2 (2.86%) | 68 (97.14%) | 0 |
| Group | Villus Height (VH, μm) | Crypt Depth (CD, μm) | VH:CD Value | Intestine Wall Thickness (μm) | |
|---|---|---|---|---|---|
| Duodenum | EXP-1a-C-1a | 754.35 ± 39.96 C | 156.73 ± 6.78 bcAB | 4.81 | 356.82 ± 22.25 B |
| EXP-1a-C-2a | 214.33 ± 10.52 DE | 108.8 ± 6.76 dC | 1.97 | 317 ± 23.31 B | |
| EXP-2a-C-2a | 970.23 ± 61.35 B | 175.89 ± 11.41 abAB | 5.52 | 319 ± 12.17 B | |
| EXP-2a-C-1a | 958.64 ± 30.88 B | 169.26 ± 13.12 abAB | 5.66 | 310.15 ± 16.61 B | |
| POS-1a | 337.18 ± 54.45 D | 186.22 ± 6.42 aA | 1.81 | 307.04 ± 29.65 B | |
| POS-2a | 134.02 ± 9.93 E | 135 ± 4.24 cdBC | 0.99 | 134.72 ± 17.78 C | |
| NEG-Control | 1208.75 ± 56.74 A | 113.83 ± 8.71 dC | 10.62 | 751.5 ± 26.05 A | |
| Jejunum | EXP-1a-C-1a | 785.17 ± 20.46 bcB | 146.62 ± 6.41 aA | 5.36 | 215.43 ± 5.46 cBC |
| EXP-1a-C-2a | 180.33 ± 10.3 dC | 105.28 ± 6.37 bBC | 1.71 | 280.28 ± 19.16 bBC | |
| EXP-2a-C-2a | 965.17 ± 50.79 bB | 133 ± 6.99 aA | 7.26 | 215.15 ± 14.83 cBC | |
| EXP-2a-C-1a | 708.5 ± 50.17 cB | 146.78 ± 4.79 aA | 4.83 | 220.08 ± 9.78 cBC | |
| POS-1a | 140.3 ± 6.64 dC | 143.24 ± 10.4a A | 0.98 | 174.03 ± 15.36 cC | |
| POS-2a | 162.9 ± 12.04 dC | 135.91 ± 9.66 aA | 1.20 | 157.66 ± 6.43 cC | |
| NEG-Control | 1333.92 ± 93.33 aA | 93.5 ± 3.33 bC | 14.27 | 562.83 ± 26.79 aA | |
| Ileum | EXP-1a-C-1a | 1040.2 ± 20.01 B | 164.82 ± 8.97 aA | 6.31 | 299.48 ± 41.52 bcCD |
| EXP-1a-C-2a | 175.56 ± 10.4 D | 113.56 ± 7.33 bC | 1.55 | 314 ± 12.07 bcCD | |
| EXP-2a-C-2a | 380.58 ± 15.48 C | 162 ± 9.08 aA | 2.35 | 293.98 ± 37.04 bcCD | |
| EXP-2a-C-1a | 382.61 ± 53.40 C | 127.89 ± 6.58 bC | 2.99 | 371.56 ± 23.87 aCD | |
| POS-1a | 140.26 ± 8.26 D | 165.41 ± 12.26 aA | 0.85 | 289.91 ± 20 bcCD | |
| POS-2a | 124.72 ± 10.95 D | 128.37 ± 6.6b BC | 0.97 | 186.35 ± 5.6 cD | |
| NEG-Control | 1221.83 ± 36.38 A | 84 ± 4.48398 cD | 14.55 | 493.25 ± 51.14 aA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Chen, Y.; Yuan, W.; Peng, Q.; Zhang, F.; Ye, Y.; Huang, D.; Ding, Z.; Lin, L.; He, H.; et al. Evaluation of Cross-Protection between G1a- and G2a-Genotype Porcine Epidemic Diarrhea Viruses in Suckling Piglets. Animals 2020, 10, 1674. https://doi.org/10.3390/ani10091674
Zhang Y, Chen Y, Yuan W, Peng Q, Zhang F, Ye Y, Huang D, Ding Z, Lin L, He H, et al. Evaluation of Cross-Protection between G1a- and G2a-Genotype Porcine Epidemic Diarrhea Viruses in Suckling Piglets. Animals. 2020; 10(9):1674. https://doi.org/10.3390/ani10091674
Chicago/Turabian StyleZhang, Yuhan, Yanjun Chen, Weifeng Yuan, Qi Peng, Fanfan Zhang, Yu Ye, Dongyan Huang, Zhen Ding, Longhua Lin, Houjun He, and et al. 2020. "Evaluation of Cross-Protection between G1a- and G2a-Genotype Porcine Epidemic Diarrhea Viruses in Suckling Piglets" Animals 10, no. 9: 1674. https://doi.org/10.3390/ani10091674
APA StyleZhang, Y., Chen, Y., Yuan, W., Peng, Q., Zhang, F., Ye, Y., Huang, D., Ding, Z., Lin, L., He, H., Wu, Q., Song, D., & Tang, Y. (2020). Evaluation of Cross-Protection between G1a- and G2a-Genotype Porcine Epidemic Diarrhea Viruses in Suckling Piglets. Animals, 10(9), 1674. https://doi.org/10.3390/ani10091674