Effects of Aging on Expression of Mic60 and OPA1 and Mitochondrial Morphology in Myocardium of Tibetan Sheep


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Isolation of the Myocardial Mitochondria
2.3. Extraction of Tissue RNA
2.4. Design and Synthesis of Primers
2.5. RT-qPCR Reaction
2.6. Immunohistochemistry and ELISA
2.7. Transmission Electron Microscopy Preparation
2.8. Statistical Analyses
3. Results
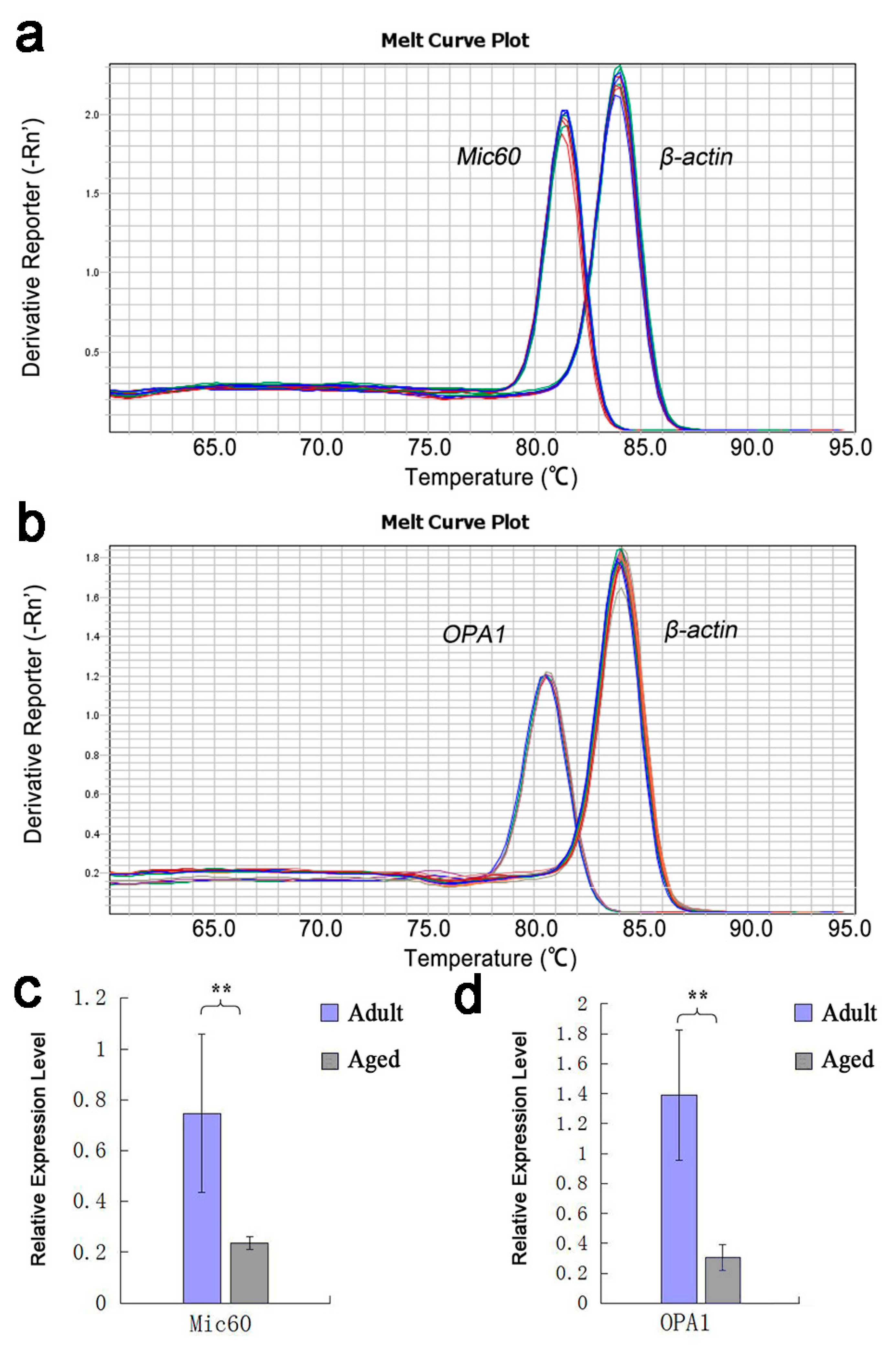
3.1. Analysis of the Result of RT-qPCR
3.2. Expression and Analysis of Mic60 and OPA1 Proteins
3.3. Mitochondrial Morphological Structure Comparison
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dimmer, K.S.; Scorrano, L. (De) constructing mitochondria: What for? Physiology 2006, 21, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Frezza, C.; Cipolat, S.; Brito, O.M.D.; Micaroni, M.; Beznoussenko, G.V.; Rudka, R.T.; Bartoli, D.; Polishuck, R.S.; Danial, N.N.; Strooper, B.D.; et al. OPA1 Controls Apoptotic Cristae Remodeling Independently from Mitochondrial Fusion. Cell 2006, 126, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Shaw, J.M. Mitochondrial Morphology and Dynamics in Yeast and Multicellular Eukaryotes. Annu. Rev. Genet. 2005, 39, 503–536. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.C. Mitochondrial Fusion and Fission in Mammals. Annu. Rev. Cell. Dev. Bi. 2006, 22, 79–99. [Google Scholar] [CrossRef]
- Cogliati, S.; Frezza, C.; Soriano, M.E.; Varanita, T.; Quintana-Cabrera, R.; Corrado, M.; Cipolat, S.; Costa, V.; Casarin, A.; Gomes, L.C.; et al. Mitochondrial Cristae Shape Determines Respiratory Chain Supercomplexes Assembly and Respiratory Efficiency. Cell 2013, 155, 160–171. [Google Scholar] [CrossRef]
- Gilkerson, R.W.; Selker, J.M.L.; Capaldi, R.A. The cristal membrane of mitochondria is the principal site of oxidative phosphorylation. FEBS Lett. 2003, 546, 355–358. [Google Scholar] [CrossRef]
- Chaban, Y.; Boekema, E.J.; Dudkina, N.V. Structures of mitochondrial oxidative phosphorylation supercomplexes and mechanisms for their stabilisation. BBA-Bioenerg. 2014, 1837, 418–426. [Google Scholar] [CrossRef]
- Vogel, F.; Bornhovd, C.; Neupert, W.; Reicher, A.S. Dynamic subcompartmentalization of the mitochondrial inner membrane. J. Cell Biol. 2006, 175, 237–247. [Google Scholar] [CrossRef]
- Carelli, V.; Morgia, C.L.; Iommarini, L.; Carroccia, R.; Mattiazzi, M.; Sangiorgi, S.; Farne, S.; Maresca, A.; Foscarini, B.; Lanzi, L.; et al. Mitochondrial Optic Neuropathies: How Two Genomes may Kill the Same Cell Type? Biosci. Rep. 2007, 27, 173–184. [Google Scholar] [CrossRef]
- Gieffers, C.; Korioth, F.; Heimann, P.; Ungermann, C.; Frey, J. Mitofilin is a transmembrane protein of the inner mitochondrial membrane expressed as two isoforms. Exp. Cell. Res. 1997, 232, 395–399. [Google Scholar] [CrossRef]
- Olichon, A.; Elachouri, G.; Baricault, L.; Delettre, C.; Belenguer, P.; Lenaers, G. OPA1 alternate splicing uncouples an evolutionary conserved function in mitochondrial fusion from a vertebrate restricted function in apoptosis. Cell Death Differ. 2007, 14, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Youle, R.J.; van der Bliek, A.M. Mitochondrial Fission, Fusion, and Stress. Science 2012, 337, 1062–1065. [Google Scholar] [CrossRef] [PubMed]
- Spinazzi, M.; Cazzola, S.; Bortolozzi, M.; Baracca, A.; Loro, E.; Casarin, A.; Solaini, G.; Sgarbi, G.; Casalena, G.; Cenacchi, G.; et al. A novel deletion in the gtpase domain of opa1 causes defects in mitochondrial morphology and distribution, but not in function. Hum. Mol. Genet. 2008, 17, 3291–3302. [Google Scholar] [CrossRef] [PubMed]
- Griparic, L.; Kanazawa, T.; Van Der Bliek, A.M. Regulation of the mitochondrial dynamin-like protein Opa1 by proteolytic cleavage. J. Cell Biol. 2007, 178, 757–764. [Google Scholar] [CrossRef]
- Griparic, L.; Van Der Wel, N.N.; Orozco, I.J.; Peters, P.J.; Van Der Bliek, A.M. Loss of the intermembrane space protein Mgm1/OPA1 induces swelling and localized constrictions along the lengths of mitochondria. J. Biol. Chem. 2004, 279, 18792–18798. [Google Scholar] [CrossRef]
- Von der Malsburg, K.; Mueller, J.M.; Bohnert, M.; Oeljeklaus, S.; Kwiatkowska, P.; Becker, T.; Loniewska-Lwowska, A.; Wiese, S.; Rao, S.; Milenkovic, D.; et al. Dual Role of Mitofilin in Mitochondrial Membrane Organization and Protein Biogenesis. Dev. Cell 2011, 21, 694–707. [Google Scholar] [CrossRef]
- Head, B.P.; Zulaika, M.; Ryazantsev, S.; van der Bliek, A.M. A novel mitochondrial outer membrane protein, MOMA-1, that affects cristae morphology in Caenorhabditis elegans. Mol. Biol. Cell 2011, 22, 831–841. [Google Scholar] [CrossRef]
- Yang, R.F.; Sun, L.H.; Zhang, R.; Zhang, Y.; Luo, Y.X.; Zheng, W.; Zhang, Z.Q.; Chen, H.Z.; Liu, D.P. Suppression of Mic60 compromises mitochondrial transcription and oxidative phosphorylation. Sci. Rep. 2015, 5, 7990. [Google Scholar] [CrossRef]
- Koerner, C.; Barrera, M.; Dukanovic, J.; Eydt, K.; Harner, M.; Rabl, R.; Vogel, F.; Rapaport, D.; Neupert, W.; Reichert, A.S. The C-terminal domain of Fcj1 is required for formation of crista junctions and interacts with the TOB/SAM complex in mitochondria. Mol. Biol. Cell 2012, 23, 2143–2155. [Google Scholar] [CrossRef]
- John, G.B.; Shang, Y.; Li, L.; Renken, C.; Mannella, C.A.; Selker, J.M.L.; Rangell, L.; Bennett, M.J.; Zha, J. The mitochondrial inner membrane protein mitofilin controls cristae morphology. Mol. Biol. Cell 2005, 16, 1543–1554. [Google Scholar] [CrossRef]
- Chantler, P.D.; Goldspink, D.F.; Clements, R.E.; Sharp, L.; Schlosshan, D.; Tan, L.B. Congestive heart failure: Extent of cardiac functional changes caused by aging and organ dysfunction. Heart 2006, 92, 686–688. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Livak, K.; Schmittgen, T. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-△△Ct Method. Methods 2000, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L.; Sun, L.H.; Yue, Y.S. Correlation between Mic60 haploid insufficiency and cardiac aging in mouse. Chin. J. Pathol. 2017, 46, 406–410. [Google Scholar]
- Eckert, G.P.; Schiborr, C.; Hagl, S.; Abdel-Kader, R.; Muller, W.E.; Rimbach, G.; Frank, J. Curcumin prevents mitochondrial dysfunction in the brain of the senescence-accelerated mouse-prone 8. Neurochem. Int. 2013, 62, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wang, H.; Wang, Z.; Wang, Q.; Zhang, S.; Deng, Y.; Fang, Y. In vivo imaging reveals mitophagy independence in the maintenance of axonal mitochondria during normal aging. Aging Cell 2017, 16, 1180–1190. [Google Scholar]
- Preston, C.C.; Oberlin, A.S.; Holmuhamedov, E.L.; Gupta, A.; Sagar, S.; Syed, R.H.K.; Siddiqui, S.A.; Raghavakaimal, S.; Terzic, A.; Jahangir, A. Aging-induced alterations in gene transcripts and functional activity of mitochondrial oxidative phosphorylation complexes in the heart. Mech. Ageing Dev. 2008, 129, 304. [Google Scholar] [CrossRef]
- Simonson, T.S.; Yang, Y.; Huff, C.D.; Yun, H.; Qin, G.; Witherspoon, D.J.; Bai, Z.; Lorenzo, F.R.; Xing, J.; Jorde, L.B.; et al. Genetic evidence for high-altitude adaptation in Tibet. Science 2010, 329, 72. [Google Scholar] [CrossRef]
- He, Y.; Yu, S.; Hu, J.; Cui, Y.; Liu, P. Changes in the Anatomic and Microscopic Structure and the Expression of HIF-1α and VEGF of the Yak Heart with Aging and Hypoxia. PLoS ONE 2016, 11, e0149947. [Google Scholar] [CrossRef]
- Patten, D.A.; Wong, J.; Khacho, M.; Soubannier, V.; Mailloux, R.J.; Pilon-Larose, K.; MacLaurin, J.G.; Park, D.S.; McBride, H.M.; Trinkle-Mulcahy, L.; et al. OPA1-dependent cristae modulation is essential for cellular adaptation to metabolic demand. Embo J. 2014, 33, 2676–2691. [Google Scholar] [CrossRef]
- Meeusen, S.; DeVay, R.; Block, J.; Cassidy-Stone, A.; Wayson, S.; McCaffery, J.M.; Nunnari, J. Mitochondrial inner-membrane fusion and crista maintenance requires the dynamin-related GTPase Mgm1. Cell 2006, 127, 383–395. [Google Scholar] [CrossRef]
- Darshi, M.; Mendiola, V.L.; Mackey, M.R.; Murphy, A.N.; Koller, A.; Perkins, G.A.; Ellisman, M.H.; Taylor, S.S. ChChd3, an Inner Mitochondrial Membrane Protein, Is Essential for Maintaining Crista Integrity and Mitochondrial Function. J. Biol. Chem. 2011, 286, 2918–2932. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | GenBank Accession No. | Primer Sequence (5′–3′) | Primer Length | Product Length |
|---|---|---|---|---|
| Mic60 | XM_012169573 | F:TTGAGATGGTCCTTGGTT | 18 | 136 |
| R:TTGTTTCTGAGGTGGTGAG | 19 | |||
| OPA1 | XM_012140446 | F:ATGAAATAGAACTCCGAATG | 20 | 112 |
| R:GTCAACAAGCACCATCCT | 18 | |||
| β-actin | NM_001009784 | F: AGCCTTCCTTCCTGGGCATGGA | 22 | 113 |
| R:GGACAGCACCGTGTTGGCGTAGA | 23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Luo, Y.; Hu, J.; Wang, J.; Liu, X.; Li, S. Effects of Aging on Expression of Mic60 and OPA1 and Mitochondrial Morphology in Myocardium of Tibetan Sheep. Animals 2020, 10, 2160. https://doi.org/10.3390/ani10112160
Wang G, Luo Y, Hu J, Wang J, Liu X, Li S. Effects of Aging on Expression of Mic60 and OPA1 and Mitochondrial Morphology in Myocardium of Tibetan Sheep. Animals. 2020; 10(11):2160. https://doi.org/10.3390/ani10112160
Chicago/Turabian StyleWang, Guan, Yuzhu Luo, Jiang Hu, Jiqing Wang, Xiu Liu, and Shaobin Li. 2020. "Effects of Aging on Expression of Mic60 and OPA1 and Mitochondrial Morphology in Myocardium of Tibetan Sheep" Animals 10, no. 11: 2160. https://doi.org/10.3390/ani10112160
APA StyleWang, G., Luo, Y., Hu, J., Wang, J., Liu, X., & Li, S. (2020). Effects of Aging on Expression of Mic60 and OPA1 and Mitochondrial Morphology in Myocardium of Tibetan Sheep. Animals, 10(11), 2160. https://doi.org/10.3390/ani10112160