Analysis of XX SRY-Negative Sex Reversal Dogs

 ,
,  , and
, and 
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Ethical Statement
2.2. Cases
2.3. Morphological Analyses
2.4. Cytogenetic Analyses
2.5. Molecular Analyses
2.6. qPCR Analyses
2.7. Array-CGH Analyses
3. Results
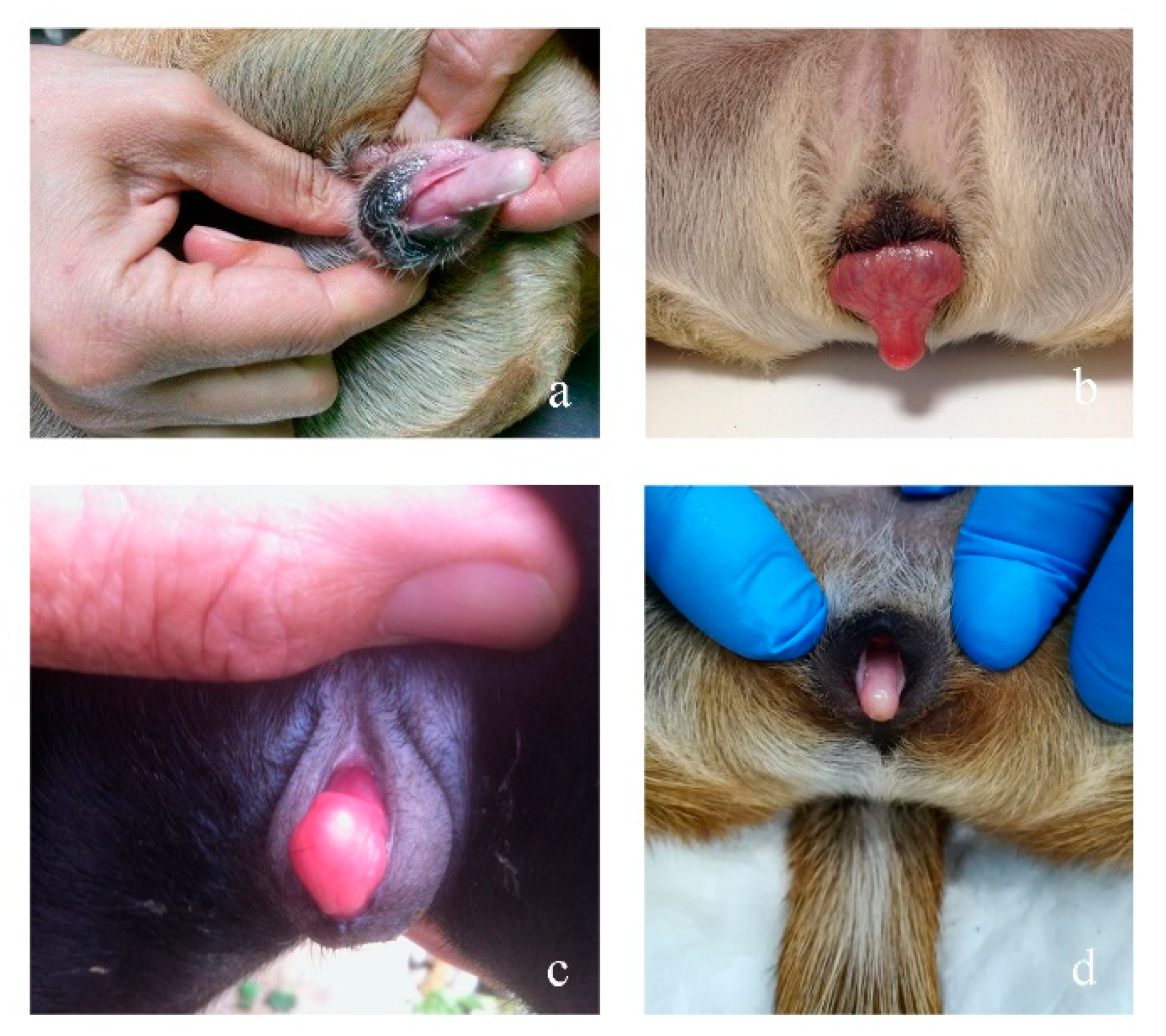
3.1. Clinical Findings
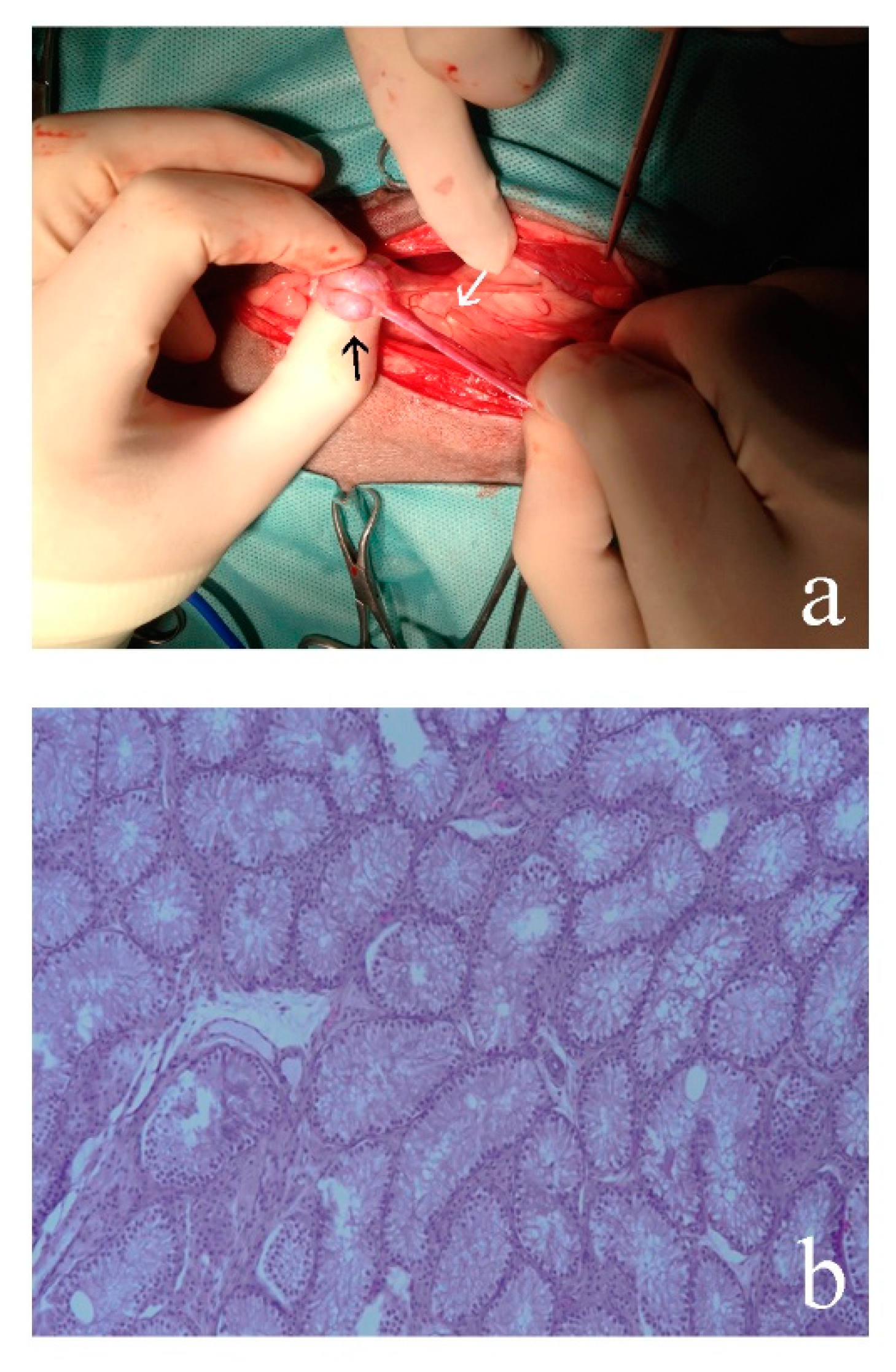
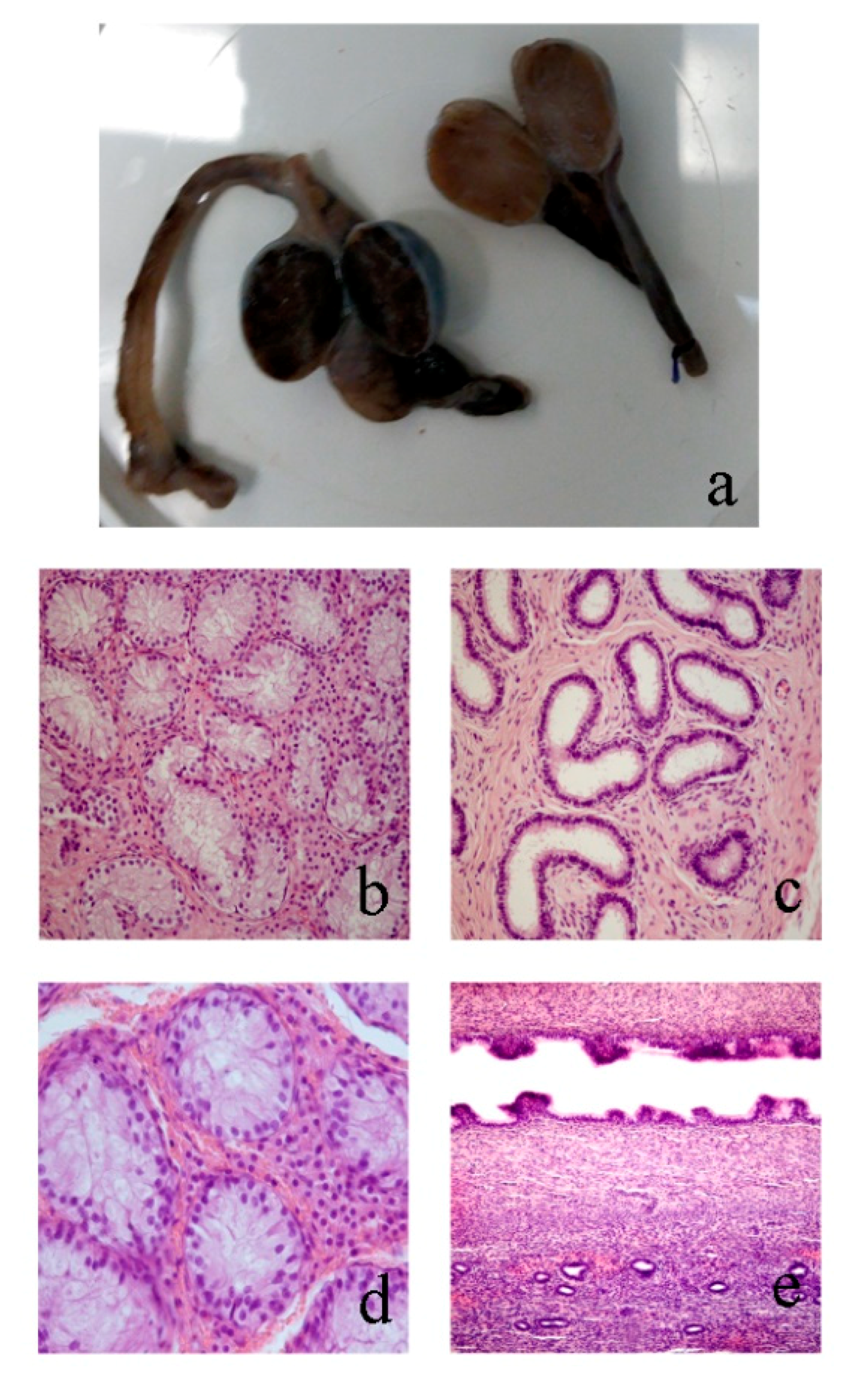
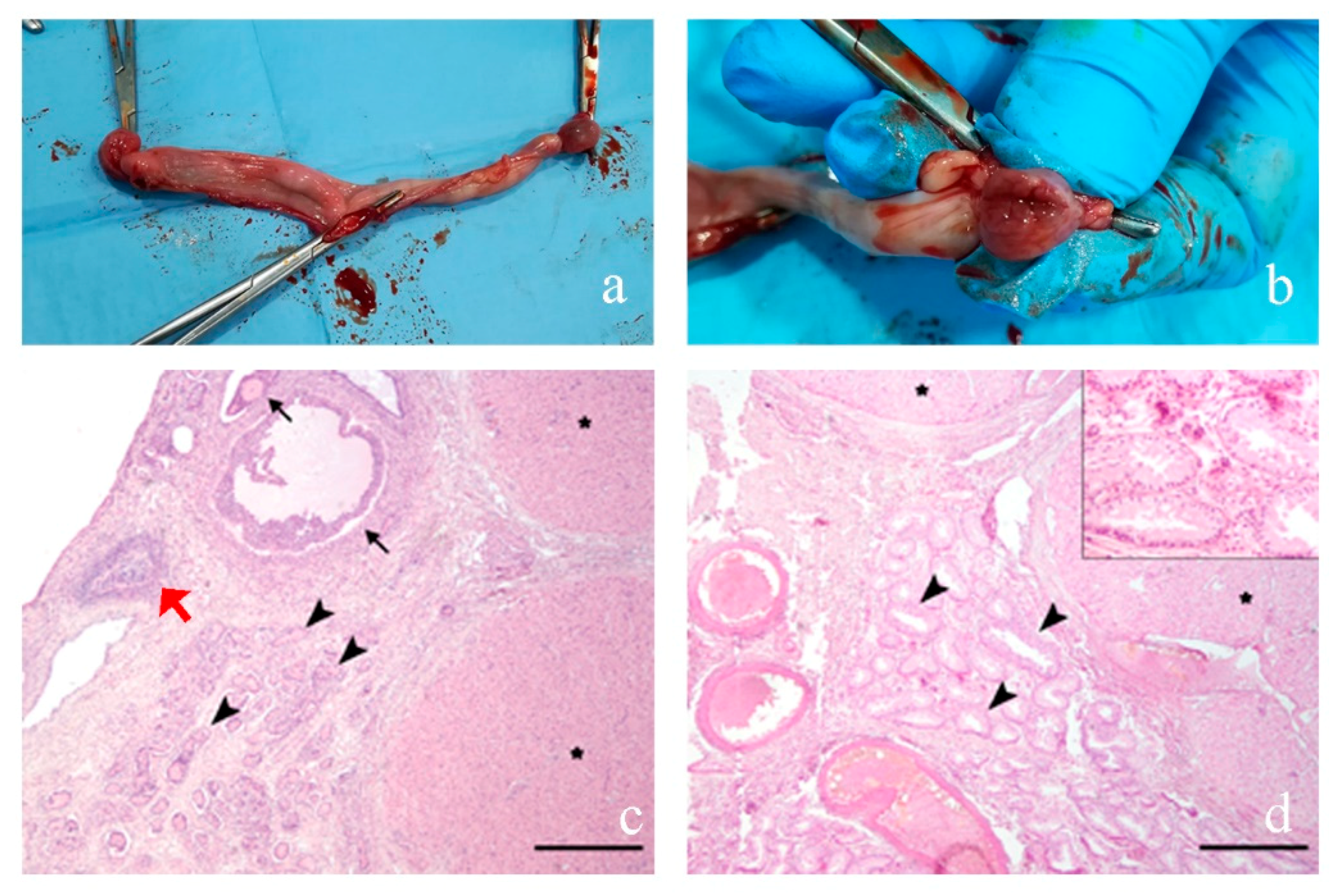
3.2. Morphological Analyses
3.2.1. Case 1
3.2.2. Case 2
3.2.3. Case 3
3.2.4. Case 4
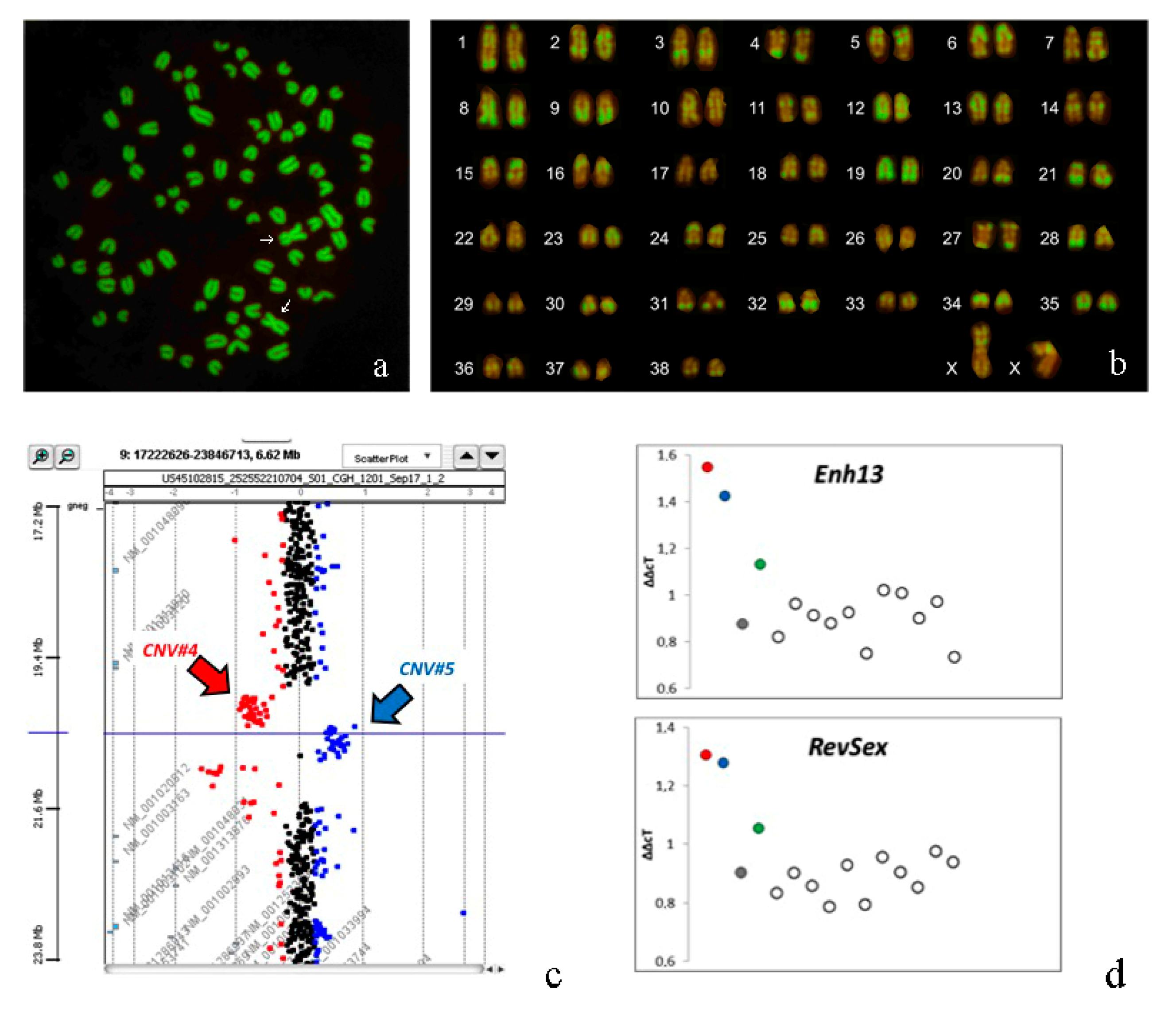
3.3. Cytogenetic Analyses
3.4. Molecular Analyses
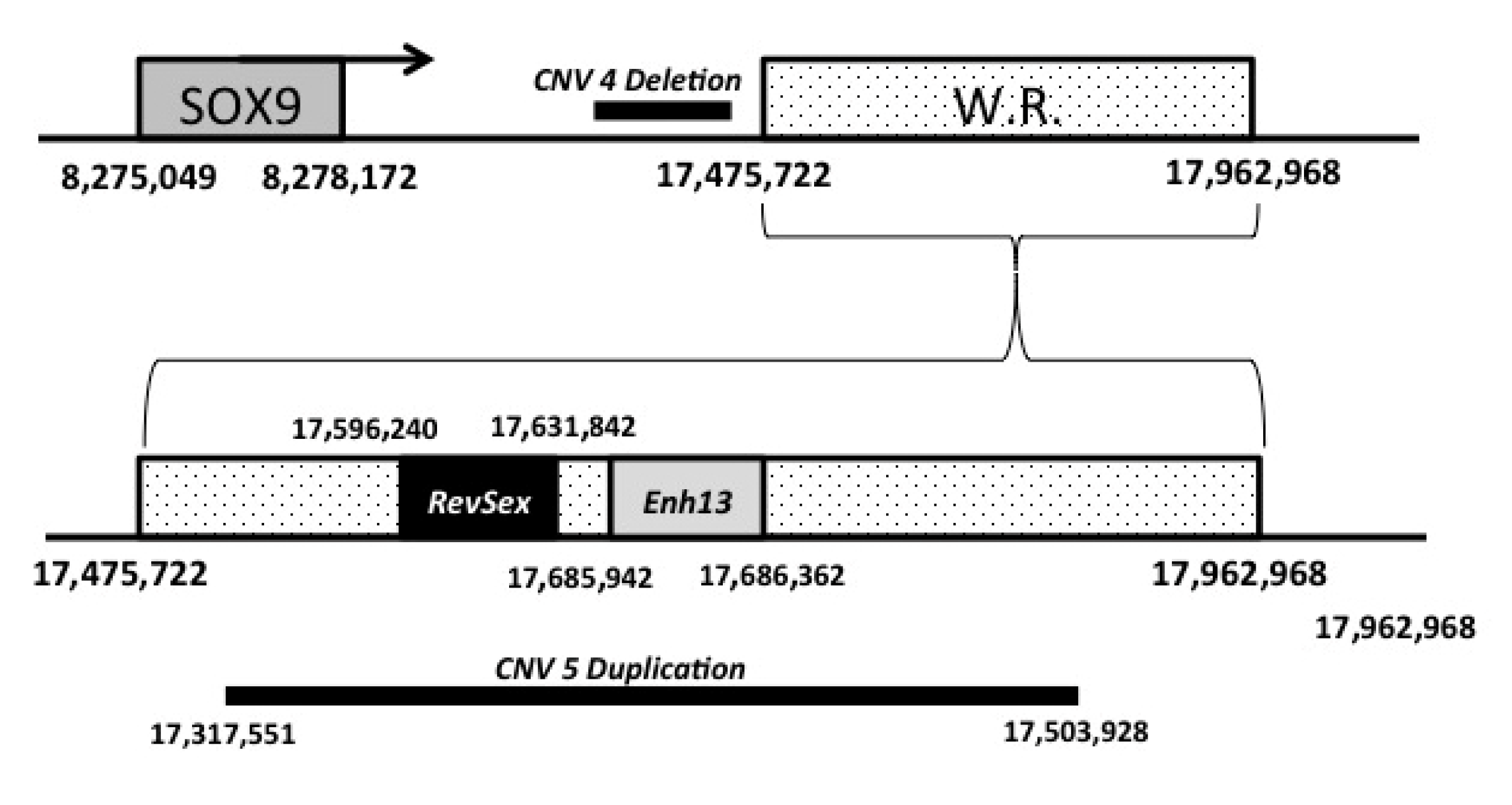
3.5. Array-CGH Analyses
3.6. qPCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Albarella, S.; De Lorenzi, L.; Catone, G.; Magi, G.E.; Petrucci, L.; Vullo, C.; D’Anza, E.; Parma, P.; Raudsepp, T.; Ciotola, F.; et al. Diagnosis of XX/XY Blood Cell Chimerism at a Low Percentage in Horses. J. Equine Vet. Sci. 2018, 69, 129–135. [Google Scholar] [CrossRef]
- Szczerbal, I.; Nowacka-Woszuk, J.; Albarella, S.; Switonski, M. Technical note: Droplet digital PCR as a new molecular method for a simple and reliable diagnosis of freemartinism in cattle. J. Dairy Sci. 2019, 102, 10100–10104. [Google Scholar] [CrossRef]
- Capel, B. To Be or Not To Be a Testis. Reproduction 2019. [Google Scholar] [CrossRef] [PubMed]
- Parma, P.; Veyrunes, F.; Pailhoux, E. Sex Reversal in Non-Human Placental Mammals. Sex Dev. 2016, 10, 326–344. [Google Scholar] [CrossRef] [PubMed]
- Pailhoux, E.; Popescu, P.; Parma, P.; Boscher, J.; Legault, C.; Molteni, L.; Fellous, M.; Cotinot, C. Genetic analysis of 38,XX males with genital ambiguities and true hermaphrodites in pigs. Anim. Genet. 1994, 25, 299–305. [Google Scholar] [CrossRef]
- Vaiman, D.; Koutita, O.; Oustry, A.; Elsen, J.M.; Manfredi, E.; Fellous, M.; Cribiu, E.P. Genetic mapping of the autosomal region involved in XX sex-reversal and horn development in goats. Mamm. Genome 1996, 7, 133–137. [Google Scholar] [CrossRef]
- Albarella, S.; D’Anza, E.; Galdiero, G.; Esposito, L.; De Biase, D.; Paciello, O.; Ciotola, F.; Peretti, V. Cytogenetic Analyses in Ewes with Congenital Abnormalities of the Genital Apparatus. Animals 2019, 10, 776. [Google Scholar] [CrossRef]
- Pajares, G.; Balseiro, A.; Pérez-Pardal, L.; Gamarra, J.A.; Monteagudo, L.V.; Goyache, F.; Royo, L.J. Sry-negative XX true hermaphroditism in a roe deer. Anim. Reprod. Sci. 2009, 112, 190–197. [Google Scholar] [CrossRef]
- Wilker, C.E.; Meyers-Wallen, V.N.; Schlafer, D.H.; Dykes, N.L.; Kovacs, A.; Ball, B.A. XX sex reversal in a llama. J. Am. Vet. Med. Assoc. 1994, 1, 112–115. [Google Scholar]
- De Lorenzi, L.; Arrighi, S.; Rossi, E.; Grignani, P.; Previderè, C.; Bonacina, S.; Cremonesi, F.; Parma, P. XY (SRY-positive) Ovarian Disorder of Sex Development in Cattle. Sex Dev. 2018, 12, 196–203. [Google Scholar] [CrossRef]
- Albarella, S.; Ciotola, F.; D’Anza, E.; Coletta, A.; Zicarelli, L.; Peretti, V. Congenital Malformations in River Buffalo (Bubalus bubalis). Animals 2017, 10, 7. [Google Scholar] [CrossRef] [PubMed]
- Villagómez, D.A.; Lear, T.L.; Chenier, T.; Lee, S.; McGee, R.B.; Cahill, J.; Foster, R.A.; Reyes, E.; St John, E.; King, W.A. Equine disorders of sexual development in 17 mares including XX, SRY-negative, XY, SRY-negative and XY, SRY-positive genotypes. Sex Dev. 2011, 5, 16–25. [Google Scholar] [CrossRef]
- Ciotola, F.; Albarella, S.; Pasolini, M.P.; Auletta, L.; Esposito, L.; Iannuzzi, L.; Peretti, V. Molecular and Cytogenetic Studies in a Case of XX SRY-Negative Sex Reversal in an Arabian Horse. Sex Dev. 2012, 6, 104–107. [Google Scholar] [CrossRef]
- De Lorenzi, L.; Banco, B.; Previderè, C.; Bonacina, S.; Romagnoli, S.; Grieco, V.; Parma, P. Testicular XX (SRY-Negative) Disorder of Sex Development in Cat. Sex Dev. 2017, 11, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Krzemińska, P.; D’Anza, E.; Ciotola, F.; Paciello, O.; Restucci, B.; Peretti, V.; Albarella, S.; Switonski, M. Polymorphisms of MAMLD1, SRD5A2, and AR Candidate Genes in Seven Dogs (78,XY; SRY-Positive) Affected by Hypospadias or Cryptorchidism. Sex Dev. 2019, 13, 92–98. [Google Scholar] [CrossRef]
- Nowacka-Woszuk, J.; Szczerbal, I.; Pausch, H.; Hundi, S.; Hytönen, M.K.; Grzemski, A.; Flisikowski, K.; Lohi, H.; Switonski, M.; Szydlowski, M. Deep sequencing of a candidate region harboring the SOX9 gene for the canine XX disorder of sex development. Anim. Genet. 2017, 48, 330–337. [Google Scholar] [CrossRef] [PubMed]
- d’Ovidio, D.; Melidone, R.; Rossi, G.; Albarella, S.; Noviello, E.; Fioretti, A.; Meomartino, L. Multiple congenital malformations in a ferret (Mustela Putorius Furo). J. Exot. Pet Med. 2015, 24, 92–97. [Google Scholar] [CrossRef]
- Vetro, A.; Dehghani, M.R.; Kraoua, L.; Giorda, R.; Beri, S.; Cardarelli, L.; Merico, M.; Manolakos, E.; Parada-Bustamante, A.; Castro, A.; et al. Testis development in the absence of SRY: Chromosomal rearrangements at SOX9 and SOX3. Eur. J. Hum. Genet. 2015, 23, 1025–1032. [Google Scholar] [CrossRef]
- Gonen, N.; Futtner, C.R.; Wood, S.; Garcia-Moreno, S.A.; Salamone, I.M.; Samson, S.C.; Sekido, R.; Poulat, F.; Maatouk, D.M.; Lovell-Badge, R. Sex reversal following deletion of a single distal enhancer of Sox9. Science 2018, 29, 1469–1473. [Google Scholar] [CrossRef]
- Rossi, E.; Radi, O.; De Lorenzi, L.; Vetro, A.; Groppetti, D.; Bigliardi, E.; Luvoni, G.C.; Rota, A.; Camerino, G.; Zuffardi, O.; et al. Sox9 duplications are a relevant cause of Sry-negative XX sex reversal dogs. PLoS ONE 2014, 10, e101244. [Google Scholar] [CrossRef]
- Parma, P.; Radi, O.; Vidal, V.; Chaboissier, M.C.; Dellambra, E.; Valentini, S.; Guerra, L.; Schedl, A.; Camerino, G. R-spondin 1 is essential in sex determination, skin differentiation and malignancy. Nat. Genet. 2006, 38, 1304–1309. [Google Scholar] [CrossRef]
- Pailhoux, E.; Vigier, B.; Chaffaux, S.; Servel, N.; Taourit, S.; Furet, J.-P.; Fellos, M.; Grosclaude, F.; Cribiu, E.P.; Cotinot, C.; et al. A 11.7-kb deletion triggers intersexuality and polledness in goats. Nat. Genet. 2001, 29, 453–458. [Google Scholar] [CrossRef]
- Sutton, E.; Hughes, J.; White, S.; Sekido, R.; Tan, J.; Arboleda, V.; Rogers, N.; Knower, K.; Rowley, L.; Eyre, H.; et al. Identification of SOX3 as an XX male sex reversal gene in mice and humans. J. Clin. Investig. 2011, 121, 328–341. [Google Scholar] [CrossRef] [PubMed]
- Macrì, F.; Ciotola, F.; Rapisarda, G.; Lanteri, G.; Albarella, S.; Aiudi, G.; Liotta, L.; Marino, F. A rare case of simple syndactyly in a puppy. J. Small Anim. Pract. 2014, 55, 170–173. [Google Scholar] [CrossRef]
- Switoński, M.; Reimann, N.; Bosma, A.A.; Long, S.; Bartnitzke, S.; Pieńkowska, A.; Moreno-Milan, M.M.; Fischer, P. Report on the progress of standardization of the G-banded canine (Canis familiaris) karyotype. Committee for the Standardized Karyotype of the Dog (Canis familiaris). Chromosome Res. 1996, 4, 306–309. [Google Scholar] [CrossRef]
- Yan, S.; Bai, C.; Li, Y.; Li, Y.; Hou, J.; Zhao, Z.; Han, W. Sex identification of dog by PCR based on the differences in the AMELX and AMELY genes. Anim. Genet. 2013, 44, 606. [Google Scholar] [CrossRef]
- De Lorenzi, L.; Groppetti, D.; Arrighi, S.; Pujar, S.; Nicoloso, L.; Molteni, L.; Pecile, A.; Cremonesi, F.; Parma, P.; Meyers-Wallen, V. Mutations in the RSPO1 coding region are not the main cause of canine SRY-negative XX sex reversal in several breeds. Sex Dev. 2008, 2, 84–95. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzi, L.; Kopecna, O.; Gimelli, S.; Cernohorska, H.; Zannotti, M.; Béna, F.; Molteni, L.; Rubes, J.; Parma, P. Reciprocal translocation t(4;7)(q14;q28) in cattle: Molecular characterization. Cytogenet. Genome Res. 2010, 129, 298–304. [Google Scholar] [CrossRef]
- Hubler, M.; Hauser, B.; Meyers-Wallen, V.N.; Arnold, S. Sry-negative XX true hermaphrodite in a Basset hound. Theriogenology 1999, 51, 1391–1403. [Google Scholar] [CrossRef]
- Nowacka-Woszuk, J.; Szczerbal, I.; Stachowiak, M.; Szydlowski, M.; Nizanski, W.; Dzimira, S.; Maslak, A.; Payan-Carreira, R.; Wydooghe, E.; Nowak, T.; et al. Association between polymorphisms in the SOX9 region and canine disorder of sex development (78,XX; SRY-negative) revisited in a multibreed case-control study. PLoS ONE 2019, 14, e0218565. [Google Scholar] [CrossRef] [PubMed]
- Szczerbal, I.; Nowacka-Woszuk, J.; Nizanski, W.; Dzimira, S.; Ligocka, Z.; Jastrzebska, A.; Kabala, B.; Biernacik, M.; Przadka, P.; Switonski, M. Disorders of Sex Development Are an Emerging Problem in French Bulldogs: A Description of Six New Cases and a Review of the Literature. Sex Dev. 2020, 13, 205–211. [Google Scholar] [CrossRef]
- Poth, T.; Breuer, W.; Walter, B.; Hecht, W.; Hermanns, W. Disorders of sex development in the dog-Adoption of a new nomenclature and reclassification of reported cases. Anim. Reprod. Sci. 2010, 121, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Rossi, E.; Radi, O.; De Lorenzi, L.; Iannuzzi, A.; Camerino, G.; Zuffardi, O.; Parma, P. A Revised Genome Assembly of the Region 5’ to Canine SOX9 Includes the RevSex Orthologous Region. Sex Dev. 2015, 9, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.K.; Swartz, J.D.; Rush, L.J.; Alvarez, C.E. Mapping DNA structural variation in dogs. Genome Res. 2009, 19, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Mortlock, S.A.; Williamson, P.; Khatkar, M.S. Copy number variation and variant discovery in Bullmastiff dogs. Anim. Genet. 2019, 50, 177–181. [Google Scholar] [CrossRef]
- Nicholas, T.J.; Baker, C.; Eichler, E.E.; Akey, J.M. A high-resolution integrated map of copy number polymorphisms within and between breeds of the modern domesticated dog. BMC Genom. 2011, 12, 414. [Google Scholar] [CrossRef]
- Talas, U.; Dunlop, J.; Khalaf, S.; Leigh, I.M.; Kelsell, D.P. Human elastase 1: Evidence for expression in the skin and the identification of a frequent frameshift polymorphism. J. Investig. Derm. 2000, 114, 165–170. [Google Scholar] [CrossRef]
- Bennett, E.P.; Hassan, H.; Mandel, U.; Hollingsworth, M.A.; Akisawa, N.; Ikematsu, Y.; Merkx, G.; Geurts van Kessel, A.; Olofsson, S.; Clausen, H. Cloning and characterization of a close homologue of human UDP-N-acetyl-alpha-d-galactosamine:polypeptide N-acetylgalactosaminyltransferase-T3, designated GalNAc-T6: Evidence for genetic but not functional redundancy. J. Biol. Chem. 1999, 274, 25362–25370. [Google Scholar] [CrossRef]
- Ge, K.; Prendergast, G.C. Bin2, a functionally nonredundant member of the BAR adaptor gene family. Genomics 2000, 67, 210–220. [Google Scholar] [CrossRef]
- Oh, B.; Hwang, S.Y.; Solter, D.; Knowles, B.B. Spindlin, a major maternal transcript expressed in the mouse during the transition from oocyte to embryo. Development 1997, 124, 493–503. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | Sequence (5′-3′) | Amp Size (bp) | Genome Pos 1 |
|---|---|---|---|---|
| Sry | Sry-F | GCTGGGCGGAGAAATGAGTA | 783 | Not available |
| Sry-R | CCAAGGTTTCCGGACTGTCA | |||
| Amelx/y | DSI-F | ATAATGACAAAGAAAACATGAC | 215/247 | Chrx: 7,828,350–7,828,136 2 |
| DSI-R | CTGCTGAGCTGGCACCAT | |||
| Rspo1 | Rspo1-Ex1-f | GCAGGCGTTAGCAAGAGC | 297 | Chr15: 4,862,841–4,863,138 |
| Rspo1-Ex1-r | ATCTGCAACGGTCATCACG | |||
| Rspo1-Ex2-f | AAGCACGTTCACGTTAGTCTTG | 398 | Chr15: 4,871,649–4,872,047 | |
| Rspo1-Ex2-r | ACCAATGGGTCAAAGCACTC | |||
| Rspo1-Ex3-f | GTCACTCGGGCCTCCTCTA | 478 | Chr15: 4,873,791–4,874,269 | |
| Rspo1-Ex3-r | GCAGAAAAGCTCGGAGACAA | |||
| Rspo1-Ex4-f | ACTGACACTGCCTCCAGCAT | 480 | Chr15: 4,874,276–4,874,754 | |
| Rspo1-Ex4-r | CTGTTGTCTGCCAGCGTCT | |||
| Rspo1-Ex5-f | GGGGACCCTGAGACTGTGTA | 399 | Chr15: 4,875,173-4,875,572 | |
| Rspo1-Ex5-r | TCCAGTTCCGTAAAGCTTCC | |||
| Enh13 | Enh13-f | GCAATGTGCACAGTTTCAGAG | 118 | Chr9: 17,686,202–17,686,320 |
| Enh13-r | TGAGGAATTAGAAGGCCATGA | |||
| RevSex | RevSex-Dog-F | GACACTGTCCTGGGGAGAAA | 100 | Chr9:17,605,471- 17,605,570 |
| RevSex-Dog-R | TGAAGGCCAAGAGGCTAAGA | |||
| Bglr2 | BGLR2-F | GTGGAAGCCTGCAATTGTCT | 203 | Chr6: 734,406–734,609 |
| BGLR2-R | CCGTGAACAGGTGTAATGCT |
| Case Number | Breed | Age at First Clinical Evaluation | Phenotype | Surgical Findings |
|---|---|---|---|---|
| 1 | Staffordshire terrier | 1Y | Female. | Ultrasound showed two ovotestis like structures in the abdomen caudally to the kidneys. |
| 2 | French bulldog | 9M | Female. Presence of a little palpable mass in inguinal region | A gonad with a uterine horn was removed from the abdomen and another gonad was removed from the inguinal region |
| 3 | French bulldog | 6M | Female. Presence of a little palpable mass in inguinal region | One gonad was in inguinal position and connected with a tubular structure. The other gonad was in the abdomen caudally to the kidney and was connected with a tubular structure that showed a fork linked to the vas deferens. Both structures ended in a uterine horn-like structure that lead directly into the vagina. The prostate was absent. Enlarged clitoris with penis bone protruded from an abnormal vulvae opening. |
| 4 | Mongrel | 1Y | Female. 4.8 kg. | Two ovotestis like structures were found in the abdomen connected with uterine horns that merge in a uterine like structure ending in the vagina. |
| CNV | CHR | CanFam3 | END (bp) | Analyzed Subjects | ||||
|---|---|---|---|---|---|---|---|---|
| START (bp) | SIZE (kb) | 1 | 2 | 3 | 4 | |||
| 1 | Chr 4 | 106,352 | 469,199 | 363 | DEL | DEL | DEL | DEL |
| 2 | Chr 5 | 78,189,869 | 78,389,978 | 200 | DEL | DUP | DEL | |
| 3 | Chr 6 | 45,163,433 | 47,125,036 | 1962 | GAIN | GAIN | GAIN | GAIN |
| 4 | Chr 9 | 16,906,864 | 17,317,551 | 411 | DEL | DEL | DEL | |
| 5 | Chr 9 | 17,503,928 | 17,962,221 | 458 | DUP | DUP | DUP | |
| 6 | Chr 9 | 38,978,944 | 38,995,409 | 16 | DEL | DEL | DEL | |
| 7 | Chr 12 | 2,191,427 | 2,270,973 | 80 | DEL | DEL | DEL | |
| 8 | Chr 23 | 20,508,926 | 20,725,281 | 216 | GAIN | GAIN | ||
| 9 | Chr 26 | 27,171,599 | 27,220,687 | 49 | DELHO | DELHO | DELHO | DELHO |
| 10 | Chr 27 | 3,532,831 | 3,573,782 | 41 | GAIN | GAIN | ||
| 11 | Chr X | 71,752,458 | 72,234,092 | 482 | DELHO | DEL | DEL | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albarella, S.; Lorenzi, L.D.; Rossi, E.; Prisco, F.; Riccardi, M.G.; Restucci, B.; Ciotola, F.; Parma, P. Analysis of XX SRY-Negative Sex Reversal Dogs. Animals 2020, 10, 1667. https://doi.org/10.3390/ani10091667
Albarella S, Lorenzi LD, Rossi E, Prisco F, Riccardi MG, Restucci B, Ciotola F, Parma P. Analysis of XX SRY-Negative Sex Reversal Dogs. Animals. 2020; 10(9):1667. https://doi.org/10.3390/ani10091667
Chicago/Turabian StyleAlbarella, Sara, Lisa De Lorenzi, Elena Rossi, Francesco Prisco, Marita Georgia Riccardi, Brunella Restucci, Francesca Ciotola, and Pietro Parma. 2020. "Analysis of XX SRY-Negative Sex Reversal Dogs" Animals 10, no. 9: 1667. https://doi.org/10.3390/ani10091667
APA StyleAlbarella, S., Lorenzi, L. D., Rossi, E., Prisco, F., Riccardi, M. G., Restucci, B., Ciotola, F., & Parma, P. (2020). Analysis of XX SRY-Negative Sex Reversal Dogs. Animals, 10(9), 1667. https://doi.org/10.3390/ani10091667